
Characterization of the Genetic Variability within Ziziphus nummularia Genotypes by Phenotypic Traits and SSR Markers with Special Reference to Geographic Distribution


Abstract
:1. Introduction
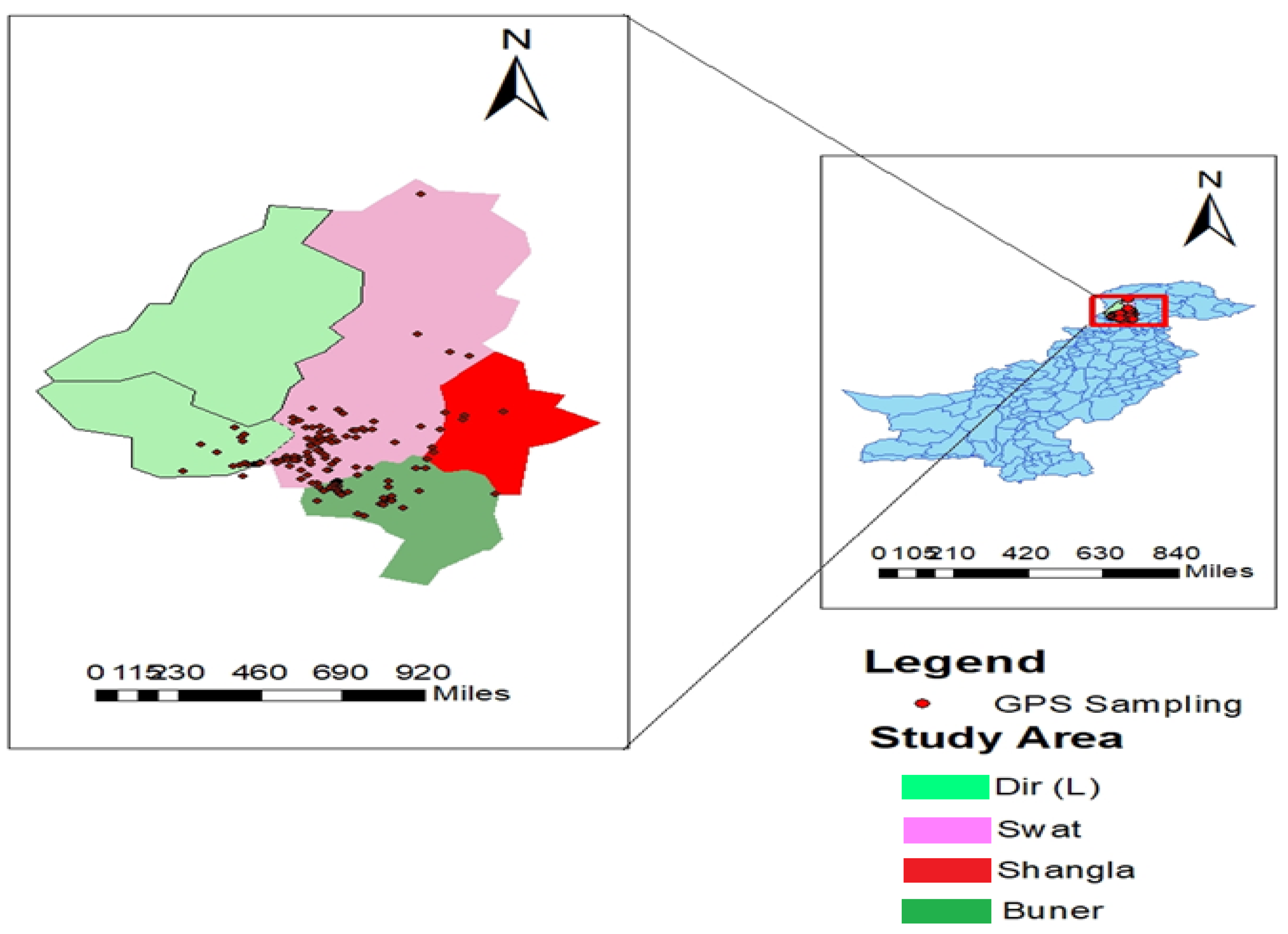
2. Materials and Methods
2.1. Phenotypic Evaluation
2.2. Genomic DNA Extraction
2.3. PCR Reaction for SSR Markers
2.4. Data Analysis
3. Results
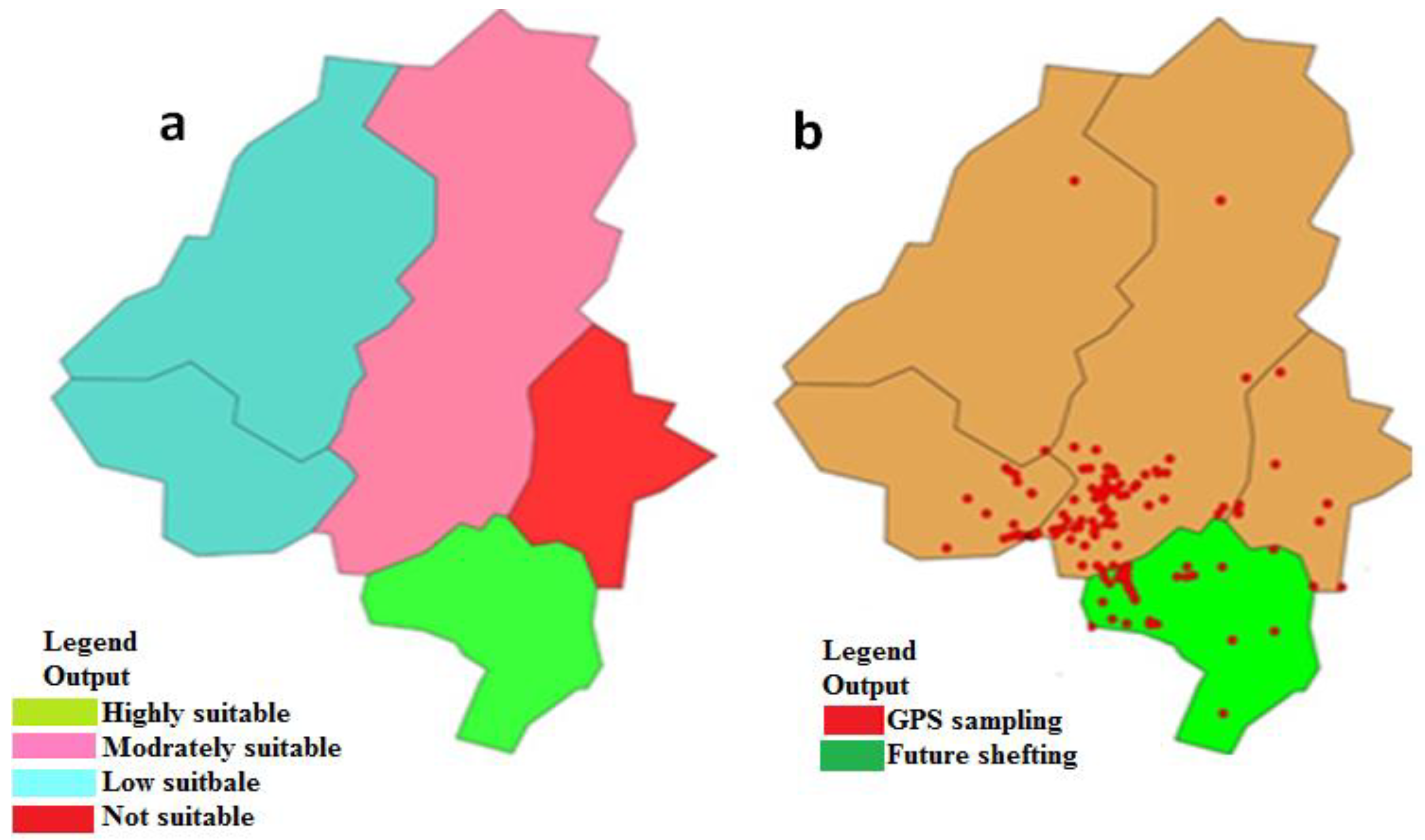
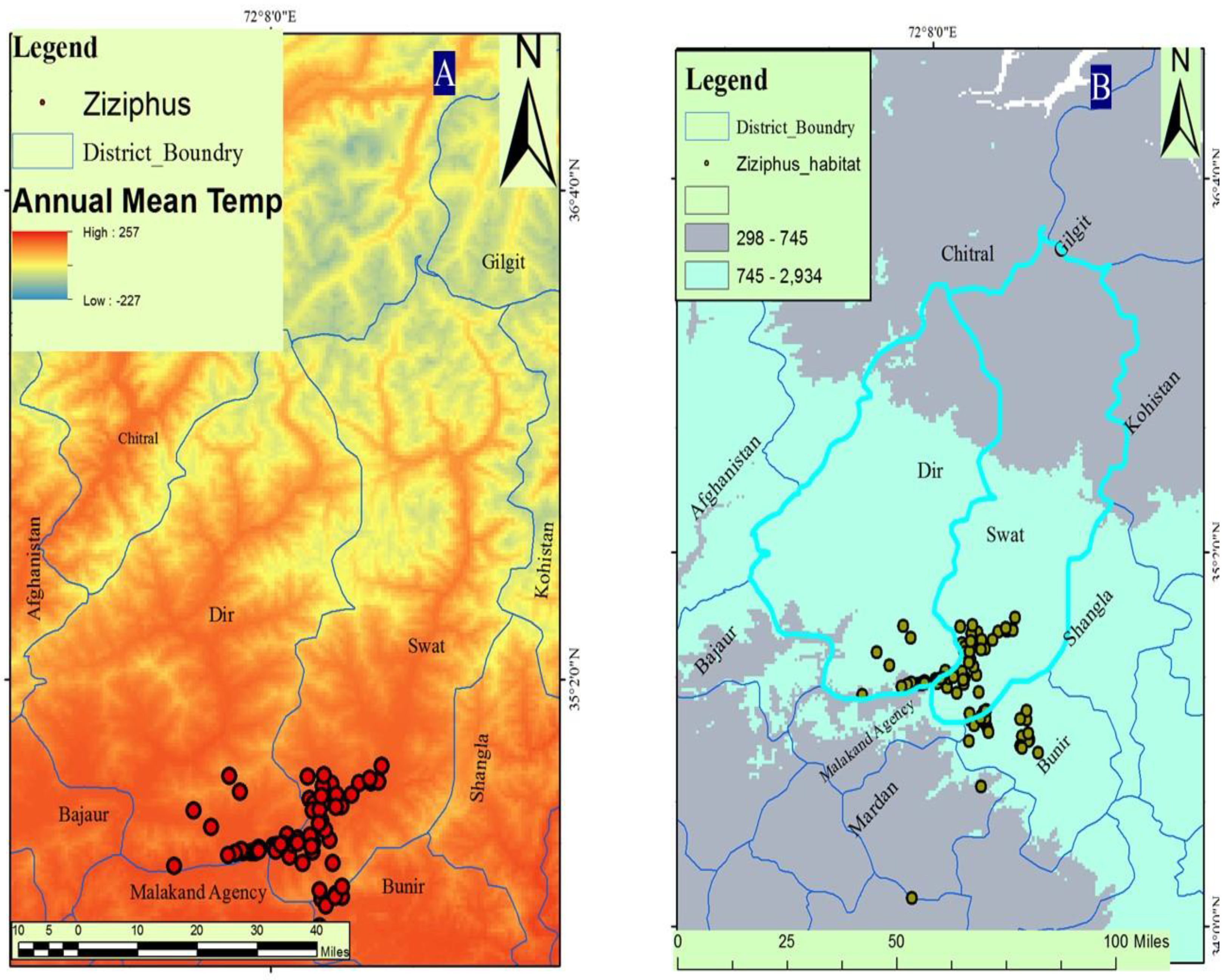
3.1. Bioclimatic Layers
3.2. Phenotypic Diversity and Correlation among the Traits
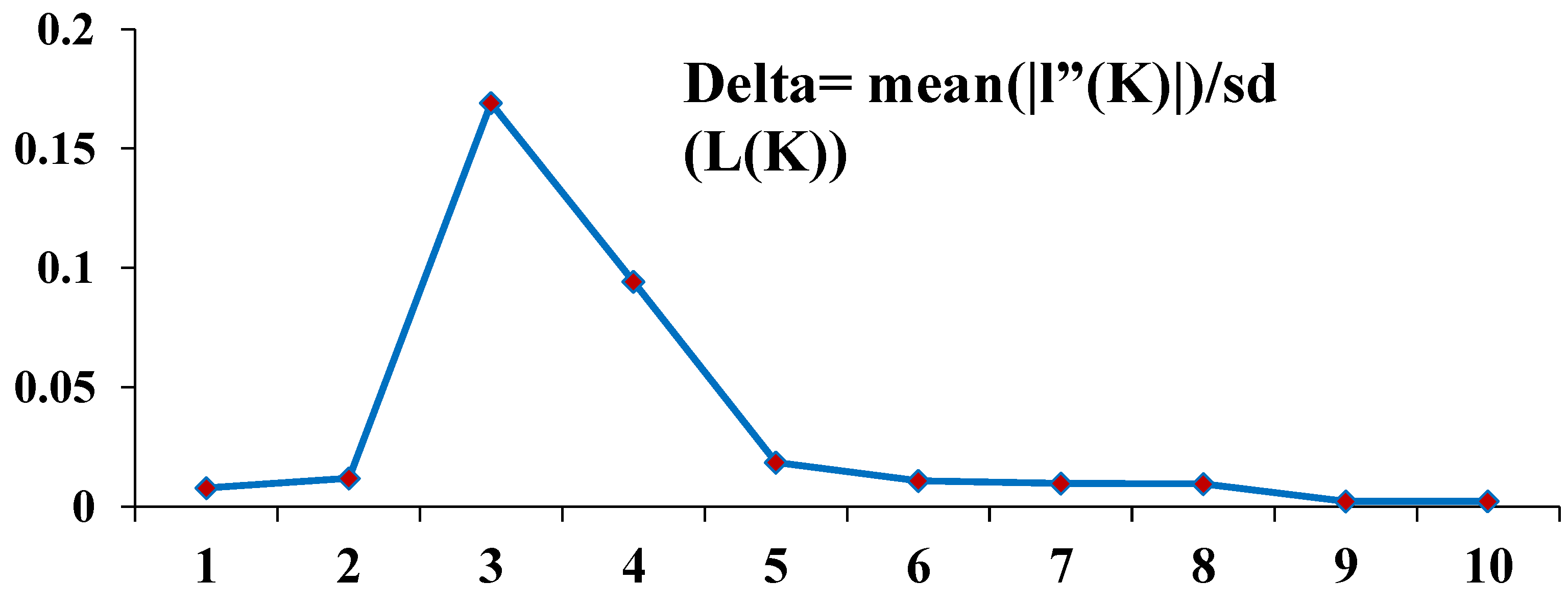
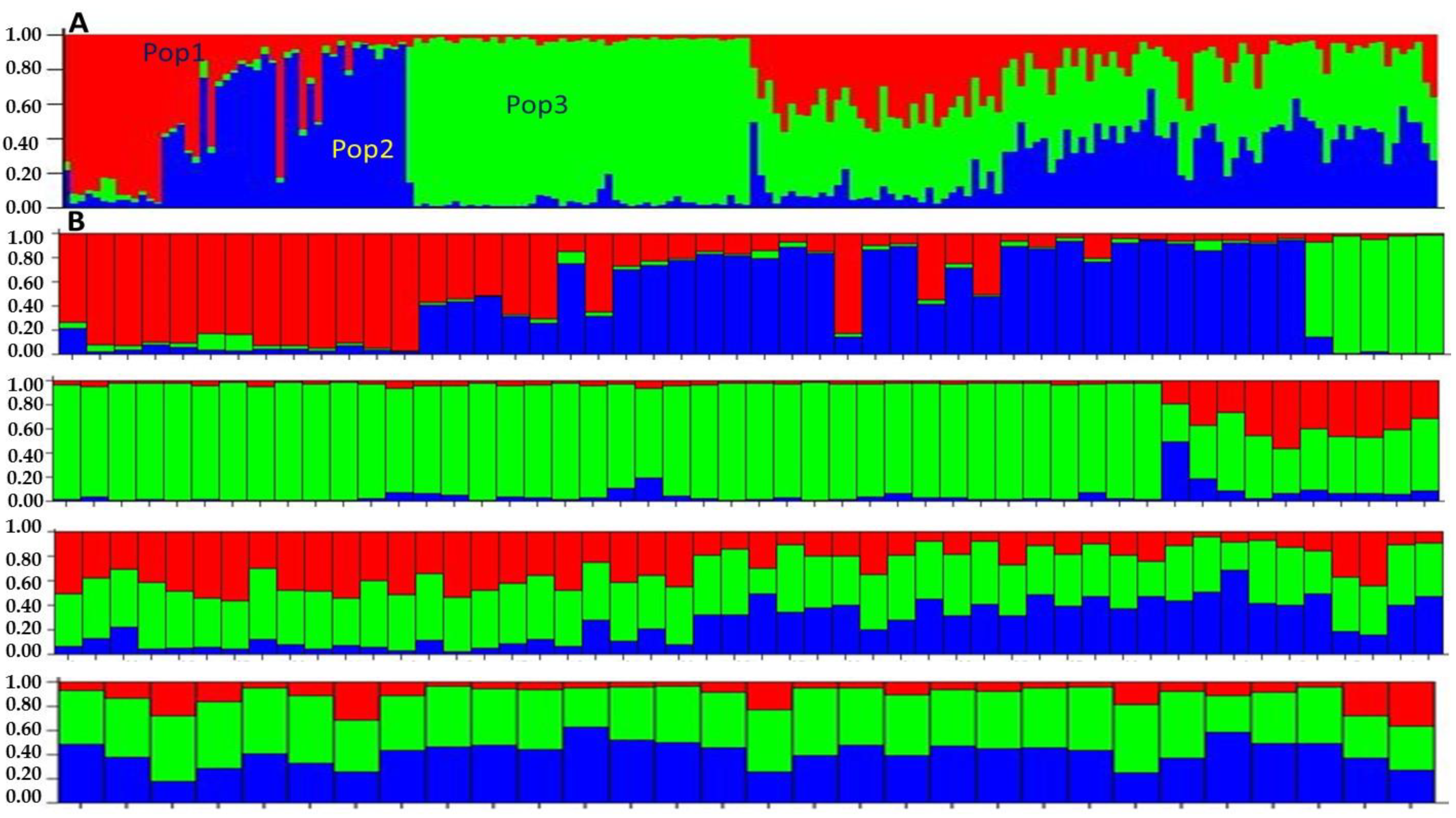
3.3. Genetic Diversity and Population Structure
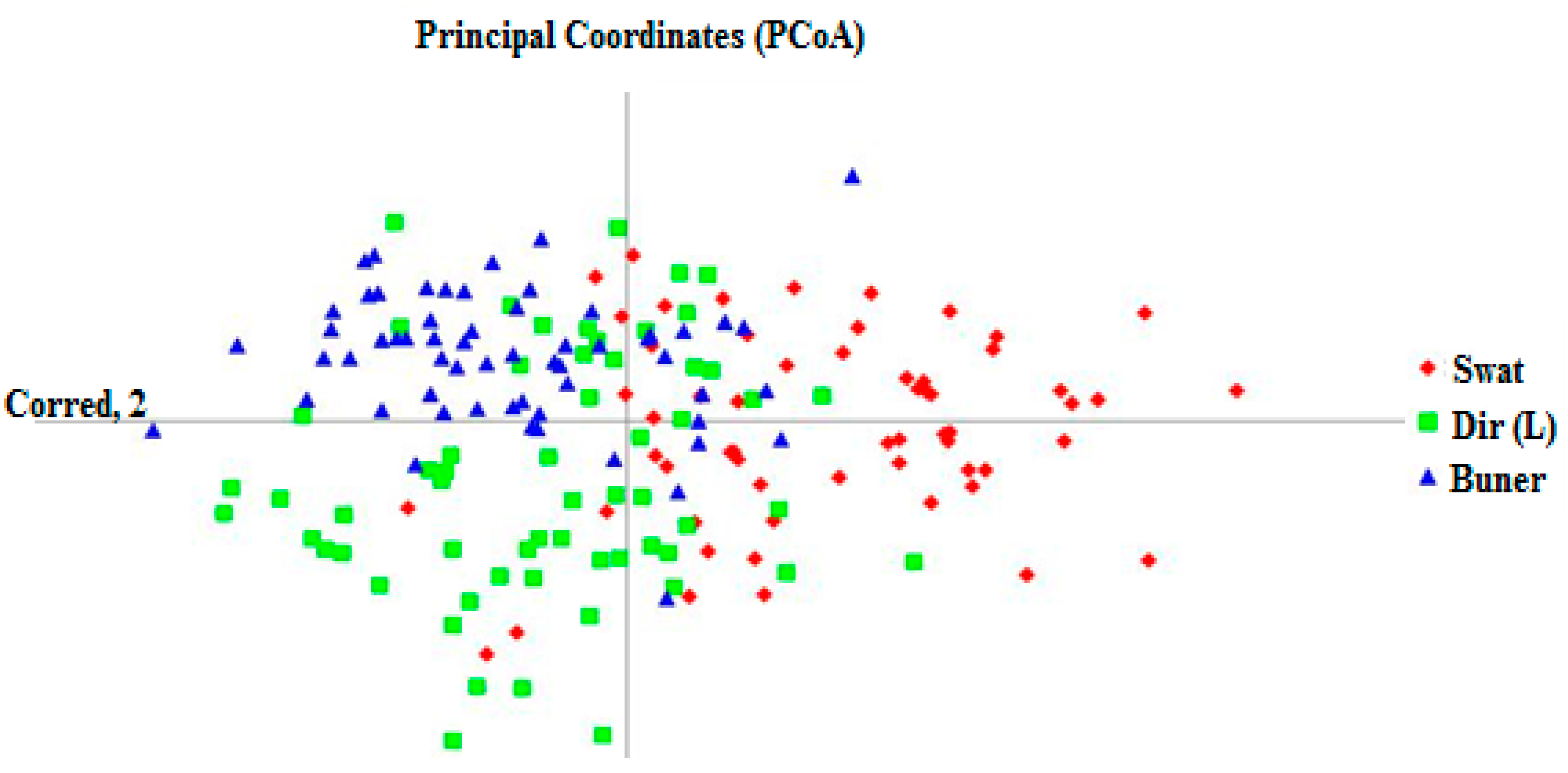
3.4. Neighbor-Joining (NJ) and Principal Coordinate Analysis (PCoA)
3.5. Analysis of Molecular Variance (AMOVA)
4. Discussion
4.1. Distribution and Climate Change
4.2. Morphological Variation in Z. nummularia Genotypes
4.3. Genetic Differentiation and Population Structure Analysis
4.4. Analysis of the Evolutionary Relationships of the Z. nummularia Population
4.5. Conservation Implication
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frankham, R.; Ballou, J.D.; Ralls, K.; Eldridge, M.; Dudash, M.R.; Fenster, C.B.; Lacy, R.C.; Sunnucks, P. Genetic Management of Fragmented Animal and Plant Populations; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- Vít, P.; Douda, J.; Krak, K.; Havrdová, A.; Mandák, B. Two new polyploid species closely related to Alnus glutinosa in Europe and North Africa–an analysis based on morphometry, karyology, flow cytometry and microsatellites. Taxon 2017, 66, 567–583. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Q.; Lu, C.; Liu, Q.; Pan, J.; Zhang, J.; Dong, S. Genetic diversity of Prunus armeniaca L. var. ansu Maxim. germplasm revealed by simple sequence repeat (SSR) markers. PloS ONE 2022, 17, e0269424. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Wuyun, T.; Wang, L.; Zhao, H. Genetic diversity and population structure of wild apricot in Xinjiang revealed by SSR markers. Acta Bot. Boreali-Occident. Sin. 2016, 36, 1757–1763. [Google Scholar]
- Zhang, D.Q.; Zhou, N. Genetic diversity and population structure of the endangered conifer Taxus wallichiana var. mairei (Taxaceae) revealed by Simple Sequence Repeat (SSR) markers. Biochem. Syst. Ecol. 2013, 49, 107–114. [Google Scholar] [CrossRef]
- Aksehirli-Pakyurek, M.; Koubouris, G.C.; Petrakis, P.V.; Hepaksoy, S.; Metzidakis, I.T.; Yalcinkaya, E. Cultivated and Wild Olives in Crete, Greece-Genetic Diversity and Relationships with Major Turkish Cultivars Revealed by SSR Markers. Plant Mol. Biol. Report. 2017, 35, 575–585. [Google Scholar] [CrossRef]
