
The Gain and Loss of Cryptochrome/Photolyase Family Members during Evolution


Abstract
:1. Introduction
1.1. The Cryptochrome/Photolyase Family (CRY/PL Family)
1.2. 6-4 & CPD Photolyases (6-4 PL & CPD PL)
1.3. Plant-Type CRY (PCRY)
1.4. Animal Cryptochromes: Drosophila-Type & Mammalian-Type CRY (DCRY & MCRY)
1.5. DASH-CRY
1.6. Other CRY/PL Family Members (PCRY-Like, CRY4, and PPL)
1.7. Aim of This Project
2. Materials and Methods
2.1. Tree of Life
2.2. BLASTP
2.3. Phylogenetic Tree
2.4. Identification of Cryptochrome/Photolyase Subfamilies
2.5. Characterization of CRY/PL Family Sequences via Their Protein Motifs
2.6. Assignment of CRY/PL Sequences to Their Subfamilies and Organisms
2.7. Figures
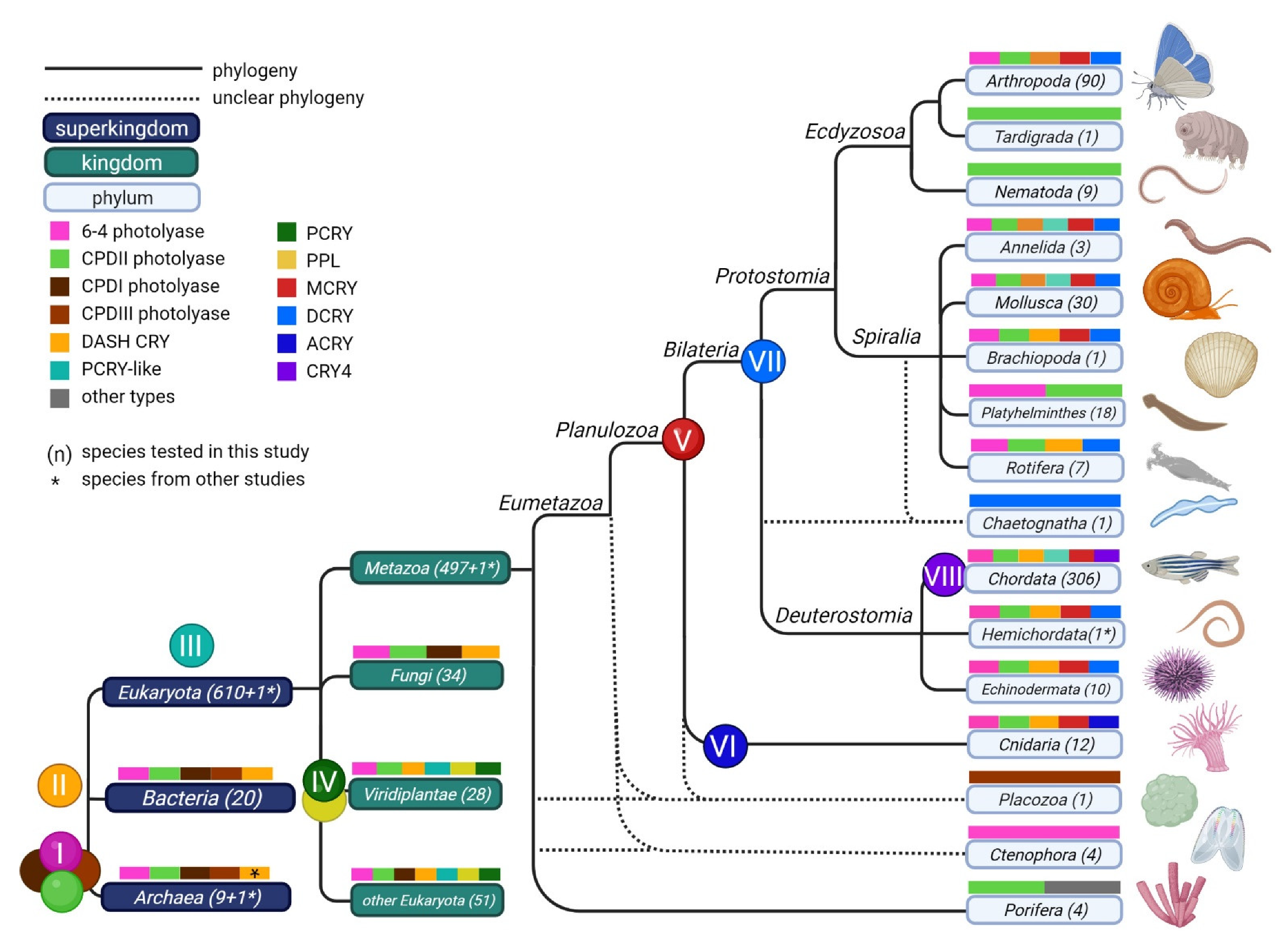
3. Results
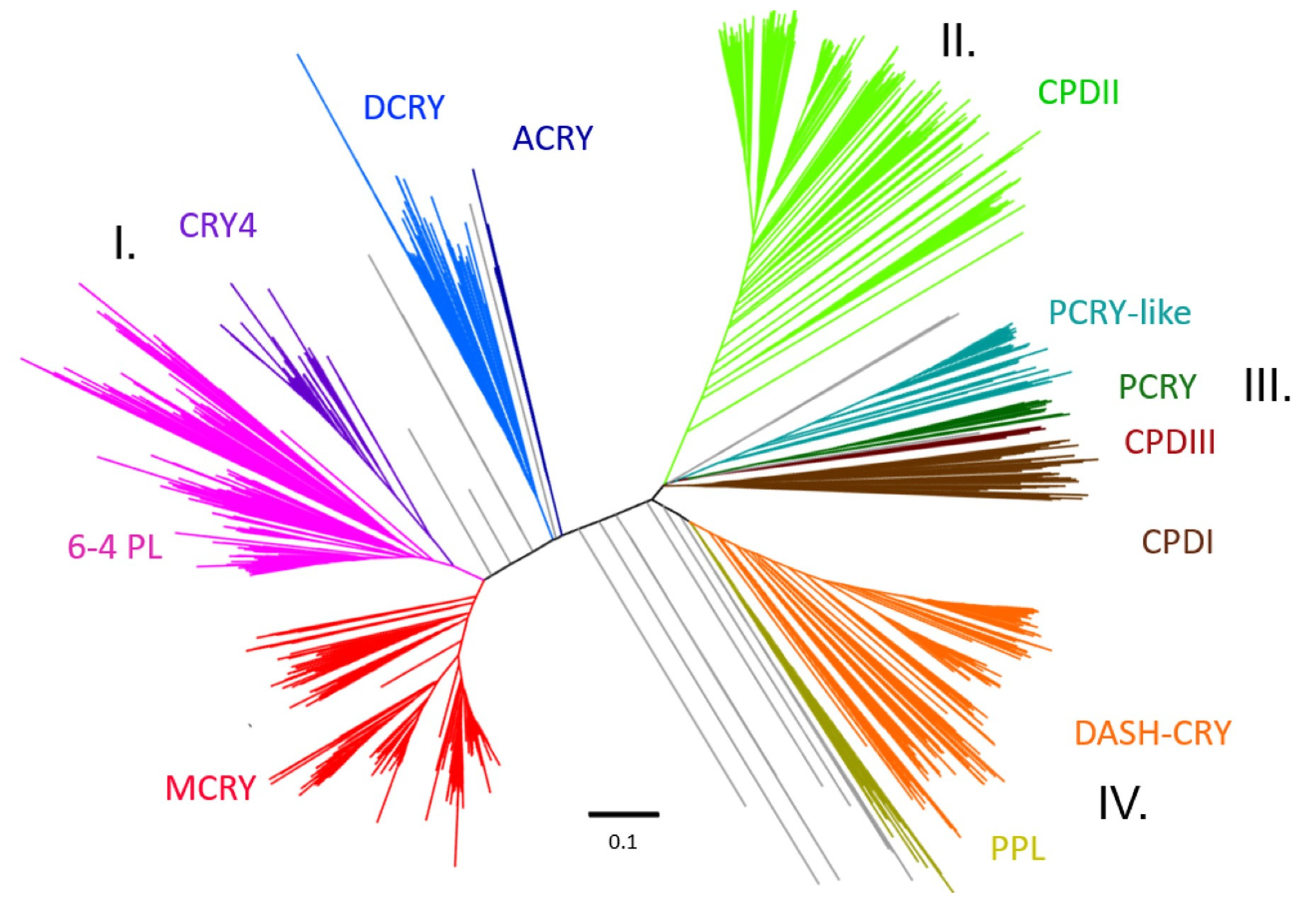
3.1. Phylogenetic Tree
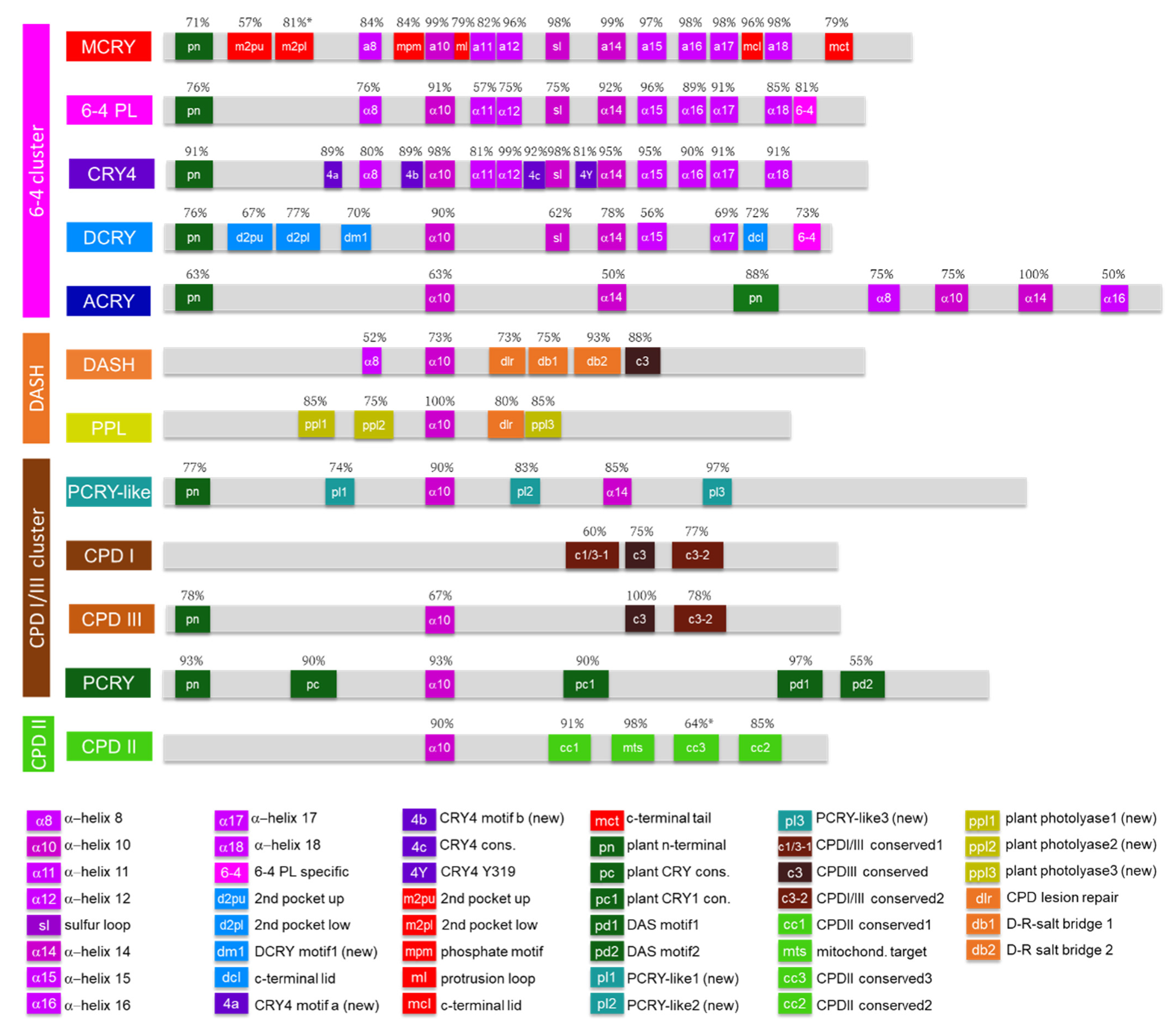
3.2. Motif Analyses
3.2.1. Novel Motifs Enable Strict Differentiation between 6-4 PLs and CRY4s
3.2.2. MCRYs Are Highly Conserved
3.2.3. DCRY and ACRY Differ Greatly in Their Motifs
3.2.4. CPDI and CPDIII PLs Are Relatively Similar, Whereas CPDII PL Differ Greatly
3.2.5. PCRY-Likes Lack PCRY-Specific Motifs, but Possess Three Novel Characteristic Motifs
3.2.6. Novel PPL Motifs Allow Strict Discrimination between DASH-CRY and PPL
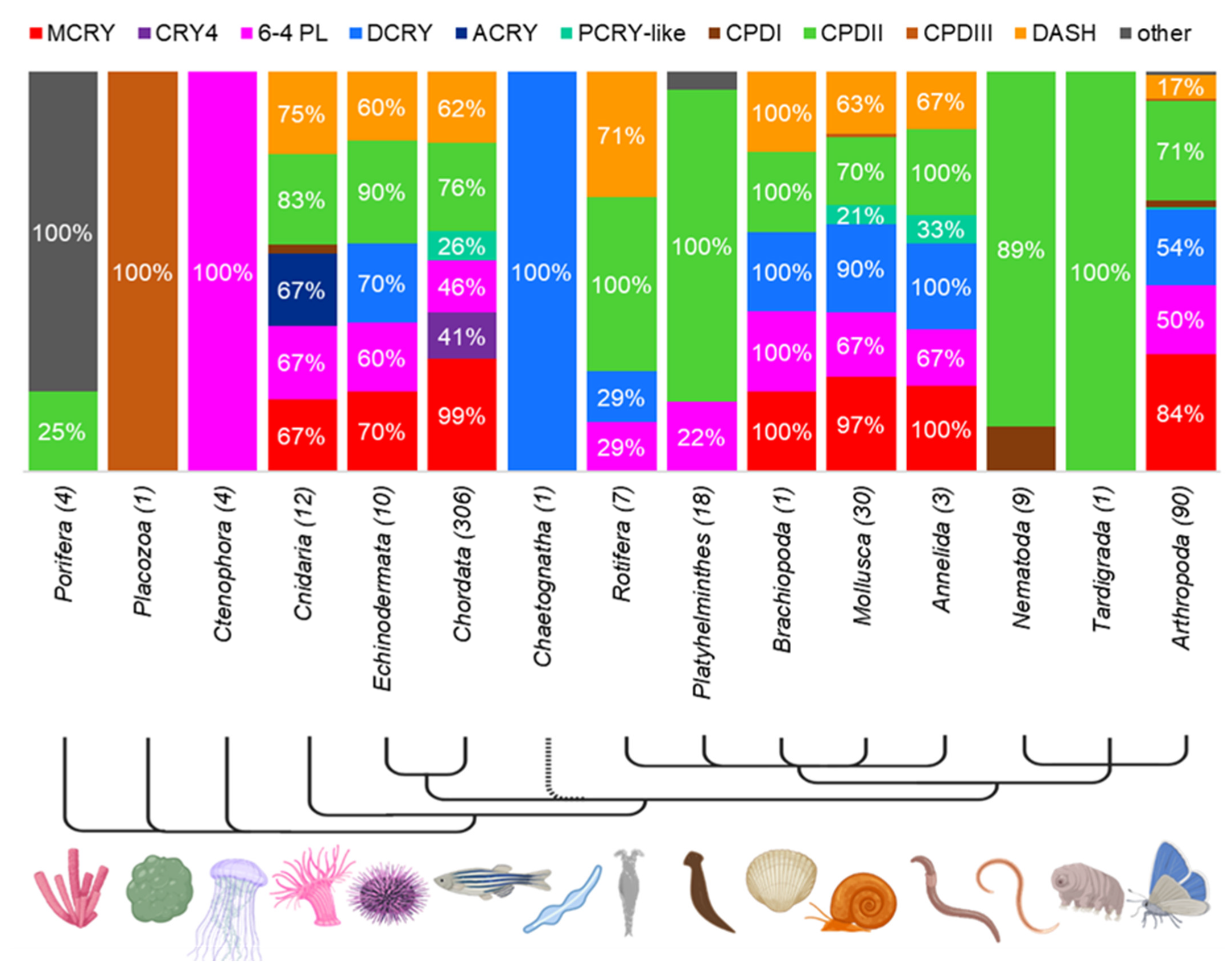
3.3. Distribution of CRY/PL Family Members within the (Super-)Kingdoms
3.4. Prevalence of CRY/PL Family Members within the Metazoa
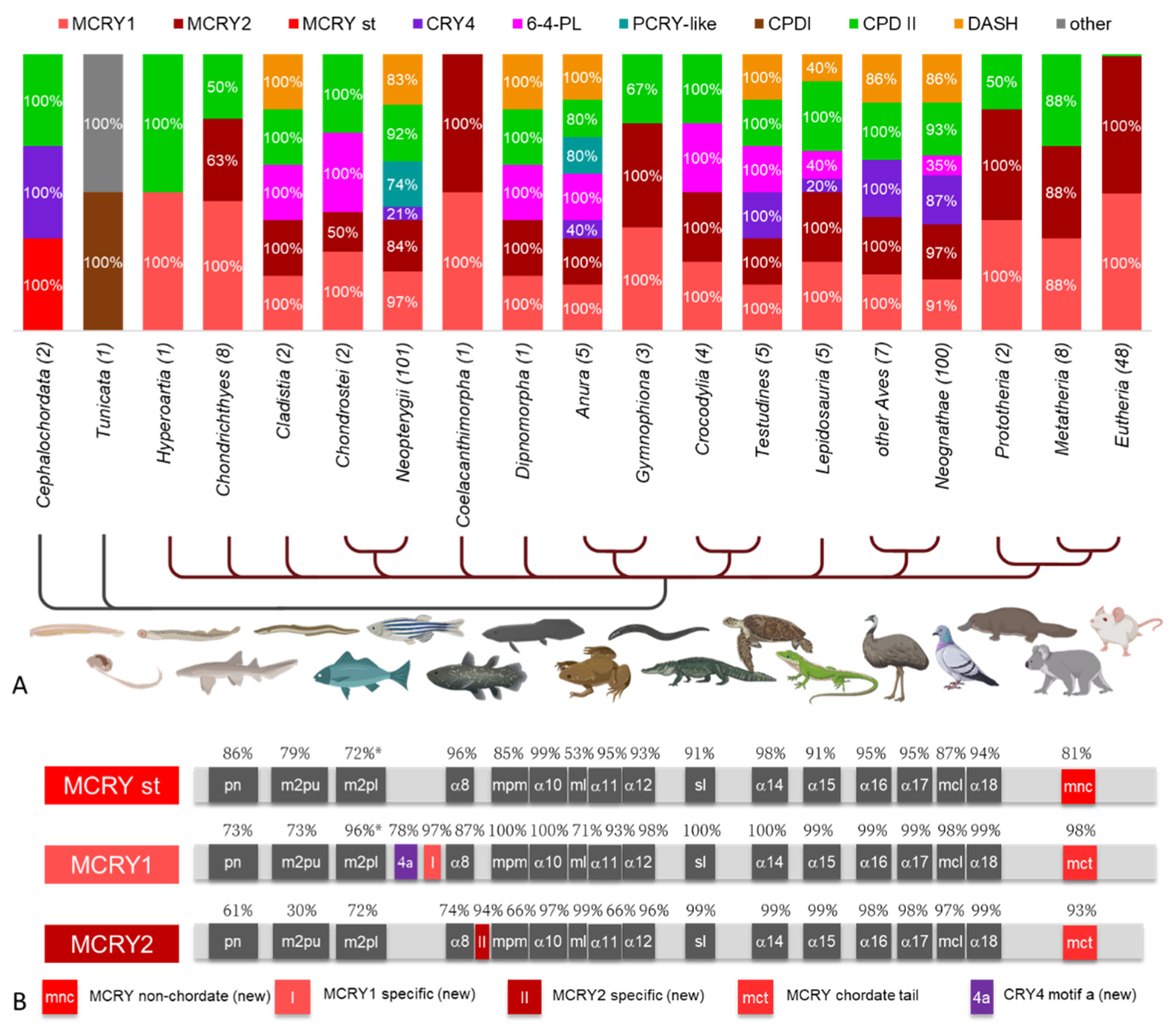
3.5. Prevalence of CRY/PL Family Members within Chordata
3.6. Prevalence of Members of the CRY/PL Family within the Phylum Arthropoda
4. Discussion
4.1. Genesis of the CRY/PL Family Members
4.1.1. Photolyases Are Ancient Molecules Found in All Three Kingdoms
4.1.2. DASH-CRY: The First Cryptochrome or Still a Photolyase?
4.1.3. Plant CRY-Like (PCRY-Like)
4.1.4. Plant CRY (PCRY) and Plant Photolyase (PPL)
4.2. The Rise of Animal Cryptochromes
4.2.1. Original Metazoans Strongly Differ in Their CRY/PL
4.2.2. Cnidarian: Animal CRY Expansion > Birth of MCRY and ACRY
4.2.3. MCRY Is More Ancient (Already in Planulozoa), DCRY Is Only Found in Bilateria
4.2.4. CRY4: The Chordate-Specific Magnetoreceptor
4.2.5. Animal CRYs in Protostomia: Spiralia & Ecdyzosoa
4.3. Selection and Restriction to Certain CRY/PL Family Members
4.3.1. Echinoderms share multiple CRY/PLs, including DCRY
4.3.2. The Original Chordates Have a Reduced CRY/PL Repertoire
4.3.3. CRY/PLs in Bony Fishes, Coelacanths and in the Lungfish
4.3.4. CRY/PL Distribution among Tetrapods
4.3.5. Highly Reduced CRY/PLs in Mammals
4.3.6. The Arrow Worm Chaetognata Only Feature DCRY
4.3.7. Enriched CRY/PLs in Mollusks and in Annelids
4.3.8. Strong Reduction of CRY/PLs in Platyhelminths & Nematodes
4.3.9. Arthropoda: Varying Distribution of CRY/PL Members within the Chelicerata
4.3.10. DASH-CRY Is Quite Common in Crustacea
4.3.11. MCRY, DCRY, 6-4 and CPDII Photolyases Are the Hexapod Typical CRY/PLs
4.4. The Sudden Appearance and Disappearance of CRY/PL Members within Single Taxa
4.4.1. Gene Duplication & Gene Loss
4.4.2. Horizontal Gene Transfer (HGT) & Microbiome
4.4.3. Do Ancient Animals Have More CRY/PL Family Members Than the Recent Ones?
4.4.4. Methodical Errors: Incomplete Annotations & Possible DNA Contamination
4.5. When and Why Did the Repair Enzyme Turn into a Clock Protein?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ozturk, N. Phylogenetic and Functional Classification of the Photolyase/Cryptochrome Family. Photochem. Photobiol. 2017, 93, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Sancar, A. Structure and Function of DNA Photolyase and Cryptochrome Blue-Light Photoreceptors. Chem. Rev. 2003, 103, 2203–2238. [Google Scholar] [CrossRef] [PubMed]
- Menck, C.F.M. Shining a Light on Photolyases. Nat. Genet. 2002, 32, 338–339. [Google Scholar] [CrossRef]
- Sancar, A. Mechanisms of DNA Repair by Photolyase and Excision Nuclease (Nobel Lecture). Angew. Chem. Int. Ed. 2016, 55, 8502–8527. [Google Scholar] [CrossRef] [PubMed]
- Kavakli, I.H.; Baris, I.; Tardu, M.; Gül, Ş.; Öner, H.; Çal, S.; Bulut, S.; Yarparvar, D.; Berkel, Ç.; Ustaoğlu, P.; et al. The Photolyase/Cryptochrome Family of Proteins as DNA Repair Enzymes and Transcriptional Repressors. Photochem. Photobiol. 2017, 93, 93–103. [Google Scholar] [CrossRef]
- Todo, T.; Takemori, H.; Ryo, H.; Lhara, M.; Matsunaga, T.; Nikaido, O.; Sato, K.; Nomura, T. A New Photoreactivating Enzyme That Specifically Repairs Ultraviolet Light-Induced (6-4)Photoproducts. Nature 1993, 361, 371–374. [Google Scholar] [CrossRef]
- Boyd, J.B.; Harris, P.V. Isolation and Characterization of a Photorepair-Deficient Mutant in Drosophila melanogaster. Genetics 1987, 116, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, N.; Kao, Y.-T.; Selby, C.P.; Kavaklı, I.H.; Partch, C.L.; Zhong, D.; Sancar, A. Purification and Characterization of a Type III Photolyase from Caulobacter Crescentus. Biochemistry 2008, 47, 10255–10261. [Google Scholar] [CrossRef]
