
Expression of Transcription Factor ZBTB20 in the Adult Primate Neurogenic Niche under Physiological Conditions or after Ischemia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Immunofluorescence
2.3. Image Acquisition and Analysis
2.4. ISH Image Analysis
3. Results
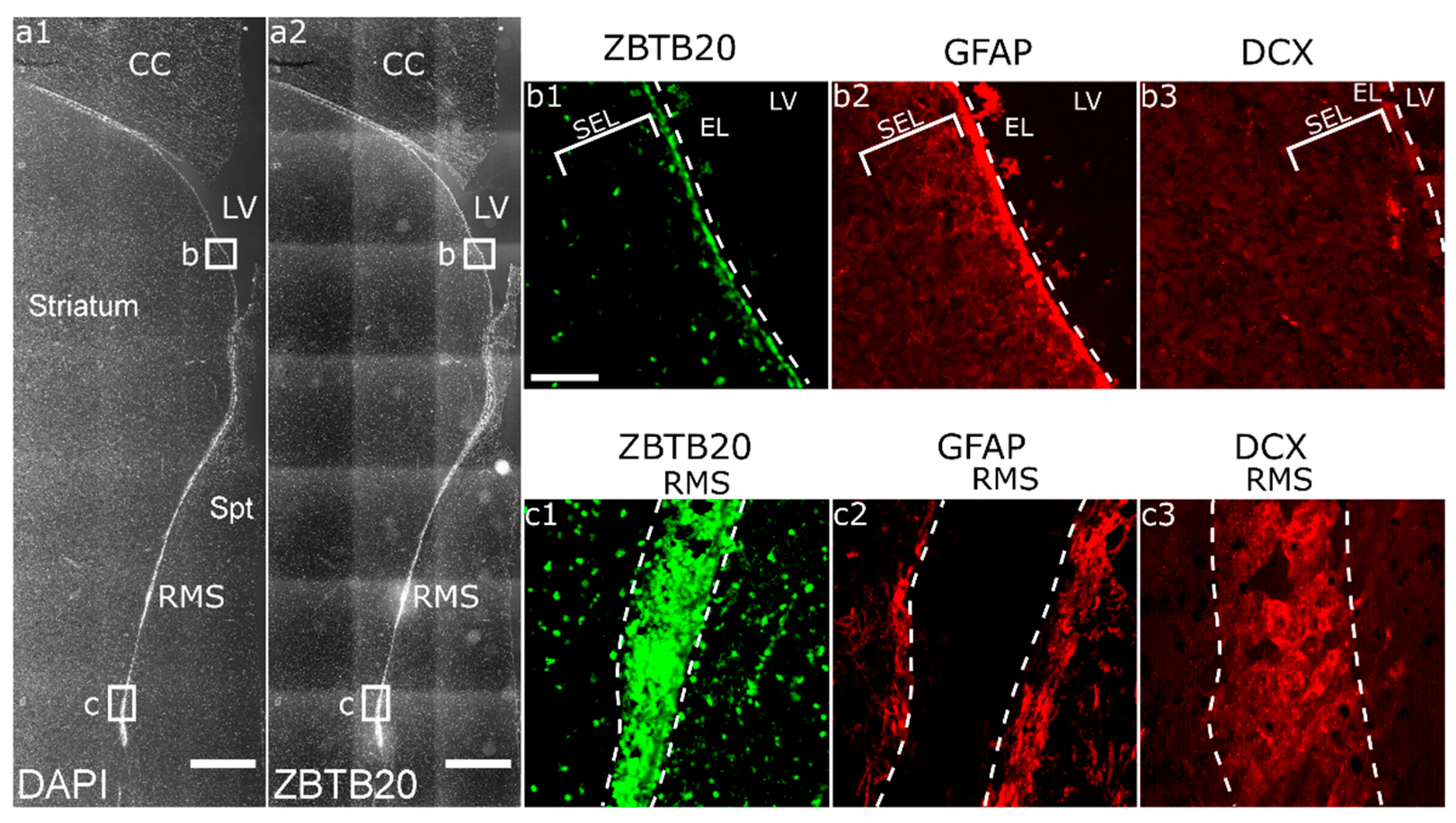
3.1. ZBTB20 Is Expressed in the Macaque SVZa
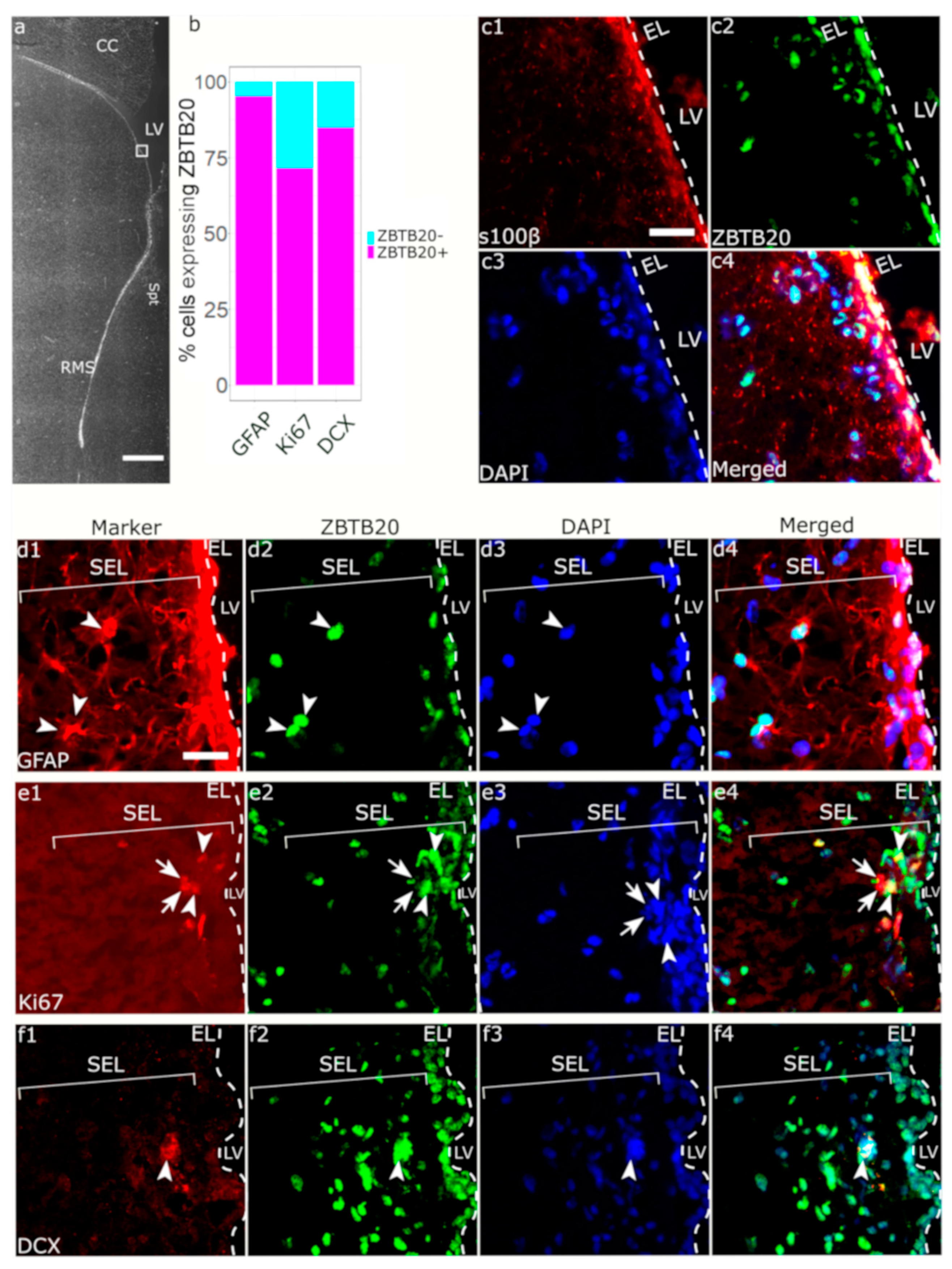
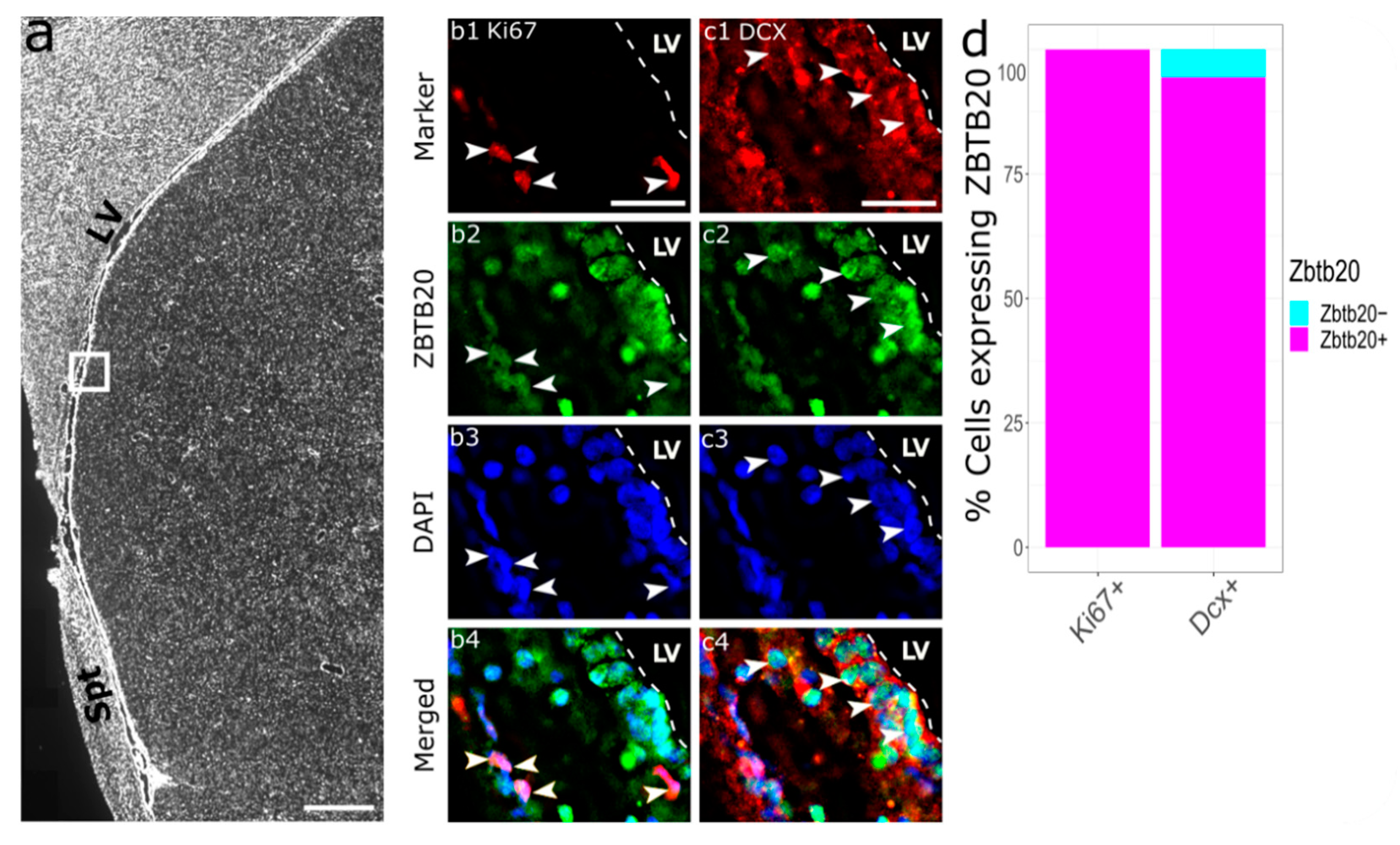
3.2. Immunohistochemical Characterization of ZBTB20+ Cells in Macaque SVZa
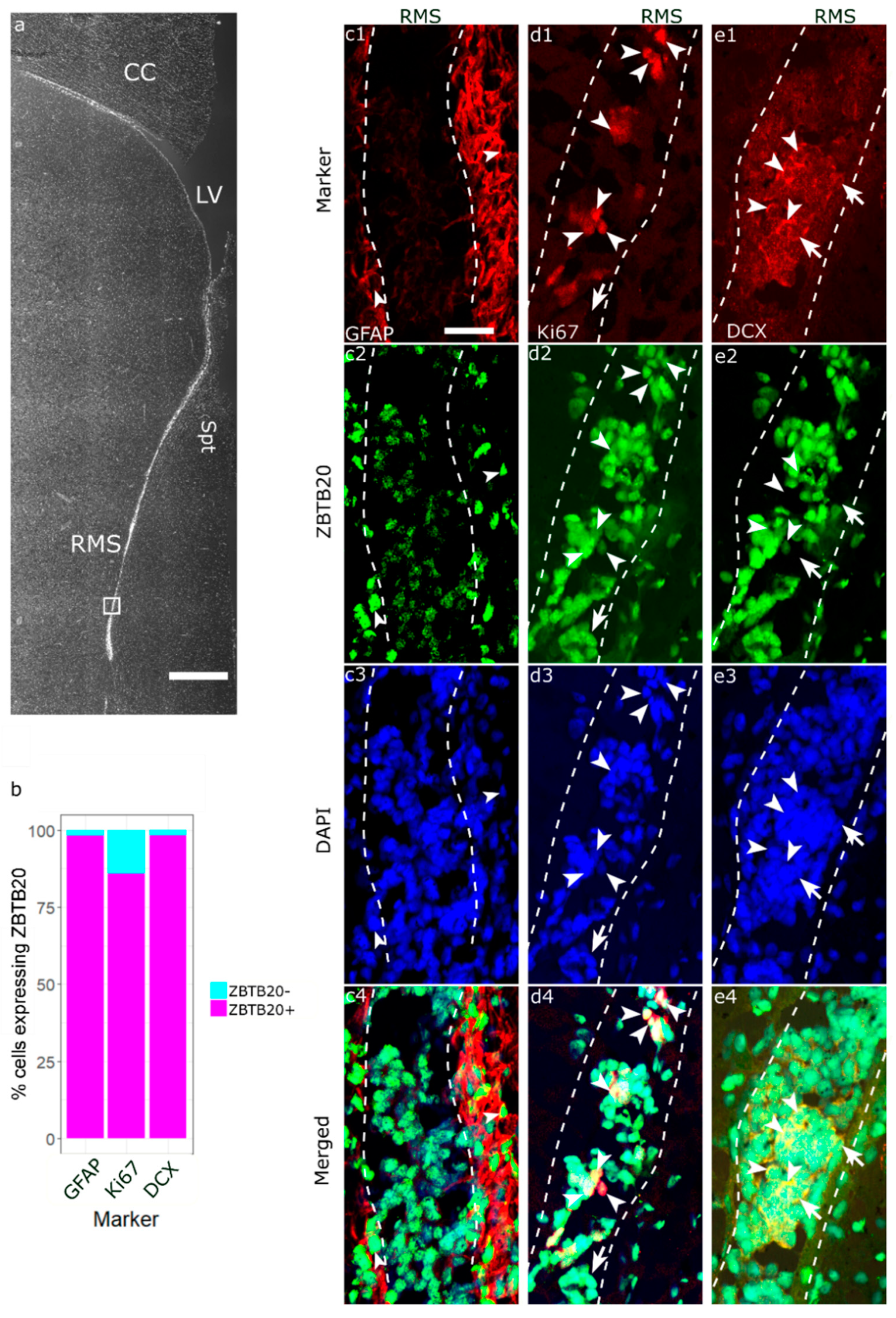
3.3. Immunohistochemical Characterization of ZBTB20+ Cells in RMS
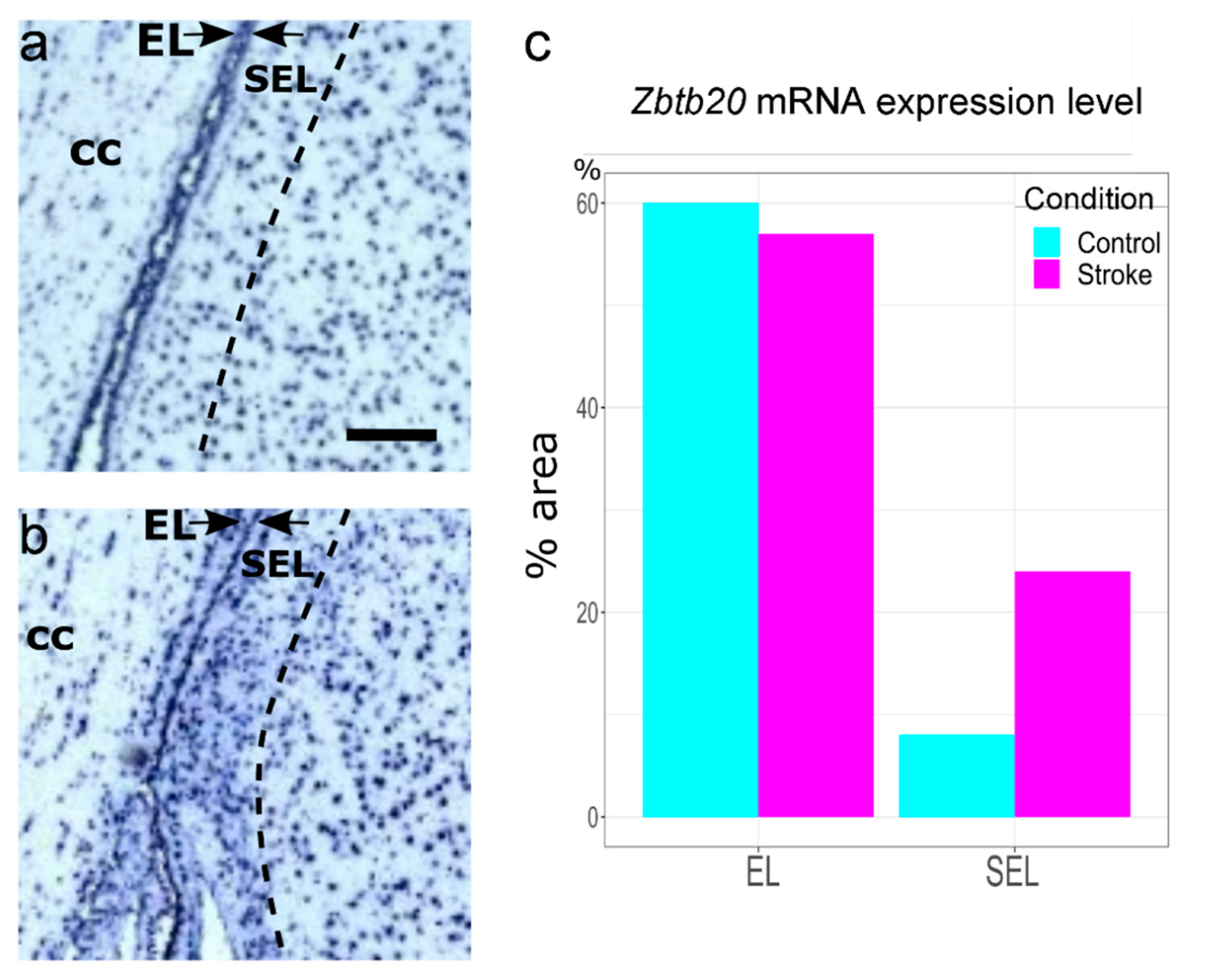
3.4. Enhanced ZBTB20 mRNA Expression in the Adult Monkey SEL following an Ischemic Insult
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obernier, K.; Alvarez-Buylla, A. Neural Stem Cells: Origin, Heterogeneity and Regulation in the Adult Mammalian Brain. Development 2019, 146, dev156059. [Google Scholar] [CrossRef] [PubMed]
- Jurkowski, M.P.; Bettio, L.K.; Woo, E.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell. Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Pilz, G.-A.; Bottes, S.; Betizeau, M.; Jörg, D.J.; Carta, S.; Simons, B.D.; Helmchen, F.; Jessberger, S. Live Imaging of Neurogenesis in the Adult Mouse Hippocampus. Science 2018, 359, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Abbott, L.C.; Nigussie, F. Adult Neurogenesis in the Mammalian Dentate Gyrus. Anat. Histol. Embryol. 2020, 49, 3–16. [Google Scholar] [CrossRef]