- Liu, Z.L.; Wan, S.L.; Yan, C.P.; Hu, Z.D.; He, X.H.; Zeng, P.Z. Genetic Diversity of Jinsha Pomelo and Its Closely-related Germplasms Assessed by SSR Molecular Markers. Agric. Biotechnol. 2017, 6, 15–22. [Google Scholar]
- Belamkar, V.; Selvaraj, M.G.; Ayers, J.L.; Payton, P.R.; Puppala, N.; Burow, M.D. A first insight into population structure and linkage disequilibrium in the US peanut minicore collection. Genetica 2011, 139, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wang, J.; Wang, L.; Liu, P.; Zhao, J.; Zhao, Z.; Yao, S.; Stănică, F.; Liu, Z.; Wang, L.; et al. The historical and current research progress on jujube–a superfruit for the future. Hortic. Res. 2020, 1, 7. [Google Scholar] [CrossRef]
- Rashwan, A.K.; Karim, N.; Shishir, M.R.; Bao, T.; Lu, Y.; Chen, W. Jujube fruit: A potential nutritious fruit for the development of functional food products. J. Funct. Foods 2020, 75, 104205. [Google Scholar] [CrossRef]
- Muhammad, N.; Zhao, Z.H.; Yang, M.; Liu, M.J. Current status of Ziziphus species: Their distribution, cultivation, and production in Pakistan. SHS Acta Horticulturae 1350 V International Jujube Symposium. Acta Hortic. 2022, 1350, 23–34. [Google Scholar] [CrossRef]
- Ren, X.P.; Jiang, H.F.; Liao, B.S.; Zhang, X.J.; Lei, Y.; Huang, G.Q.; Yan, L.Y.; Chen, Y.N. Distributing and genetic diversity of high oleic acid germplasm in Peanut (Arachia Hypogaea L.) core collection of China. J. Plant Genet. Resour. 2011, 12, 513–518. [Google Scholar]
- Nisar Uddin, N.M.; Nisar, M.; Muhammad, J.; Ali, N. Genetic diversity in the natural population of Ziziphus jujuba Miil. Int. J. Bot. Stud. 2019, 4, 29–35. [Google Scholar]
- Allen, J.L.; Lendemer, J.C. Climate change impacts on endemic, high-elevation lichens in a biodiversity hotspot. Biodivers. Conserv. 2016, 25, 555–568. [Google Scholar] [CrossRef]
- Acevedo, M.A.; Beaudrot, L.; Meléndez-Ackerman, E.J.; Tremblay, R.L. Local extinction risk under climate change in a neotropical asymmetrically dispersed epiphyte. J. Ecol. 2020, 108, 1553–1564. [Google Scholar] [CrossRef]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Muhammad, N.; Luo, Z.; Yang, M.; Liu, Z.; Liu, M. The nutritional, medicinal, and drought-resistance properties of ziziphus mill. Make it an important candidate for alleviating food insecurity in arid Regions—A case of Pakistan. Horticulturae 2022, 8, 867. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, J.G.; Kang, B.C.; Song, K. Assessment of the genetic diversity of the breeding lines and a genome wide association study of three horticultural traits using worldwide cucumber (Cucumis spp.) germplasm collection. Agronomy 2020, 10, 1736. [Google Scholar] [CrossRef]
- Muhammad, N.; Khan, M.K.; Ali, N.; Fazal, S. Narrow genetic diversity is a major threat to Abutilon indicum L. in Swat valley, Pakistan. Biosci Res. Biosci. 2020, 17, 4. [Google Scholar]