- Kiontke, S.; Geisselbrecht, Y.; Pokorny, R.; Carell, T.; Batschauer, A.; Essen, L.O. Crystal Structures of an Archaeal Class II DNA Photolyase and Its Complex with UV-Damaged Duplex DNA. EMBO J. 2011, 30, 4437–4449. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, L.; Zhong, D. Photolyase: Dynamics and Mechanisms of Repair of Sun-Induced DNA Damage. Photochem. Photobiol. 2017, 93, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Cashmore, A.R. HY4 Gene of A. Thaliana Encodes a Protein with Characteristics of a Blue-Light Photoreceptor. Nature 1993, 366, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, B.; Su, J.; Liao, J.; Lin, C.; Oka, Y. Cryptochromes Orchestrate Transcription Regulation of Diverse Blue Light Responses in Plants. Photochem. Photobiol. 2017, 93, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Dissel, S.; Codd, V.; Fedic, R.; Garner, K.J.; Costa, R.; Kyriacou, C.P.; Rosato, E. A Constitutively Active Cryptochrome in Drosophila melanogaster. Nat. Neurosci. 2004, 7, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Ceriani, M.F.; Darlington, T.K.; Staknis, D.; Más, P.; Petti, A.A.; Weitz, C.J.; Kay, S.A. Light-Dependent Sequestration of TIMELESS by CRYPTOCHROME. Science 1999, 285, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, N.; VanVickle-Chavez, S.J.; Akileswaran, L.; Gelder, R.N.V.; Sancar, A. Ramshackle (Brwd3) Promotes Light-Induced Ubiquitylation of Drosophila Cryptochrome by DDB1-CUL4-ROC1 E3 Ligase Complex. Proc. Natl. Acad. Sci. USA 2013, 110, 4980–4985. [Google Scholar] [CrossRef] [PubMed]
- Peschel, N.; Chen, K.F.; Szabo, G.; Stanewsky, R. Light-Dependent Interactions between the Drosophila Circadian Clock Factors Cryptochrome, Jetlag, and Timeless. Curr. Biol. 2009, 19, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Todo, T.; Ryo, H.; Yamamoto, K.; Toh, H.; Inui, T.; Ayaki, H.; Nomura, T.; Ikenaga, M. Similarity among the Drosophila (6-4)Photolyase, a Human Photolyase Homolog, and the DNA Photolyase-Blue-Light Photoreceptor Family. Science 1996, 272, 109–112. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Sancar, A. Vitamin B2-Based Blue-Light Photoreceptors in the Retinohypothalamic Tract as the Photoactive Pigments for Setting the Circadian Clock in Mammals. Proc. Natl. Acad. Sci. USA 1998, 95, 6097–6102. [Google Scholar] [CrossRef]
- Mei, Q.; Dvornyk, V. Evolutionary History of the Photolyase/Cryptochrome Superfamily in Eukaryotes. PLoS ONE 2015, 10, e0135940. [Google Scholar] [CrossRef]
- Miles, J.A.; Davies, T.A.; Hayman, R.D.; Lorenzen, G.; Taylor, J.; Anjarwalla, M.; Allen, S.J.R.; Graham, J.W.D.; Taylor, P.C. A Case Study of Eukaryogenesis: The Evolution of Photoreception by Photolyase/Cryptochrome Proteins. J. Mol. Evol. 2020, 88, 662–673. [Google Scholar] [CrossRef]
- Vechtomova, Y.L.; Telegina, T.A.; Kritsky, M.S. Evolution of Proteins of the DNA Photolyase/Cryptochrome Family. Biochemistry 2020, 85, S131–S153. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K.; Okamoto, K.; Daiyasu, H.; Miyashita, H.; Iwai, S.; Toh, H.; Ishiura, M.; Todo, T. Bacterial Cryptochrome and Photolyase: Characterization of Two Photolyase-like Genes of Synechocystis Sp. PCC6803. Nucleic Acids Res. 2000, 28, 2353–2362. [Google Scholar] [CrossRef] [PubMed]
- Kiontke, S.; Göbel, T.; Brych, A.; Batschauer, A. DASH-Type Cryptochromes—Solved and Open Questions. Biol. Chem. 2020, 401, 1487–1493. [Google Scholar] [CrossRef] [PubMed]
- Brudler, R.; Hitomi, K.; Daiyasu, H.; Toh, H.; Kucho, K.; Ishiura, M.; Kanehisa, M.; Roberts, V.A.; Todo, T.; Tainer, J.A.; et al. Identification of a New Cryptochrome Class: Structure, Function, and Evolution. Mol. Cell 2003, 11, 59–67. [Google Scholar] [CrossRef]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutiérrez, G.; Miralles-Durán, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal Cryptochrome with DNA Repair Activity Reveals an Early Stage in Cryptochrome Evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 15130–15135. [Google Scholar] [CrossRef]
- Navarro, E.; Niemann, N.; Kock, D.; Dadaeva, T.; Gutiérrez, G.; Engelsdorf, T.; Kiontke, S.; Corrochano, L.M.; Batschauer, A.; Garre, V. The DASH-Type Cryptochrome from the Fungus Mucor Circinelloides Is a Canonical CPD-Photolyase. Curr. Biol. 2020, 30, 4483–4490.e4. [Google Scholar] [CrossRef]
- Nsa, I.Y.; Karunarathna, N.; Liu, X.; Huang, H.; Boetteger, B.; Bell-Pedersen, D. A Novel Cryptochrome-Dependent Oscillator in Neurospora Crassa. Genetics 2015, 199, 233–245. [Google Scholar] [CrossRef]
- Juhas, M.; von Zadow, A.; Spexard, M.; Schmidt, M.; Kottke, T.; Büchel, C. A Novel Cryptochrome in the Diatom Phaeodactylum Tricornutum Influences the Regulation of Light-Harvesting Protein Levels. FEBS J. 2014, 281, 2299–2311. [Google Scholar] [CrossRef]
- Oliveri, P.; Fortunato, A.E.; Petrone, L.; Ishikawa-Fujiwara, T.; Kobayashi, Y.; Todo, T.; Antonova, O.; Arboleda, E.; Zantke, J.; Tessmar-Raible, K.; et al. The Cryptochrome/Photolyase Family in Aquatic Organisms. Mar. Genom. 2014, 14, 23–37. [Google Scholar] [CrossRef]
- Fortunato, A.E.; Annunziata, R.; Jaubert, M.; Bouly, J.P.; Falciatore, A. Dealing with Light: The Widespread and Multitasking Cryptochrome/Photolyase Family in Photosynthetic Organisms. J. Plant Physiol. 2015, 172, 42–54. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Ishikawa, T.; Hirayama, J.; Daiyasu, H.; Kanai, S.; Toh, H.; Fukuda, I.; Tsujimura, T.; Terada, N.; Kamei, Y.; et al. Molecular Analysis of Zebrafish Photolyase/Cryptochrome Family: Two Types of Cryptochromes Present in Zebrafish. Genes Cells 2000, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Hochstoeger, T.; Said, T.A.; Maestre, D.; Walter, F.; Vilceanu, A.; Pedron, M.; Cushion, T.D.; Snider, W.; Nimpf, S.; Nordmann, G.C.; et al. The Biophysical, Molecular, and Anatomical Landscape of Pigeon CRY4: A Candidate Light-Based Quantal Magnetosensor. Sci. Adv. 2020, 6, eabb9110. [Google Scholar] [CrossRef] [PubMed]