- Anacker, C.; Hen, R. Adult Hippocampal Neurogenesis and Cognitive Flexibility—Linking Memory and Mood. Nat. Rev. Neurosci. 2017, 18, 335–346. [Google Scholar] [CrossRef]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the Adult Hippocampus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018812. [Google Scholar] [CrossRef]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional Neurogenesis in the Adult Hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef]
- Eriksson, P.S.; Perfilieva, E.; Björk-Eriksson, T.; Alborn, A.-M.; Nordborg, C.; Peterson, D.A.; Gage, F.H. Neurogenesis in the Adult Human Hippocampus. Nat. Med. 1998, 4, 1313–1317. [Google Scholar] [CrossRef]
- Knoth, R.; Singec, I.; Ditter, M.; Pantazis, G.; Capetian, P.; Meyer, R.P.; Horvat, V.; Volk, B.; Kempermann, G. Murine Features of Neurogenesis in the Human Hippocampus across the Lifespan from 0 to 100 Years. PLoS ONE 2010, 5, e8809. [Google Scholar] [CrossRef]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599.e5. [Google Scholar] [CrossRef] [Green Version]
- Dennis, C.V.; Suh, L.S.; Rodriguez, M.L.; Kril, J.J.; Sutherland, G.T. Human Adult Neurogenesis across the Ages: An Immunohistochemical Study. Neuropathol. Appl. Neurobiol. 2016, 42, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, Y. In Vivo Electroporation and Time-Lapse Imaging of the Rostral Migratory Stream in Developing Rodent Brain. Curr. Protoc. Neurosci. 2019, 87, e65. [Google Scholar] [CrossRef] [PubMed]