- Uddin, N.; Ali, N.; Nisar, M.; Liu, M.; Liu, Z.; Muhammad, N.; Rahman, I.U. SSR-based population structure, molecular diversity and identity cards of Ziziphus species from Pakistan and China. Genet. Resour. Crop Evol. 2021, 68, 2391–2409. [Google Scholar] [CrossRef]
- Uddin, N.; Muhammad, N.; Ali, N.; Nisar, M.; Liu, M. Genomic constitution and relationships of Ziziphus species collected from Malakand Division, Pakistan. Physiol. Mol. Biol. Plants 2022, 11, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Uddin, N.; Ali, N.; Muhammad, N.; Zehra, S.A.; Nisar, N.; Khan, M.K. Morphological attributes and total seed protein revealed diversity in Ziziphus nummularia Burm. f. populations in Malakand division, Pakistan. Pak J. Bot 2021, 53, 1727–1735. [Google Scholar] [CrossRef]
- Xiao, J.; Zhao, J.; Liu, M.; Liu, P.; Dai, L.; Zhao, Z. Genome-wide characterization of simple sequence repeat (SSR) loci in Chinese jujube and jujube SSR primer transferability. PLoS ONE 2015, 10, e0127812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wu, Y.T.; Guo, W.Z.; Zhang, T.Z. Fast screening of microsatellite markers in cotton with PAGE/silver staining. Acta Gossypii Sin. 2000, 12, 267–269. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.O.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Edwards, D.; Zander, M.; Dalton-Morgan, J.; Batley, J. New technologies for ultrahigh-throughput genotyping in plant taxonomy. In Molecular Plant Taxonomy; Humana Press: Totowa, NJ, USA, 2014; pp. 151–175. [Google Scholar]
- Pachauri, V.; Taneja, N.; Vikram, P.; Singh, N.K.; Singh, S. Molecular and morphological characterization of Indian farmers rice varieties (Oryza sativa L.). Aust. J. Crop Sci. 2013, 7, 923. [Google Scholar]
- Sarikamis, G.; Yanmaz, R.U.; Ermis, S.; Bakir, M.E.; Yuksel, C. Genetic characterization of pea (Pisum sativum) germplasm from Turkey using morphological and SSR markers. Genet. Mol. Res. 2010, 9, 591–600. [Google Scholar] [CrossRef]
- Gixhari, B.; Pavelkova, M.; Ismaili, H.; Vrapi, H.; Jaupi, A.; Smýkal, P. Genetic diversity of Albanian pea (Pisum sativum L.) landraces assessed by morphological traits and molecular markers. Czech J. Genet. Plant Breed. 2014, 50, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, G.; Kudesia, R.; Srivastava, M.K. Evaluation of genetic diversity in pea (Pisum sativum L) using RAPD analysis. Genet. Eng. Biotechnol. J. 2010, 16, 1–5. [Google Scholar]
- Zhou, P.-Y.; Hui, L.-X.; Huang, S.-J.; Ni, Z.-X.; Yu, F.-X.; Xu, L.-A. Study on the Genetic Structure Based on Geographic Populations of the Endangered Tree Species: Liriodendron chinense. Forests 2021, 12, 917. [Google Scholar] [CrossRef]
- Ponnaiah, M.; Shiferaw, E.; Pe, M.E.; Porceddu, E. Development and application of EST-SSRs for diversity analysis in Ethiopian grass pea. Plant Genet. Resour. 2011, 9, 276–280. [Google Scholar] [CrossRef]