- Xie, C. Searching for Unity in Diversity of Animal Magnetoreception: From Biology to Quantum Mechanics and Back. Innovation 2022, 3, 100229. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.L.; Lang, D.W.; Small, G.D. Cloning and Characterization of a Class II DNA Photolyase from Chlamydomonas. Plant Mol. Biol. 1999, 40, 1063–1071. [Google Scholar] [CrossRef]
- Gentile, C.; Rivas, G.; Meireles-Filho, A.; Lima, J.; Peixoto, A. Circadian Expression of Clock Genes in Two Mosquito Disease Vectors: Cry2 Is Different. J. Biol. Rhythm. 2009, 24, 444–451. [Google Scholar] [CrossRef]
- Kiontke, S.; Gnau, P.; Haselsberger, R.; Batschauer, A.; Essen, L.-O. Structural and Evolutionary Aspects of Antenna Chromophore Usage by Class II Photolyases. J. Biol. Chem. 2014, 289, 19659–19669. [Google Scholar] [CrossRef]
- Rubin, E.B.; Shemesh, Y.; Cohen, M.; Elgavish, S.; Robertson, H.M.; Bloch, G. Molecular and Phylogenetic Analyses Reveal Mammalian-like Clockwork in the Honey Bee (Apis mellifera) and Shed New Light on the Molecular Evolution of the Circadian Clock. Genome Res. 2006, 16, 1352–1365. [Google Scholar] [CrossRef]
- Haug, M.F.; Gesemann, M.; Lazović, V.; Neuhauss, S.C. Eumetazoan Cryptochrome Phylogeny and Evolution. Genome Biol. Evol. 2015, 7, 601–619. [Google Scholar] [CrossRef]
- Nitta, Y.; Matsui, S.; Kato, Y.; Kaga, Y.; Sugimoto, K.; Sugie, A. Analysing the Evolutional and Functional Differentiation of Four Types of Daphnia Magna Cryptochrome in Drosophila Circadian Clock. Sci. Rep. 2019, 9, 8857. [Google Scholar] [CrossRef]
- Gornik, S.G.; Bergheim, B.G.; Morel, B.; Stamatakis, A.; Foulkes, N.S.; Guse, A. Photoreceptor Diversification Accompanies the Evolution of Anthozoa. Mol. Biol. Evol. 2021, 38, 1744–1760. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.; Bradnam, K.; John, J.S.; Darling, A.; Lin, D.; Fass, J.; Yu, H.O.K.; Buffalo, V.; Zerbino, D.R.; Diekhans, M.; et al. Assemblathon 1: A Competitive Assessment of de Novo Short Read Assembly Methods. Genome Res. 2011, 21, 2224–2241. [Google Scholar] [CrossRef]
- Kotwica-Rolinska, J.; Chodáková, L.; Smýkal, V.; Damulewicz, M.; Provazník, J.; Wu, B.C.-H.; Hejníková, M.; Chvalová, D.; Doležel, D. Loss of Timeless Underlies an Evolutionary Transition within the Circadian Clock. Mol. Biol. Evol. 2021, 39, msab346. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, N. Light-Dependent Reactions of Animal Circadian Photoreceptor Cryptochrome. FEBS J. 2021. [CrossRef] [PubMed]
- Eisen, J.A.; Hanawalt, P.C. A Phylogenomic Study of DNA Repair Genes, Proteins, and Processes. Mutat. Res./DNA Repair 1999, 435, 171–213. [Google Scholar] [CrossRef]
- Lucas-Lledó, J.I.; Lynch, M. Evolution of Mutation Rates: Phylogenomic Analysis of the Photolyase/Cryptochrome Family. Mol. Biol. Evol. 2009, 26, 1143–1153. [Google Scholar] [CrossRef]
- Cockell, C.S. A Photobiological History of Earth. In Ecosystems, Evolution, and Ultraviolet Radiation; Cockell, C.S., Blaustein, A.R., Eds.; Springer: New York, NY, USA, 2001; pp. 1–35. ISBN 978-1-4757-3486-7. [Google Scholar]
- Zhang, F.; Scheerer, P.; Oberpichler, I.; Lamparter, T.; Krauß, N. Crystal Structure of a Prokaryotic (6-4) Photolyase with an Fe-S Cluster and a 6,7-Dimethyl-8-Ribityllumazine Antenna Chromophore. Proc. Natl. Acad. Sci. USA 2013, 110, 7217–7222. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.; Wen, B.; Shi, H.; Chi, C.; Liu, C.; Wang, K.; Tao, X.; Wang, M.; Lv, J.; et al. Identification of a Novel Class of Photolyases as Possible Ancestors of Their Family. Mol. Biol. Evol. 2021, 38, 4505–4519. [Google Scholar] [CrossRef]
- Castrillo, M.; García-Martínez, J.; Avalos, J. Light-Dependent Functions of the Fusarium Fujikuroi CryD DASH Cryptochrome in Development and Secondary Metabolism. Appl. Environ. Microbiol. 2013, 79, 2777–2788. [Google Scholar] [CrossRef]
- Veluchamy, S.; Rollins, J.A. A CRY-DASH-Type Photolyase/Cryptochrome from Sclerotinia sclerotiorum Mediates Minor UV-A-Specific Effects on Development. Fungal Genet. Biol. 2008, 45, 1265–1276. [Google Scholar] [CrossRef]
- Rredhi, A.; Petersen, J.; Schubert, M.; Li, W.; Oldemeyer, S.; Li, W.; Westermann, M.; Wagner, V.; Kottke, T.; Mittag, M. DASH Cryptochrome 1, a UV-A Receptor, Balances the Photosynthetic Machinery of Chlamydomonas reinhardtii. New Phytol. 2021, 232, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Daiyasu, H.; Ishikawa, T.; Kuma, K.; Iwai, S.; Todo, T.; Toh, H. Identification of Cryptochrome DASH from Vertebrates. Genes Cells 2004, 9, 479–495. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Baxter, R.; Smith, B.S.; Partch, C.L.; Colbert, C.L.; Deisenhofer, J. Crystal Structure of Cryptochrome 3 from Arabidopsis thaliana and Its Implications for Photolyase Activity. Proc. Natl. Acad. Sci. USA 2006, 103, 17701–17706. [Google Scholar] [CrossRef] [PubMed]
- Selby, C.P.; Sancar, A. A Cryptochrome/Photolyase Class of Enzymes with Single-Stranded DNA-Specific Photolyase Activity. Proc. Natl. Acad. Sci. USA 2006, 103, 17696–17700. [Google Scholar] [CrossRef]
- Drudge, C.N.; Warren, L.A. Prokaryotic Horizontal Gene Transfer in Freshwater Lakes: Implications of Dynamic Biogeochemical Zonation. J. Environ. Prot. 2012, 3, 1634–1654. [Google Scholar] [CrossRef]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot Spots of Horizontal Gene Transfer (HGT) in Aquatic Environments, with a Focus on a New HGT Mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Darwin, C.; Darwin, F. The Power of Movement in Plants; D. Appleton and Company: New York, NY, USA, 1881. [Google Scholar]