- Mizrak, D.; Levitin, H.M.; Delgado, A.C.; Crotet, V.; Yuan, J.; Chaker, Z.; Silva-Vargas, V.; Sims, P.A.; Doetsch, F. Single-Cell Analysis of Regional Differences in Adult V-SVZ Neural Stem Cell Lineages. Cell Rep. 2019, 26, 394–406.e5. [Google Scholar] [CrossRef]
- Bressan, C.; Saghatelyan, A. Intrinsic Mechanisms Regulating Neuronal Migration in the Postnatal Brain. Front. Cell. Neurosci. 2021, 14, 620379. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Wienisch, M.; Murthy, V.N. Development and Refinement of Functional Properties of Adult-Born Neurons. Neuron 2017, 96, 883–896.e7. [Google Scholar] [CrossRef] [PubMed]
- Pencea, V.; Bingaman, K.D.; Freedman, L.J.; Luskin, M.B. Neurogenesis in the Subventricular Zone and Rostral Migratory Stream of the Neonatal and Adult Primate Forebrain. Exp. Neurol. 2001, 172, 1–16. [Google Scholar] [CrossRef]
- Sawamoto, K.; Hirota, Y.; Alfaro-Cervello, C.; Soriano-Navarro, M.; He, X.; Hayakawa-Yano, Y.; Yamada, M.; Hikishima, K.; Tabata, H.; Iwanami, A.; et al. Cellular Composition and Organization of the Subventricular Zone and Rostral Migratory Stream in the Adult and Neonatal Common Marmoset Brain. J. Comp. Neurol. 2011, 519, 690–713. [Google Scholar] [CrossRef]
- Kornack, D.R.; Rakic, P. The Generation, Migration, and Differentiation of Olfactory Neurons in the Adult Primate Brain. Proc. Natl. Acad. Sci. USA 2001, 98, 4752–4757. [Google Scholar] [CrossRef]