- Cieslarová, J.; Hýbl, M.; Griga, M.; Smýkal, P. Molecular analysis of temporal genetic structuring in pea (Pisum sativum L.) cultivars bred in the Czech Republic and in former Czechoslovakia since the mid-20th century. Czech J. Genet. Plant Breed. 2012, 48, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Jing, R.; Ambrose, M.A.; Knox, M.R.; Smykal, P.; Hybl, M.; Ramos, A.; Caminero, C.; Burstin, J.; Duc, G.; Van Soest, L.J.; et al. Genetic diversity in European Pisum germplasm collections. Theor. Appl. Genet. 2012, 125, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagenblad, J.; Zie, J.; Leino, M.W. Exploring the population genetics of genebank and historical landrace varieties. Genet. Resour. Crop Evol. 2012, 59, 1185–1199. [Google Scholar] [CrossRef]
- Martin-Sanz, A.; Caminero, C.; Jing, R.; Flavell, A.J.; de la Vega, M.P. Genetic diversity among Spanish pea (Pisum sativum L.) landraces, pea cultivars and the World Pisum sp. core collection assessed by retrotransposon-based insertion polymorphisms (RBIPs). Span. J. Agric. Res. 2011, 9, 166–178. [Google Scholar] [CrossRef]
- Kumari, P.; Basal, N.; Singh, A.K.; Rai, V.P.; Srivastava, C.P.; Singh, P.K. Genetic diversity studies in pea (Pisum sativum L.) using simple sequence repeat markers. Genet. Mol. Res. 2013, 12, 3540–3550. [Google Scholar] [CrossRef]
- Kuleung, C.; Baenziger, P.S.; Kachman, S.D.; Dweikat, I. Evaluating the genetic diversity of triticale with wheat and rye SSR markers. Crop Sci. 2006, 46, 1692–1700. [Google Scholar] [CrossRef]
- Wróblewska, A. Genetic diversity and spatial genetic structure of Chamaedaphne calyculata (Ericaceae) at the western periphery in relation to its main continuous range in Eurasia. Folia Geobot. 2014, 49, 193–208. [Google Scholar] [CrossRef]
- Billotte, N.; Jourjon, M.F.; Marseillac, N.; Berger, A.; Flori, A.; Asmady, H.; Adon, B.; Singh, R.; Nouy, B.; Potier, F.; et al. QTL detection by multi-parent linkage mapping in oil palm (Elaeis guineensis Jacq.). Theor. Appl. Genet. 2010, 120, 1673–1687. [Google Scholar] [CrossRef] [Green Version]
- Szczecińska, M.; Sramko, G.; Wołosz, K.; Sawicki, J. Genetic diversity and population structure of the rare and endangered plant species Pulsatilla patens (L.) Mill in East Central Europe. PloS ONE 2016, 11, e0151730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.; Ramos-Cabrer, A.M.; Carnide, V.; Pinto-Carnide, O.; Assunção, A.; Marreiros, A.; Rodrigues, R.; Pereira-Lorenzo, S.; Castro, I. Genetic pool structure of local apple cultivars from Portugal assessed by microsatellites. Tree Genet. Genomes 2016, 12, 1–5. [Google Scholar] [CrossRef]