- Koornneef, M.; Rolff, E.; Spruit, C.J.P. Genetic Control of Light-Inhibited Hypocotyl Elongation in Arabidopsis thaliana (L.) Heynh. Zeitschrift Pflanzenphysiologie 1980, 100, 147–160. [Google Scholar] [CrossRef]
- Lin, C.; Ahmad, M.; Gordon, D.; Cashmore, A.R. Expression of an Arabidopsis Cryptochrome Gene in Transgenic Tobacco Results in Hypersensitivity to Blue, UV-A, and Green Light. Proc. Natl. Acad. Sci. USA 1995, 92, 8423–8427. [Google Scholar] [CrossRef]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of Flowering Time by Arabidopsis Photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef]
- Somers, D.E.; Devlin, P.F.; Kay, S.A. Phytochromes and Cryptochromes in the Entrainment of the Arabidopsis Circadian Clock. Science 1998, 282, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Scheerer, P.; Zhang, F.; Kalms, J.; von Stetten, D.; Krauß, N.; Oberpichler, I.; Lamparter, T. The Class III Cyclobutane Pyrimidine Dimer Photolyase Structure Reveals a New Antenna Chromophore Binding Site and Alternative Photoreduction Pathways. J. Biol. Chem. 2015, 290, 11504–11514. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Yang, Z.; Gomez, A.; Liu, B.; Lin, C.; Oka, Y. Signaling Mechanisms of Plant Cryptochromes in Arabidopsis thaliana. J. Plant Res. 2016, 129, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zuo, Z.; Wang, X.; Liu, Q.; Gu, L.; Oka, Y.; Lin, C. Beyond the Photocycle-How Cryptochromes Regulate Photoresponses in Plants? Curr. Opin. Plant Biol. 2018, 45, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Metterville, D.; Briscoe, A.D.; Reppert, S.M. Insect Cryptochromes: Gene Duplication and Loss Define Diverse Ways to Construct Insect Circadian Clocks. Mol. Biol. Evol. 2007, 24, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Boero, F.; Schierwater, B.; Piraino, S. Cnidarian Milestones in Metazoan Evolution. Integr. Comp. Biol. 2007, 47, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Daley, A.C.; Antcliffe, J.B. Evolution: The Battle of the First Animals. Curr. Biol. 2019, 29, R257–R259. [Google Scholar] [CrossRef]
- Genikhovich, G.; Technau, U. On the Evolution of Bilaterality. Development 2017, 144, 3392–3404. [Google Scholar] [CrossRef]
- Jékely, G.; Paps, J.; Nielsen, C. The Phylogenetic Position of Ctenophores and the Origin(s) of Nervous Systems. EvoDevo 2015, 6, 1. [Google Scholar] [CrossRef]
- Nielsen, C. Early Animal Evolution: A Morphologist’s View. R. Soc. Open Sci. 2019, 6, 190638. [Google Scholar] [CrossRef]
- King, N.; Rokas, A. Embracing Uncertainty in Reconstructing Early Animal Evolution. Curr. Biol. 2017, 27, R1081–R1088. [Google Scholar] [CrossRef] [Green Version]
- Sebé-Pedrós, A.; Degnan, B.M.; Ruiz-Trillo, I. The Origin of Metazoa: A Unicellular Perspective. Nat. Rev. Genet. 2017, 18, 498–512. [Google Scholar] [CrossRef]
- Laumer, C.E.; Gruber-Vodicka, H.; Hadfield, M.G.; Pearse, V.B.; Riesgo, A.; Marioni, J.C.; Giribet, G. Support for a Clade of Placozoa and Cnidaria in Genes with Minimal Compositional Bias. eLife 2018, 7, e36278. [Google Scholar] [CrossRef] [PubMed]
- Whelan, N.V.; Kocot, K.M.; Moroz, L.L.; Halanych, K.M. Error, Signal, and the Placement of Ctenophora Sister to All Other Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5773–5778. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.S.; Ozturk, N.; Fahey, B.; Plachetzki, D.C.; Degnan, B.M.; Sancar, A.; Oakley, T.H. Blue-Light-Receptive Cryptochrome Is Expressed in a Sponge Eye Lacking Neurons and Opsin. J. Exp. Biol. 2012, 215, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Schröder, H.C.; Markl, J.S.; Grebenjuk, V.A.; Korzhev, M.; Steffen, R.; Wang, X. Cryptochrome in Sponges. J. Histochem. Cytochem. 2013, 61, 814–832. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Begovic, E.; Chapman, J.; Putnam, N.H.; Hellsten, U.; Kawashima, T.; Kuo, A.; Mitros, T.; Salamov, A.; Carpenter, M.L.; et al. The Trichoplax Genome and the Nature of Placozoans. Nature 2008, 454, 955–960. [Google Scholar] [CrossRef]
- Dohrmann, M.; Wörheide, G. Novel Scenarios of Early Animal Evolution—Is It Time to Rewrite Textbooks? Integr. Comp. Biol. 2013, 53, 503–511. [Google Scholar] [CrossRef]
- Reitzel, A.; Tarrant, A.; Levy, O. Circadian Clocks in the Cnidaria: Environmental Entrainment, Molecular Regulation, and Organismal Outputs. Integr. Comp. Biol. 2013, 53, 118–130. [Google Scholar] [CrossRef]
- Rutila, J.E.; Suri, V.; Le, M.; So, W.V.; Rosbash, M.; Hall, J.C. CYCLE Is a Second BHLH-PAS Clock Protein Essential for Circadian Rhythmicity and Transcription of Drosophila period and timeless. Cell 1998, 93, 805–814. [Google Scholar] [CrossRef]
- Sebé-Pedrós, A.; Chomsky, E.; Pang, K.; Lara-Astiaso, D.; Gaiti, F.; Mukamel, Z.; Amit, I.; Hejnol, A.; Degnan, B.M.; Tanay, A. Early Metazoan Cell Type Diversity and the Evolution of Multicellular Gene Regulation. Nat. Ecol. Evol. 2018, 2, 1176–1188. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Chelliah, Y.; Wickramaratne, A.; Jarocha, L.; Karki, N.; Xu, W.; Mouritsen, H.; Hore, P.J.; Hibbs, R.E.; Green, C.B.; et al. Chemical and Structural Analysis of a Photoactive Vertebrate Cryptochrome from Pigeon. Proc. Natl. Acad. Sci. USA 2019, 116, 19449–19457. [Google Scholar] [CrossRef] [PubMed]