- Bédard, A.; Lévesque, M.; Bernier, P.J.; Parent, A. The Rostral Migratory Stream in Adult Squirrel Monkeys: Contribution of New Neurons to the Olfactory Tubercle and Involvement of the Antiapoptotic Protein Bcl-2: The Rostral Migratory Stream in Adult Primates. Eur. J. Neurosci. 2002, 16, 1917–1924. [Google Scholar] [CrossRef]
- Wang, C. Identification and Characterization of Neuroblasts in the Subventricular Zone and Rostral Migratory Stream of the Adult Human Brain. Cell Res. 2011, 21, 17. [Google Scholar] [CrossRef] [Green Version]
- Sanai, N.; Tramontin, A.D.; Barbaro, N.M.; Gupta, N.; Kunwar, S.; Lawton, M.T.; McDermott, M.W.; Parsa, A.T.; Verdugo, J.M.-G.; Berger, M.S.; et al. Unique Astrocyte Ribbon in Adult Human Brain Contains Neural Stem Cells but Lacks Chain Migration. Nature 2004, 427, 5. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotin, S.; Duran-Moreno, M.; Belzunegui, S.; Luquin, M.R.; Garcia-Verdugo, J.M. Ultrastructure of the Subventricular Zone in Macaca Fascicularis and Evidence of a Mouse-Like Migratory Stream. J. Comp. Neurol. 2009, 514, 533–554. [Google Scholar] [CrossRef] [PubMed]
- Quiñones-Hinojosa, A.; Chaichana, K. The Human Subventricular Zone: A Source of New Cells and a Potential Source of Brain Tumors. Exp. Neurol. 2007, 205, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Marques, B.L.; Carvalho, G.A.; Freitas, E.M.M.; Chiareli, R.A.; Barbosa, T.G.; Di Araújo, A.G.P.; Nogueira, Y.L.; Ribeiro, R.I.; Parreira, R.C.; Vieira, M.S.; et al. The Role of Neurogenesis in Neurorepair after Ischemic Stroke. Semin. Cell Dev. Biol. 2019, 95, 98–110. [Google Scholar] [CrossRef]