- Potts, S.M.; Han, Y.; Khan, M.A.; Kushad, M.M.; Rayburn, A.L.; Korban, S.S. Genetic diversity and characterization of a core collection of Malus germplasm using simple sequence repeats (SSRs). Plant Mol. Biol. Rep. 2012, 30, 827–837. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Repeat Motif | Forward | Reverse | BP | Temp °C |
|---|---|---|---|---|---|
| ZSSR-21 | AT(21) | ACTCATTCCGTAAATTACACAGCC | TGAATTTCTAAATTTCACCAAAAACAA | 228 | 57 °C |
| ZSSR-93 | TTG(13) | GGAAGGACTTTGTCAGCATGGTAG | AACAGCATATTTGGATCCATTTCG | 155 | 57 °C |
| ZSSR-95 | TTG(11) | CGGTGAGAGACATTTTGTTGGATT | TTCCTTACTTTCCCACCTTGTTCA | 152 | 57 °C |
| ZSSR-97 | TTG(9) | GTCCAAAGGCCCAACTTCTTTAGT | AGGGGACTACTCCTCTGCTGAGAT | 155 | 57 °C |
| ZSSR-110 | TTA(15) | ACCTTTCGCTTAGTTCTTGCTGAA | GGACTTTTGTCCGGACCTTAAAA | 225 | 57 °C |
| ZSSR-129 | TGG(9) | TGCTAATGAAAGGAACTCTGGGTC | TGATGGGTATGAAGAAGCATCAGA | 158 | 57 °C |
| ZSSR-152 | TAT(18) | TCAGAATTTCTCACTTTGGCTAACAA | TGCACCGATCCTCTCCTCTC | 160 | 56 °C |
| ZSSR-175 | GTT(9) | CAAAGGAAAATCTCTATGGTTGTCG | CGCTACCATGTTAAAATTTGTCCC | 155 | 57 °C |
| ZSSR-181 | CTT(17) | TTTTGTCTCTCCCTCTTTTCTCCA | GGCCTTTTCATGAAGCTTTTGTTA | 137 | 57 °C |
| ZSSR-183 | CTT(14) | ATAGCAGCAATGGCTTTTTCTTTG | TTGAATTCCATGACATGAAGGTTG | 156 | 57 °C |
| ZSSR-188 | CTT(11) | ATCTCGTTAGTGCCTATCACCAGC | ACAGGAACATAAAAGAAGAAGAACGC | 160 | 57 °C |
| ZSSR-192 | CTG(9) | GCAGTTCTACATCATTCCCTCCTC | GAGATAGCATATCTGTTGGGTGGG | 157 | 56 °C |
| ZSSR-207 | CAT(8) | CCAAACACCAACCTTGTAATTGGT | TGTTCATGGAGACGATAAATCTGTTACT | 160 | 55 °C |
| ZSSR-209 | CAG(8) | AATGATCATGGGGAAACCAGTAGA | CTTCACTGCTCTGTTTGCTGTTGT | 153 | 57 °C |
| ZSSR-239 | ATG(12) | GCAAGTACCATACACAGGATACGTC | GCATAAAGTTTGTGGAAAACGTAATTT | 158 | 58 °C |
| ZSSR-222 | ATT(17) | GCAGCTGGATGAGAACCATAA | ACAATACAATACAAAGCCACATTAGTTC | 146 | 58 °C |
| ZSSR-247 | ATC(11) | CGCTACAAGTGTGCGATTATGAAA | GCCCAAAAGCTACAATACCTACCC | 201 | 58 °C |
| ZSSR-261 | ATA(10) | CTTGTCCAAAAGCTAACATACTTGC | AGGACAGTTCAGTAGGGTTTGTATTATT | 238 | 56 °C |
| ZSSR-262 | ATA(9) | CGTGGACCAAGTCTATACCAAAATG | TGGTTTTTCTTCTCCTAATCCATGTG | 240 | 56 °C |
| ZSSR-414 | AC(30) | TGCCACTAAAACGTATCCAATCAA | GGCATATCTGCTGAGGTGTATGTG | 232 | 56 °C |
| ZSSR-416 | AC(26) | AGGGCCAAACAAATTACAAGGATT | TTGGATGTTGAAGCTGTTTTACCTC | 151 | 57 °C |
| ZSSR-460 | AG(11) | GTTGTTCTTTTGGACCAATCCAGT | TGCATAGAACACCTTAGACAATGGAA | 240 | 56 °C |
| ZSSR-465 | CA(25) | GGTTGTGCTCATATGGGGATGTAT | TCTCCAAGATCCTTTTTGTTTTGC | 220 | 57 °C |
| ZSSR-485 | CT(28) | GTGCATGCATAAAAATCAAAACGA | GGGGTTTTATAGAAAGAGCGTGGT | 186 | 57 °C |
| ZSSR-490 | CT(23) | TTTGCTTCTTGGCTTCGACTAAAG | AGCCTACACAAAGGACTCTTTCCA | 180 | 57 °C |
| ZSSR-513 | GA(25) | TTCCTACCCCATCTGTACCTACTGT | TGGATGAATGATTGAAATGAAAAA | 223 | 57 °C |
| ZSSR-774 | AGAAA(5) | ATTGGGTTAGTCGAAAAATGGTCA | CCAATCTACAAGTGCTATGAGGCA | 153 | 57 °C |