- James, D.G.; Kappen, L. Further Insights on the Migration Biology of Monarch Butterflies, Danaus plexippus (Lepidoptera: Nymphalidae) from the Pacific Northwest. Insects 2021, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Bazalova, O.; Kvicalova, M.; Valkova, T.; Slaby, P.; Bartos, P.; Netusil, R.; Tomanova, K.; Braeunig, P.; Lee, H.-J.; Sauman, I.; et al. Cryptochrome 2 Mediates Directional Magnetoreception in Cockroaches. Proc. Natl. Acad. Sci. USA 2016, 113, 1660–1665. [Google Scholar] [CrossRef] [PubMed]
- Netušil, R.; Tomanová, K.; Chodáková, L.; Chvalová, D.; Doležel, D.; Ritz, T.; Vácha, M. Cryptochrome-Dependent Magnetoreception in a Heteropteran Insect Continues Even after 24 h in Darkness. J. Exp. Biol. 2021, 224, jeb243000. [Google Scholar] [CrossRef] [PubMed]
- Gegear, R.J.; Casselman, A.; Waddell, S.; Reppert, S.M. Cryptochrome Mediates Light-Dependent Magnetosensitivity in Drosophila. Nature 2008, 454, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Marley, R.; Giachello, C.N.G.; Scrutton, N.S.; Baines, R.A.; Jones, A.R. Cryptochrome-Dependent Magnetic Field Effect on Seizure Response in Drosophila larvae. Sci. Rep. 2014, 4, 5799. [Google Scholar] [CrossRef]
- Wu, C.-L.; Fu, T.-F.; Chiang, M.-H.; Chang, Y.-W.; Her, J.-L.; Wu, T. Magnetoreception Regulates Male Courtship Activity in Drosophila. PLoS ONE 2016, 11, e0155942. [Google Scholar] [CrossRef]
- Gegear, R.J.; Foley, L.E.; Casselman, A.; Reppert, S.M. Animal Cryptochromes Mediate Magnetoreception by an Unconventional Photochemical Mechanism. Nature 2010, 463, 804. [Google Scholar] [CrossRef]
- Wan, G.; Hayden, A.N.; Iiams, S.E.; Merlin, C. Cryptochrome 1 Mediates Light-Dependent Inclination Magnetosensing in Monarch Butterflies. Nat. Commun. 2021, 12, 771. [Google Scholar] [CrossRef]
- Isely, N.; Lamare, M.; Marshall, C.; Barker, M. Expression of the DNA Repair Enzyme, Photolyase, in Developmental Tissues and Larvae, and in Response to Ambient UV-R in the Antarctic Sea Urchin Sterechinus neumayeri. Photochem. Photobiol. 2009, 85, 1168–1176. [Google Scholar] [CrossRef]
- Helfer, G.; Fidler, A.E.; Vallone, D.; Foulkes, N.S.; Brandstaetter, R. Molecular Analysis of Clock Gene Expression in the Avian Brain. Chronobiol. Int. 2006, 23, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Kubo, Y.; Akiyama, M.; Fukada, Y.; Okano, T. Molecular Cloning, MRNA Expression, and Immunocytochemical Localization of a Putative Blue-Light Photoreceptor CRY4 in the Chicken Pineal Gland. J. Neurochem. 2006, 97, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Kubo, Y.; Okano, K.; Okano, T. Identification and Characterization of Cryptochrome4 in the Ovary of Western Clawed Frog Xenopus tropicalis. Zool. Sci. 2014, 31, 152–159. [Google Scholar] [CrossRef]
- Liu, C.; Hu, J.; Qu, C.; Wang, L.; Huang, G.; Niu, P.; Zhong, Z.; Hong, F.; Wang, G.; Postlethwait, J.H.; et al. Molecular Evolution and Functional Divergence of Zebrafish (Danio rerio) Cryptochrome Genes. Sci. Rep. 2015, 5, 8113. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, W.; Wiltschko, R. Magnetic Compass of European Robins. Science 1972, 176, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Putman, N.F.; Endres, C.S.; Lohmann, C.M.F.; Lohmann, K.J. Longitude Perception and Bicoordinate Magnetic Maps in Sea Turtles. Curr. Biol. 2011, 21, 463–466. [Google Scholar] [CrossRef]
- Bottesch, M.; Gerlach, G.; Halbach, M.; Bally, A.; Kingsford, M.J.; Mouritsen, H. A Magnetic Compass That Might Help Coral Reef Fish Larvae Return to Their Natal Reef. Curr. Biol. 2016, 26, R1266–R1267. [Google Scholar] [CrossRef]
- Nishimura, T. The Parietal Eye of Lizards (Pogona vitticeps) Needs Light at a Wavelength Lower than 580 Nm to Activate Light-Dependent Magnetoreception. Animals 2020, 10, 489. [Google Scholar] [CrossRef]
- Smith, J.L.B. The Second Coelacanth. Nature 1953, 171, 99–101. [Google Scholar] [CrossRef]
- Yokoyama, S.; Zhang, H.; Radlwimmer, F.B.; Blow, N.S. Adaptive Evolution of Color Vision of the Comoran Coelacanth (Latimeria chalumnae). Proc. Natl. Acad. Sci. USA 1999, 96, 6279–6284. [Google Scholar] [CrossRef] [Green Version]
- Pinzon-Rodriguez, A.; Bensch, S.; Muheim, R. Expression Patterns of Cryptochrome Genes in Avian Retina Suggest Involvement of Cry4 in Light-Dependent Magnetoreception. J. R. Soc. Interface 2018, 15, 20180058. [Google Scholar] [CrossRef] [PubMed]
- Günther, A.; Einwich, A.; Sjulstok, E.; Feederle, R.; Bolte, P.; Koch, K.-W.; Solov’yov, I.A.; Mouritsen, H. Double-Cone Localization and Seasonal Expression Pattern Suggest a Role in Magnetoreception for European Robin Cryptochrome 4. Curr. Biol. 2018, 28, 211–223.e4. [Google Scholar] [CrossRef] [PubMed]
- Einwich, A.; Dedek, K.; Seth, P.K.; Laubinger, S.; Mouritsen, H. A Novel Isoform of Cryptochrome 4 (Cry4b) Is Expressed in the Retina of a Night-Migratory Songbird. Sci. Rep. 2020, 10, 15794. [Google Scholar] [CrossRef] [PubMed]
- Maor, R.; Dayan, T.; Ferguson-Gow, H.; Jones, K.E. Temporal Niche Expansion in Mammals from a Nocturnal Ancestor after Dinosaur Extinction. Nat. Ecol. Evol. 2017, 1, 1889–1895. [Google Scholar] [CrossRef]
- Walls, G.L. The Vertebrate Eye and Its Adaptive Radiation. J. Nerv. Ment. Dis. 1944, 100, 332. [Google Scholar] [CrossRef]