- Tonchev, A.B.; Yamashima, T.; Sawamoto, K.; Okano, H. Transcription Factor Protein Expression Patterns by Neural or Neuronal Progenitor Cells of Adult Monkey Subventricular Zone. Neuroscience 2006, 139, 1355–1367. [Google Scholar] [CrossRef]
- Rosenthal, E.H.; Tonchev, A.B.; Stoykova, A.; Chowdhury, K. Regulation of Archicortical Arealization by the Transcription Factor Zbtb20. Hippocampus 2012, 22, 2144–2156. [Google Scholar] [CrossRef]
- Nielsen, J.V.; Thomassen, M.; Møllgård, K.; Noraberg, J.; Jensen, N.A. Zbtb20 Defines a Hippocampal Neuronal Identity Through Direct Repression of Genes That Control Projection Neuron Development in the Isocortex. Cereb. Cortex 2014, 24, 1216–1229. [Google Scholar] [CrossRef]
- Tonchev, A.B.; Tuoc, T.C.; Rosenthal, E.H.; Studer, M.; Stoykova, A. Zbtb20 Modulates the Sequential Generation of Neuronal Layers in Developing Cortex. Mol. Brain 2016, 9, 65. [Google Scholar] [CrossRef]
- Doeppner, T.R.; Herz, J.; Bähr, M.; Tonchev, A.B.; Stoykova, A. Zbtb20 Regulates Developmental Neurogenesis in the Olfactory Bulb and Gliogenesis After Adult Brain Injury. Mol. Neurobiol. 2019, 56, 567–582. [Google Scholar] [CrossRef]
- Medeiros de Araújo, J.A.; Barão, S.; Mateos-White, I.; Espinosa, A.; Costa, M.R.; Gil-Sanz, C.; Müller, U. ZBTB20 Is Crucial for the Specification of a Subset of Callosal Projection Neurons and Astrocytes in the Mammalian Neocortex. Development 2021, 148, dev196642. [Google Scholar] [CrossRef]