| Swat | Dir (L) | Buner | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Traits | NO | MN | MX | Mean | Std. Devi- | % CV | MN | MX | Mean | Std. Devi- | % CV | MN | MX | Mean | Std. Devi- | % CV |
| PH | 60 | 3.7 | 75.2 | 14.73 | 13.34 | 90.56 | 3 | 25 | 8.6 | 4.72 | 54.87 | 3.5 | 5.5 | 4.36 | 0.5 | 11.46 |
| BR | 60 | 2 | 38 | 9.17 | 6.37 | 69.47 | 2 | 16 | 5.73 | 2.4 | 41.84 | 13.5 | 20 | 16.38 | 1.44 | 8.77 |
| LL | 60 | 11.8 | 83 | 32.65 | 18.77 | 57.47 | 11.2 | 62.8 | 22.65 | 11.51 | 50.81 | 7.57 | 11.57 | 9.15 | 0.91 | 9.97 |
| LW | 60 | 8.8 | 55.4 | 24.34 | 13.58 | 55.78 | 5.8 | 40.6 | 14.97 | 6.85 | 45.76 | 5.17 | 7.83 | 6.46 | 0.74 | 11.47 |
| LT | 60 | 0.13 | 4.45 | 0.42 | 0.34 | 80.83 | 0.11 | 0.6 | 0.32 | 0.12 | 36.46 | 3.6 | 6.17 | 4.79 | 0.65 | 13.58 |
| PL | 60 | 2.8 | 20.6 | 7.38 | 4.39 | 59.48 | 2.2 | 34.8 | 7.25 | 5.54 | 76.45 | 2.5 | 3.97 | 3.46 | 0.37 | 10.76 |
| InL | 60 | 2.18 | 65.4 | 17.27 | 10.45 | 60.5 | 1.42 | 25.2 | 13.84 | 5.4 | 39.05 | 1.67 | 5.33 | 3.53 | 0.73 | 20.75 |
| StD | 60 | 1.1 | 21.6 | 4.12 | 3.25 | 78.95 | 1 | 14 | 3.19 | 2.07 | 64.9 | 9.87 | 13.03 | 11.27 | 0.68 | 6.06 |
| FtW | 60 | 65 | 148 | 104 | 18.83 | 18.14 | 58.02 | 230.6 | 1.11 | 27.12 | 24.37 | 22 | 32 | 28.26 | 2.57 | 9.08 |
| FtD | 60 | 4.8 | 334.8 | 25.26 | 41.9 | 165.87 | 4.2 | 28.8 | 9.71 | 4.32 | 44.45 | 3.67 | 15.53 | 4.73 | 2.04 | 43.18 |
| FtL | 60 | 5.2 | 48.6 | 20.66 | 11.59 | 56.11 | 6.4 | 35.2 | 12.45 | 5.64 | 45.32 | 206.33 | 303.67 | 2.7 | 20.22 | 7.48 |
| Swat (ZNST) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PH | BR | LL | LW | LT | PL | InL | StD | FtW | FtD | FtL | |
| PH | 1.00 | ||||||||||
| BR | −0.04 | 1.000 | |||||||||
| LL | 0.386 ** | 0.395 ** | 1.000 | ||||||||
| LW | 0.492 ** | 0.258* | 0.859** | 1.000 | |||||||
| LT | −0.03 | 0.111 | 0.099 | 0.144 | 1.000 | ||||||
| PL | 0.07 | −0.066 | −0.122 | −0.144 | 0.166 | 1.000 | |||||
| InL | 0.10 | −0.111 | −0.335 ** | −0.341 ** | −0.144 | 0.255 | 1.000 | ||||
| StD | 0.681 ** | −0.188 | 0.111 | 0.268 * | 0.000 | 0.233 | 0.099 | 1.000 | |||
| FtW | −0.08 | −0.077 | −0.332 ** | −0.255 | 0.166 | 0.099 | 0.066 | −0.022 | 1.000 | ||
| FtD | −0.04 | 0.044 | −0.022 | 0.022 | −0.033 | −0.122 | −0.211 | −0.022 | −0.166 | 1.000 | |
| FtL | 0.20 | 0.366 ** | 0.341** | 0.368 ** | 0.111 | −0.188 | 0.022 | 0.166 | 0.199 | 0.144 | 1.000 |
| Dir (ZNDR) | |||||||||||
| PH | BR | LL | LW | LT | PL | InL | StD | FtW | FtD | FtL | |
| PH | 1.000 | ||||||||||
| BR | 0.144 | 1.000 | |||||||||
| LL | 0.044 | 0.301* | 1.000 | ||||||||
| LW | 0.155 | 0.233 | 0.862 ** | 1.000 | |||||||
| LT | 0.233 | 0.177 | 0.011 | 0.044 | 1.000 | ||||||
| PL | 0.066 | 0.022 | 0.565 ** | 0.656 ** | −0.088 | 1.000 | |||||
| InL | 0.111 | 0.155 | 0.022 | 0.000 | −0.011 | −0.244 | 1.000 | ||||
| StD | 0.529 ** | 0.364 ** | 0.313 * | 0.330 * | 0.177 | 0.200 | −0.045 | 1.000 | |||