- Gerkema, M.P.; Davies, W.I.L.; Foster, R.G.; Menaker, M.; Hut, R.A. The Nocturnal Bottleneck and the Evolution of Activity Patterns in Mammals. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130508. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.I.; Kamilar, J.M.; Kirk, E.C. Eye Shape and the Nocturnal Bottleneck of Mammals. Proc. R. Soc. B Biol. Sci. 2012, 279, 4962–4968. [Google Scholar] [CrossRef]
- Gillet, L.C.J.; Schärer, O.D. Molecular Mechanisms of Mammalian Global Genome Nucleotide Excision Repair. Chem. Rev. 2006, 106, 253–276. [Google Scholar] [CrossRef]
- De Laat, W.L.; Jaspers, N.G.J.; Hoeijmakers, J.H.J. Molecular Mechanism of Nucleotide Excision Repair. Genes Dev. 1999, 13, 768–785. [Google Scholar] [CrossRef]
- Kang, T.-H.; Reardon, J.T.; Kemp, M.; Sancar, A. Circadian Oscillation of Nucleotide Excision Repair in Mammalian Brain. Proc. Natl. Acad. Sci. USA 2009, 106, 2864–2867. [Google Scholar] [CrossRef] [Green Version]
- Dakup, P.; Gaddameedhi, S. Impact of the Circadian Clock on UV-Induced DNA Damage Response and Photocarcinogenesis. Photochem. Photobiol. 2017, 93, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.-H. Circadian Rhythm of NER and ATR Pathways. Biomolecules 2021, 11, 715. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.S.; Regan, J.D. Photoreactivation and Photoreactivating Enzyme Activity in an Order of Mammals (Marsupialia). Nature 1969, 223, 1066–1067. [Google Scholar] [CrossRef] [PubMed]
- Painter, R.B. DNA Damage and Repair in Eukaryotic Cells. Genetics 1974, 78, 139–148. [Google Scholar] [CrossRef]
- Chaves, I.; Nijman, R.M.; Biernat, M.A.; Bajek, M.I.; Brand, K.; da Silva, A.C.; Saito, S.; Yagita, K.; Eker, A.P.; van der Horst, G.T. The Potorous CPD Photolyase Rescues a Cryptochrome-Deficient Mammalian Circadian Clock. PLoS ONE 2011, 6, e23447. [Google Scholar] [CrossRef]
- Yasui, A.; Eker, A.P.; Yasuhira, S.; Yajima, H.; Kobayashi, T.; Takao, M.; Oikawa, A. A New Class of DNA Photolyases Present in Various Organisms Including Aplacental Mammals. EMBO J. 1994, 13, 6143–6151. [Google Scholar] [CrossRef]
- Ji, Q.; Luo, Z.-X.; Yuan, C.-X.; Wible, J.R.; Zhang, J.-P.; Georgi, J.A. The Earliest Known Eutherian Mammal. Nature 2002, 416, 816–822. [Google Scholar] [CrossRef]
- Alley, N.F.; Hore, S.B.; Frakes, L.A. Glaciations at High-Latitude Southern Australia during the Early Cretaceous. Aust. J. Earth Sci. 2020, 67, 1045–1095. [Google Scholar] [CrossRef]
- Franchini, L.F. Genetic Mechanisms Underlying Cortical Evolution in Mammals. Front. Cell Dev. Biol. 2021, 9, 73. [Google Scholar] [CrossRef]
- Xiong, Y.; Brandley, M.; Xu, S.; Zhou, K.; Yang, G. Seven New Mitochondrial Genomes and a Time-Calibrated Phylogeny of Whales. BMC Evol. Biol. 2009, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.W.; Hejnol, A.; Matus, D.Q.; Pang, K.; Browne, W.E.; Smith, S.A.; Seaver, E.; Rouse, G.W.; Obst, M.; Edgecombe, G.D.; et al. Broad Phylogenomic Sampling Improves Resolution of the Animal Tree of Life. Nature 2008, 452, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Marlétaz, F.; Gilles, A.; Caubit, X.; Perez, Y.; Dossat, C.; Samain, S.; Gyapay, G.; Wincker, P.; Le Parco, Y. Chætognath Transcriptome Reveals Ancestral and Unique Features among Bilaterians. Genome Biol. 2008, 9, R94. [Google Scholar] [CrossRef]
- Vannier, J.; Steiner, M.; Renvoisé, E.; Hu, S.-X.; Casanova, J.-P. Early Cambrian Origin of Modern Food Webs: Evidence from Predator Arrow Worms. Proc. R. Soc. B Biol. Sci. 2007, 274, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Wollesen, T.; Monje, S.V.R.; Oel, A.P.; Arendt, D. Characterization of Eyes, Photoreceptors and Opsins in Developmental Stages of the Chaetognath Spadella cephaloptera. bioRxiv 2019. [Google Scholar] [CrossRef]
- Knight, M.; Ittiprasert, W.; Arican-Goktas, H.D.; Bridger, J.M. Epigenetic Modulation, Stress and Plasticity in Susceptibility of the Snail Host, Biomphalaria Glabrata, to Schistosoma Mansoni Infection. Int. J. Parasitol. 2016, 46, 389–394. [Google Scholar] [CrossRef]
- Pan, S.C. Schistosoma Mansoni: The Ultrastructure of Larval Morphogenesis in Biomphalaria Glabrata and of Associated Host-Parasite Interactions. Jpn. J. Med. Sci. Biol. 1996, 49, 129–149. [Google Scholar] [CrossRef]
- Zantke, J.; Ishikawa-Fujiawara, T.; Arboleda, E.; Lohs, C.; Schipany, K.; Hallay, N.; Straw, A.; Todo, T.; Tessmar-Raible, K. Circadian and Circalunar Clock Interactions in a Marine Annelid. Cell Rep. 2013, 5, 99–113. [Google Scholar] [CrossRef]
- Simakov, O.; Marletaz, F.; Cho, S.-J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.-H.; Larsson, T.; Lv, J.; Arendt, D.; et al. Insights into Bilaterian Evolution from Three Spiralian Genomes. Nature 2013, 493, 526–531. [Google Scholar] [CrossRef]
- Lombardo, P.; Giustini, M.; Miccoli, F.P.; Cicolani, B. Fine-Scale Differences in Diel Activity among Nocturnal Freshwater Planarias (Platyhelminthes: Tricladida). J. Circadian Rhythm. 2011, 9, 2. [Google Scholar] [CrossRef]
- Hasegawa, K.; Saigusa, T.; Tamai, Y. Caenorhabditis Elegans Opens Up New Insights into Circadian Clock Mechanisms. Chronobiol. Int. 2005, 22, 1–19. [Google Scholar] [CrossRef]
- Olmedo, M.; Merrow, M.; Geibel, M. Sleeping Beauty? Developmental Timing, Sleep, and the Circadian Clock in Caenorhabditis Elegans. Adv. Genet. 2017, 97, 43–80. [Google Scholar] [CrossRef] [PubMed]