- Wang, A.; Wang, J.; Tian, K.; Huo, D.; Ye, H.; Li, S.; Zhao, C.; Zhang, B.; Zheng, Y.; Xu, L.; et al. An Epigenetic Circuit Controls Neurogenic Programs during Neocortex Development. Development 2021, 148, dev199772. [Google Scholar] [CrossRef] [PubMed]
- Chongtham, M.C.; Wang, H.; Thaller, C.; Hsiao, N.-H.; Vachkov, I.H.; Pavlov, S.P.; Williamson, L.H.; Yamashima, T.; Stoykova, A.; Yan, J.; et al. Transcriptome Response and Spatial Pattern of Gene Expression in the Primate Subventricular Zone Neurogenic Niche After Cerebral Ischemia. Front. Cell Dev. Biol. 2020, 8, 584314. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ginestet, C. Ggplot2: Elegant Graphics for Data Analysis: Book Reviews. J. R. Stat. Soc. Ser. A 2011, 174, 245–246. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Carson, J.P.; Eichele, G.; Chiu, W. A Method for Automated Detection of Gene Expression Required for the Establishment of a Digital Transcriptome-Wide Gene Expression Atlas. J. Microsc. 2005, 217, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Saleem, K.S.; Logothetis, N. A Combined MRI and Histology Atlas of the Rhesus Monkey Brain in Stereotaxic Coordinates; Elsevier/AP: Amsterdam, The Netherlands; Boston, MA, USA, 2012; ISBN 978-0-12-385088-1. [Google Scholar]
- Tonchev, A.B.; Yamashima, T.; Zhao, L.; Okano, H.J.; Okano, H. Proliferation of Neural and Neuronal Progenitors after Global Brain Ischemia in Young Adult Macaque Monkeys. Mol. Cell. Neurosci. 2003, 23, 292–301. [Google Scholar] [CrossRef]
- Koketsu, D.; Furuichi, Y.; Maeda, M.; Matsuoka, N.; Miyamoto, Y.; Hisatsune, T. Increased Number of New Neurons in the Olfactory Bulb and Hippocampus of Adult Non-Human Primates after Focal Ischemia. Exp. Neurol. 2006, 199, 92–102. [Google Scholar] [CrossRef]