| FtW | 0.000 | 0.077 | 0.278 * | 0.340 ** | 0.055 | 0.166 | 0.133 | −0.200 | 1.000 | ||
| FtD | 0.099 | 0.255 | 0.511 ** | 0.559 ** | −0.324* | 0.547 ** | −0.244 | 0.200 | 0.299 * | 1.000 | |
| FtL | −0.022 | 0.077 | 0.416 ** | 0.469 ** | −0.322* | 0.482 ** | −0.199 | 0.022 | 0.438 ** | 0.785 ** | 1.000 |
| Buner (ZNBU) | |||||||||||
| PH | BR | LL | LW | LT | PL | InL | StD | FtW | FtD | FtL | |
| PH | 1 | ||||||||||
| BR | −0.012 | 1.000 | |||||||||
| LL | −0.079 | 0.475 ** | 1.000 | ||||||||
| LW | 0.237 | 0.303 * | 0.298 * | 1.000 | |||||||
| LT | 0.148 | 0.508 ** | 0.397 ** | 0.329 * | 1.000 | ||||||
| PL | −0.037 | −0.076 | −0.065 | −0.263 * | −0.135 | 1.000 | |||||
| InL | −0.205 | 0.117 | 0.19 | −0.029 | 0.032 | 0.089 | 1.000 | ||||
| StD | 0.149 | −0.021 | −0.016 | 0.024 | −0.260 * | 0.061 | −0.174 | 1.000 | |||
| FtW | −0.163 | −0.018 | 0.113 | 0.072 | 0.207 | −0.093 | −0.104 | −0.300* | 1.000 | ||
| FtD | 0.364** | −0.194 | −0.129 | 0.132 | 0.005 | 0.149 | −0.329 * | 0.051 | 0.233 | 1.000 | |
| FtL | −0.285* | 0.156 | 0.252 | −0.075 | 0.058 | 0.094 | 0.352 ** | −0.253 | −0.143 | −0.602 ** | 1.000 |
| Locus | S. Size | BP | Obs_Het * | Exp_Het * | Nei ** | PIC | GD | AF | ne * | I * | na * | FIS | FIT | FST | Nm * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ZSSR-21 | 184 | 3 | 0.88 | 0.6 | 0.6 | 0.59 | 0.47 | 0.85 | 2.51 | 0.99 | 3 | −0.6 | −0.5 | 0.03 | 7.2 |
| ZSSR-93 | 268 | 3 | 0.78 | 0.6 | 0.6 | 0.55 | 0.42 | 0.76 | 2.51 | 0.99 | 3 | −0.3 | −0.29 | 0.01 | 33.77 |
| ZSSR-95 | 266 | 5 | 0.95 | 0.76 | 0.76 | 0.75 | 0.61 | 1.1 | 4.13 | 1.49 | 5 | −0.3 | −0.26 | 0.05 | 4.75 |
| ZSSR-97 | 222 | 6 | 0.48 | 0.73 | 0.72 | 0.79 | 0.53 | 0.96 | 3.62 | 1.46 | 6 | 0.17 | 0.33 | 0.19 | 1.06 |
| ZSSR-110 | 226 | 4 | 0.94 | 0.57 | 0.57 | 0.65 | 0.51 | 0.91 | 2.31 | 0.95 | 4 | −0.7 | −0.65 | 0 | 193.4 |
| ZSSR-129 | 158 | 4 | 0.22 | 0.39 | 0.39 | 0.61 | 0.36 | 0.65 | 1.64 | 0.77 | 4 | 0.41 | 0.46 | 0.09 | 2.56 |
| ZSSR-152 | 250 | 6 | 0.97 | 0.71 | 0.7 | 0.77 | 0.29 | 0.53 | 3.38 | 1.43 | 6 | −0.4 | −0.38 | 0 | 67.41 |
| ZSSR-175 | 250 | 5 | 0.11 | 0.54 | 0.54 | 0.72 | 0.55 | 0.99 | 2.16 | 0.92 | 5 | 0.82 | 0.82 | 0.01 | 33.35 |
| ZSSR-181 | 198 | 5 | 0.91 | 0.68 | 0.68 | 0.70 | 0.66 | 1.19 | 3.13 | 1.28 | 5 | −0.4 | −0.35 | 0.02 | 11.82 |
| ZSSR-183 | 186 | 4 | 0.87 | 0.57 | 0.56 | 0.67 | 0.66 | 1.19 | 2.29 | 0.94 | 4 | −0.6 | −0.6 | 0.01 | 48.42 |
| ZSSR-188 | 298 | 6 | 0.97 | 0.72 | 0.72 | 0.77 | 0.44 | 0.8 | 3.53 | 1.44 | 6 | −0.4 | −0.35 | 0 | 74.54 |
| ZSSR-192 | 238 | 6 | 0.94 | 0.73 | 0.73 | 0.73 | 0.32 | 0.57 | 3.65 | 1.48 | 6 | −0.3 | −0.31 | 0.01 | 38.24 |
| ZSSR-207 | 300 | 4 | 0.95 | 0.62 | 0.62 | 0.54 | 0.72 | 1.3 | 2.64 | 1.05 | 4 | −0.6 | −0.53 | 0.01 | 21.39 |
| ZSSR-209 | 190 | 5 | 0.16 | 0.48 | 0.47 | 0.68 | 0.49 | 0.88 | 1.9 | 0.78 | 5 | 0.68 | 0.69 | 0.05 | 4.49 |