- Milev, N.B.; Rhee, S.-G.; Reddy, A.B. Cellular Timekeeping: It’s Redox o’Clock. Cold Spring Harb. Perspect. Biol. 2018, 10, a027698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hastings, M.H.; Green, E.W.; Tauber, E.; Sladek, M.; Webster, S.G.; Kyriacou, C.P.; Wilcockson, D.C. Dissociation of Circadian and Circatidal Timekeeping in the Marine Crustacean Eurydice Pulchra. Curr. Biol. 2013, 23, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, J.F.; Hoelters, L.S.; Swain, M.T.; Wilcockson, D.C. Identification and Temporal Expression of Putative Circadian Clock Transcripts in the Amphipod Crustacean Talitrus Saltator. PeerJ 2016, 4, e2555. [Google Scholar] [CrossRef] [PubMed]
- Biscontin, A.; Wallach, T.; Sales, G.; Grudziecki, A.; Janke, L.; Sartori, E.; Bertolucci, C.; Mazzotta, G.; De Pittà, C.; Meyer, B.; et al. Functional Characterization of the Circadian Clock in the Antarctic Krill, Euphausia Superba. Sci. Rep. 2017, 7, 17742. [Google Scholar] [CrossRef]
- Lapadula, W.J.; Mascotti, M.L.; Juri Ayub, M. Whitefly Genomes Contain Ribotoxin Coding Genes Acquired from Plants. Sci. Rep. 2020, 10, 15503. [Google Scholar] [CrossRef]
- Méteignier, L.-V.; Papon, N.; Courdavault, V. Plant to Insect Horizontal Gene Transfer: Empowering Whiteflies. Trends Genet. 2021, 37, 688–690. [Google Scholar] [CrossRef]
- Gilbert, C.; Maumus, F. Multiple Horizontal Acquisitions of Plant Genes in the Whitefly Bemisia Tabaci. bioRxiv 2022. [Google Scholar] [CrossRef]
- Werckenthin, A.; Derst, C.; Stengl, M. Sequence and Expression of per, Tim1, and Cry2 Genes in the Madeira Cockroach Rhyparobia Maderae. J. Biol. Rhythm. 2012, 27, 453–466. [Google Scholar] [CrossRef]
- Ueda, H.; Tamaki, S.; Miki, T.; Uryu, O.; Kamae, Y.; Nose, M.; Shinohara, T.; Tomioka, K. Cryptochrome Genes Mediate Photoperiodic Responses in the Cricket Modicogryllus siamensis. Physiol. Entomol. 2018, 43, 285–294. [Google Scholar] [CrossRef]
- Cai, S.; Huang, Y.; Chen, F.; Zhang, X.; Sessa, E.; Zhao, C.; Marchant, D.B.; Xue, D.; Chen, G.; Dai, F.; et al. Evolution of Rapid Blue-Light Response Linked to Explosive Diversification of Ferns in Angiosperm Forests. New Phytol. 2021, 230, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Kanno, S.; Smit, B.; van der Horst, G.T.; Takao, M.; Yasui, A. Characterization of Photolyase/Blue-Light Receptor Homologs in Mouse and Human Cells. Nucleic Acids Res. 1998, 26, 5086–5092. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Sancar, A. Circadian Regulation of Cryptochrome Genes in the Mouse. Mol. Brain Res. 1999, 71, 238–243. [Google Scholar] [CrossRef]
- Kondrashov, F.A.; Koonin, E.V.; Morgunov, I.G.; Finogenova, T.V.; Kondrashova, M.N. Evolution of Glyoxylate Cycle Enzymes in Metazoa: Evidence of Multiple Horizontal Transfer Events and Pseudogene Formation. Biol. Direct 2006, 1, 31. [Google Scholar] [CrossRef]
- Starcevic, A.; Akthar, S.; Dunlap, W.C.; Shick, J.M.; Hranueli, D.; Cullum, J.; Long, P.F. Enzymes of the Shikimic Acid Pathway Encoded in the Genome of a Basal Metazoan, Nematostella vectensis, Have Microbial Origins. Proc. Natl. Acad. Sci. USA 2008, 105, 2533–2537. [Google Scholar] [CrossRef]
- Xia, J.; Guo, Z.; Yang, Z.; Han, H.; Wang, S.; Xu, H.; Yang, X.; Yang, F.; Wu, Q.; Xie, W.; et al. Whitefly Hijacks a Plant Detoxification Gene That Neutralizes Plant Toxins. Cell 2021, 184, 1693–1705.e17. [Google Scholar] [CrossRef]
- Biller, S.J.; Schubotz, F.; Roggensack, S.E.; Thompson, A.W.; Summons, R.E.; Chisholm, S.W. Bacterial Vesicles in Marine Ecosystems. Science 2014, 343, 183–186. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Frazier, K.; Chang, E.B. Intersection of the Gut Microbiome and Circadian Rhythms in Metabolism. Trends Endocrinol. Metab. 2020, 31, 25–36. [Google Scholar] [CrossRef]
- Sen, P.; Molinero-Perez, A.; O’Riordan, K.J.; McCafferty, C.P.; O’Halloran, K.D.; Cryan, J.F. Microbiota and Sleep: Awakening the Gut Feeling. Trends Mol. Med. 2021, 27, 935–945. [Google Scholar] [CrossRef]
- Heath-Heckman, E.A.C.; Foster, J.; Apicella, M.A.; Goldman, W.E.; McFall-Ngai, M. Environmental Cues and Symbiont Microbe-Associated Molecular Patterns Function in Concert to Drive the Daily Remodelling of the Crypt-Cell Brush Border of the Euprymna Scolopes Light Organ. Cell. Microbiol. 2016, 18, 1642–1652. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510.e12. [Google Scholar] [CrossRef] [PubMed]
- Jabbur, M.L.; Johnson, C.H. Spectres of Clock Evolution: Past, Present, and Yet to Come. Front. Physiol. 2022, 12, 2526. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.E.A.; Lakin, J.; Troth, I.; Wallace-Johnson, S.M. UV-B Radiation Was the Devonian-Carboniferous Boundary Terrestrial Extinction Kill Mechanism. Sci. Adv. 2020, 6, eaba0768. [Google Scholar] [CrossRef]
- McElwain, J.C.; Punyasena, S.W. Mass Extinction Events and the Plant Fossil Record. Trends Ecol. Evol. 2007, 22, 548–557. [Google Scholar] [CrossRef]
- Visscher, H.; Looy, C.V.; Collinson, M.E.; Brinkhuis, H.; van Konijnenburg-van Cittert, J.H.A.; Kürschner, W.M.; Sephton, M.A. Environmental Mutagenesis during the End-Permian Ecological Crisis. Proc. Natl. Acad. Sci. USA 2004, 101, 12952–12956. [Google Scholar] [CrossRef]
- Vanderstraeten, J.; Gailly, P.; Malkemper, E.P. Light Entrainment of Retinal Biorhythms: Cryptochrome 2 as Candidate Photoreceptor in Mammals. Cell. Mol. Life Sci. 2020, 77, 875–884. [Google Scholar] [CrossRef]
- Konopka, R.J.; Benzer, S. Clock Mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA Damage, Repair, and Mutagenesis. Environ. Mol. Mutagenesis 2017, 58, 235–263. [Google Scholar] [CrossRef] [Green Version]