- Nagao, M.; Ogata, T.; Sawada, Y.; Gotoh, Y. Zbtb20 Promotes Astrocytogenesis during Neocortical Development. Nat. Commun. 2016, 7, 11102. [Google Scholar] [CrossRef]
- Kan, H.; Huang, Y.; Li, X.; Liu, D.; Chen, J.; Shu, M. Zinc Finger Protein ZBTB20 Is an Independent Prognostic Marker and Promotes Tumor Growth of Human Hepatocellular Carcinoma by Repressing FoxO1. Oncotarget 2016, 7, 14336–14349. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Zhang, M.; Cheng, L.; Zhang, Y.; Wang, X. ZBTB20 Promotes Cell Migration and Invasion of Gastric Cancer by Inhibiting IκBα to Induce NF-ΚB Activation. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3862–3872. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, K.; Tang, J. Zinc Finger Protein ZBTB20 Promotes Cell Proliferation in Non-Small Cell Lung Cancer through Repression of FoxO1. FEBS Lett. 2014, 588, 4536–4542. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shi, J.-H.; Jiang, H.; Wang, K.; Lu, J.-Y.; Jiang, X.; Ma, X.; Chen, Y.-X.; Ren, A.-J.; Zheng, J.; et al. ZBTB20 Regulates EGFR Expression and Hepatocyte Proliferation in Mouse Liver Regeneration. Cell Death Dis. 2018, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Romano, R.; Bucci, C. Role of EGFR in the Nervous System. Cells 2020, 9, 1887. [Google Scholar] [CrossRef]