- Merrow, M.; Roenneberg, T. Circadian Clocks: Running on Redox. Cell 2001, 106, 141–143. [Google Scholar] [CrossRef]
- Nagy, A.D.; Reddy, A.B. Redox Clocks: Time to Rethink Redox Interventions. Free Radic. Biol. Med. 2018, 119, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Pokorny, R.; Klar, T.; Hennecke, U.; Carell, T.; Batschauer, A.; Essen, L.O. Recognition and Repair of UV Lesions in Loop Structures of Duplex DNA by DASH-Type Cryptochrome. Proc. Natl. Acad. Sci. USA 2008, 105, 21023–21027. [Google Scholar] [CrossRef]
- Cao, S.; He, S.; Lv, H.; Zhang, J.; Aslam, M.; Cheng, H.; Hu, A.; Cao, G.; Zhang, X.; Yu, Y.; et al. Genome-Wide Analysis of the Cryptochrome Gene Family in Plants. Trop. Plant Biol. 2020, 13, 117–126. [Google Scholar] [CrossRef]
- Partch, C.L.; Clarkson, M.W.; Ozgur, S.; Lee, A.L.; Sancar, A. Role of Structural Plasticity in Signal Transduction by the Cryptochrome Blue-Light Photoreceptor. Biochemistry 2005, 44, 3795–3805. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Shalitin, D. Cryptochrome Structure and Signal Transduction. Annu. Rev. Plant Biol. 2003, 54, 469–496. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K.; DiTacchio, L.; Arvai, A.S.; Yamamoto, J.; Kim, S.T.; Todo, T.; Tainer, J.A.; Iwai, S.; Panda, S.; Getzoff, E.D. Functional Motifs in the (6-4) Photolyase Crystal Structure Make a Comparative Framework for DNA Repair Photolyases and Clock Cryptochromes. Proc. Natl. Acad. Sci. USA 2009, 106, 6962–6967. [Google Scholar] [CrossRef]
- Xing, W.; Busino, L.; Hinds, T.R.; Marionni, S.T.; Saifee, N.H.; Bush, M.F.; Pagano, M.; Zheng, N. SCF(FBXL3) Ubiquitin Ligase Targets Cryptochromes at Their Cofactor Pocket. Nature 2013, 496, 64–68. [Google Scholar] [CrossRef]
- Rosensweig, C.; Reynolds, K.A.; Gao, P.; Laothamatas, I.; Shan, Y.; Ranganathan, R.; Takahashi, J.S.; Green, C.B. An Evolutionary Hotspot Defines Functional Differences between CRYPTOCHROMES. Nat. Commun. 2018, 9, 1138. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Vaidya, A.T.; Top, D.; Widom, J.; Young, M.W.; Crane, B.R. Structure of Full-Length Drosophila Cryptochrome. Nature 2011, 480, 396–399. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, L.; Shu, S.; Sancar, A.; Zhong, D. Bifurcating Electron-Transfer Pathways in DNA Photolyases Determine the Repair Quantum Yield. Science 2016, 354, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, K.; Arvai, A.S.; Yamamoto, J.; Hitomi, C.; Teranishi, M.; Hirouchi, T.; Yamamoto, K.; Iwai, S.; Tainer, J.A.; Hidema, J.; et al. Eukaryotic Class II Cyclobutane Pyrimidine Dimer Photolyase Structure Reveals Basis for Improved Ultraviolet Tolerance in Plants. J. Biol. Chem. 2012, 287, 12060–12069. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Teranishi, M.; Izumi, M.; Takahashi, M.; Takahashi, F.; Hidema, J. Transport of Rice Cyclobutane Pyrimidine Dimer Photolyase into Mitochondria Relies on a Targeting Sequence Located in Its C-terminal Internal Region. Plant J. 2014, 79, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Liu, X.; Zhou, X.; Song, Q.; Fu, M.; Xu, F.; Wang, X. Functional Analysis of a Novel Cryptochrome Gene (GbCRY1) from Ginkgo biloba. Plant Signal. Behav. 2021, 16, 1850627. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRY/PL Type | Species | Name | Accession No. |
| DCRY | Drosophila melanogaster | CRY | NP_732407.1 |
| 6-4 PL | Drosophila melanogaster | 6-4 PHR | BAA12067.1 |
| MCRY | Mus musculus | CRY1 | NP_031797.1 |
| DASH | Synechocystis sp. PCC 6803 | DASH | AGF51454.1 |
| CPDI | Escherichia coli | PHOTOLYASE | WP_001583322.1 |
| CPDII | Drosophila melanogaster | PHR | NP_523653.2 |
| CPDIII | Agrobacterium fabrum | PHOTOLYASE | WP_174020122.1 |
| PCRY | Arabidopsis thaliana | CRY2 | AAL16379.1 |
| PCRY-like | Phaeodactylum tricornutum | CryP | XP_002179379.1 |
| PPL | Arabidopsis thaliana | PHR2 | NP_182281.1 |
| CRY4 | Danio rerio | CRY4 | XP_005168334.1 |
| MCRY | CRY4 | 6-4-PL | DCRY | ACRY | PCRY like | CPDI | CPDII | CPDIII | PCRY | DASH | PPL | other | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Eukaryota | Metazoa (497) | 86% | 25% | 47% | 18% | 2% | 18% | 2% | 76% | 1% | - | 50% | - | 2% |
| Fungi (34) | - | - | 65% | - | - | - | 85% | 12% | - | - | 53% | - | - | |
| Viridiplantae (28) | - | - | 86% | - | - | 11% | 4% | 86% | 4% | 75% | 82% | 61% | - | |
| unicellular Eukaryota (51) | - | - | 63% | - | - | 25% | 14% | 73% | 2% | - | 65% | 6% | 22% | |
| Bacteria (20) | - | - | 15% | - | - | - | 65% | 25% | 15% | - | 30% | - | - | |
| Archea (9) | - | - | 33% | - | - | - | 22% | 44% | 11% | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deppisch, P.; Helfrich-Förster, C.; Senthilan, P.R. The Gain and Loss of Cryptochrome/Photolyase Family Members during Evolution. Genes 2022, 13, 1613. https://doi.org/10.3390/genes13091613
Deppisch P, Helfrich-Förster C, Senthilan PR. The Gain and Loss of Cryptochrome/Photolyase Family Members during Evolution. Genes. 2022; 13(9):1613. https://doi.org/10.3390/genes13091613
Chicago/Turabian StyleDeppisch, Peter, Charlotte Helfrich-Förster, and Pingkalai R. Senthilan. 2022. "The Gain and Loss of Cryptochrome/Photolyase Family Members during Evolution" Genes 13, no. 9: 1613. https://doi.org/10.3390/genes13091613
APA StyleDeppisch, P., Helfrich-Förster, C., & Senthilan, P. R. (2022). The Gain and Loss of Cryptochrome/Photolyase Family Members during Evolution. Genes, 13(9), 1613. https://doi.org/10.3390/genes13091613