- Llorens-Bobadilla, E.; Zhao, S.; Baser, A.; Saiz-Castro, G.; Zwadlo, K.; Martin-Villalba, A. Single-Cell Transcriptomics Reveals a Population of Dormant Neural Stem Cells That Become Activated upon Brain Injury. Cell Stem Cell 2015, 17, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Codega, P.; Silva-Vargas, V.; Paul, A.; Maldonado-Soto, A.R.; DeLeo, A.M.; Pastrana, E.; Doetsch, F. Prospective Identification and Purification of Quiescent Adult Neural Stem Cells from Their In Vivo Niche. Neuron 2014, 82, 545–559. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoyanov, D.S.; Ivanov, M.N.; Yamashima, T.; Tonchev, A.B. Expression of Transcription Factor ZBTB20 in the Adult Primate Neurogenic Niche under Physiological Conditions or after Ischemia. Genes 2022, 13, 1559. https://doi.org/10.3390/genes13091559
Stoyanov DS, Ivanov MN, Yamashima T, Tonchev AB. Expression of Transcription Factor ZBTB20 in the Adult Primate Neurogenic Niche under Physiological Conditions or after Ischemia. Genes. 2022; 13(9):1559. https://doi.org/10.3390/genes13091559
Chicago/Turabian StyleStoyanov, Dimo S., Martin N. Ivanov, Tetsumori Yamashima, and Anton B. Tonchev. 2022. "Expression of Transcription Factor ZBTB20 in the Adult Primate Neurogenic Niche under Physiological Conditions or after Ischemia" Genes 13, no. 9: 1559. https://doi.org/10.3390/genes13091559
APA StyleStoyanov, D. S., Ivanov, M. N., Yamashima, T., & Tonchev, A. B. (2022). Expression of Transcription Factor ZBTB20 in the Adult Primate Neurogenic Niche under Physiological Conditions or after Ischemia. Genes, 13(9), 1559. https://doi.org/10.3390/genes13091559