| ZSSR-239 | 236 | 3 | 0.62 | 0.51 | 0.51 | 0.59 | 0.41 | 0.73 | 2.03 | 0.74 | 3 | −0.2 | −0.22 | 0.02 | 14.84 |
| ZSSR-222 | 262 | 5 | 0.33 | 0.59 | 0.59 | 0.72 | 0.58 | 1.04 | 2.44 | 1.18 | 5 | 0.16 | 0.38 | 0.26 | 0.71 |
| ZSSR-247 | 128 | 3 | 0.44 | 0.53 | 0.52 | 0.59 | 0.65 | 1.17 | 2.1 | 0.9 | 3 | 0.12 | 0.14 | 0.02 | 13.07 |
| ZSSR-261 | 290 | 3 | 0.34 | 0.31 | 0.3 | 0.47 | 0.51 | 0.92 | 1.44 | 0.58 | 3 | −0.1 | −0.13 | 0.01 | 30.84 |
| ZSSR-262 | 278 | 3 | 0.86 | 0.61 | 0.61 | 0.56 | 0.41 | 0.74 | 2.55 | 1 | 3 | −0.4 | −0.4 | 0.01 | 16.51 |
| ZSSR-414 | 200 | 3 | 0.07 | 0.14 | 0.14 | 0.47 | 0.51 | 0.91 | 1.16 | 0.3 | 3 | 0.47 | 0.5 | 0.05 | 4.51 |
| ZSSR-416 | 138 | 3 | 0.68 | 0.67 | 0.66 | 0.55 | 0.58 | 1.05 | 2.97 | 1.09 | 3 | −0.1 | −0.04 | 0.06 | 3.68 |
| ZSSR-460 | 210 | 4 | 0.92 | 0.69 | 0.69 | 0.65 | 0.34 | 0.61 | 3.22 | 1.26 | 4 | −0.4 | −0.34 | 0.01 | 37.52 |
| ZSSR-465 | 248 | 4 | 0.96 | 0.63 | 0.62 | 0.65 | 0.48 | 0.87 | 2.66 | 1.07 | 4 | −0.5 | −0.54 | 0 | 92.14 |
| ZSSR-485 | 176 | 3 | 0.57 | 0.49 | 0.49 | 0.50 | 0.52 | 0.93 | 1.95 | 0.8 | 3 | −0.2 | −0.2 | 0.02 | 16.04 |
| ZSSR-490 | 224 | 2 | 0.21 | 0.24 | 0.24 | 0.33 | 0.29 | 0.52 | 1.31 | 0.4 | 2 | 0.09 | 0.13 | 0.05 | 4.48 |
| ZSSR-513 | 192 | 4 | 0.88 | 0.73 | 0.73 | 0.65 | 0.28 | 0.51 | 3.68 | 1.34 | 4 | −0.2 | −0.2 | 0.03 | 9.48 |
| ZSSR-774 | 300 | 5 | 0.89 | 0.68 | 0.68 | 0.75 | 0.55 | 0.99 | 3.14 | 1.3 | 5 | −0.3 | −0.3 | 0 | 57.96 |
| Mean | 227 | 4.43 | 0.662 | 0.575 | 0.572 | 0.671 | 0.494 | 0.89 | 2.6 | 1.03 | 4.11 | −0.2 | −0.17 | 0.04 | 6.415 |
| Source | Df | SS | Component of Variance | Percentage of Variation | Fixation Index | p-Value |
|---|---|---|---|---|---|---|
| Among Populations | 2 | 82.985 | 0.321 | 8% | FST 0.092 | 0.001 |
| Among Individuals within Population | 176 | 565.959 | 0.453 | 12% | FIS 0.102 | 0.001 |
| Within Individual | 179 | 559.000 | 3.123 | 80% | FIT 0.105 | 0.001 |
| Total | 357 | 1207.944 | 3.490 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uddin, N.; Muhammad, N.; Ali, S.S.; Ullah, R.; Bari, A.; Hussain, H.; Zhu, D. Characterization of the Genetic Variability within Ziziphus nummularia Genotypes by Phenotypic Traits and SSR Markers with Special Reference to Geographic Distribution. Genes 2023, 14, 155. https://doi.org/10.3390/genes14010155
Uddin N, Muhammad N, Ali SS, Ullah R, Bari A, Hussain H, Zhu D. Characterization of the Genetic Variability within Ziziphus nummularia Genotypes by Phenotypic Traits and SSR Markers with Special Reference to Geographic Distribution. Genes. 2023; 14(1):155. https://doi.org/10.3390/genes14010155
Chicago/Turabian StyleUddin, Nisar, Noor Muhammad, Sameh Samir Ali, Riaz Ullah, Ahmed Bari, Hidayat Hussain, and Daochen Zhu. 2023. "Characterization of the Genetic Variability within Ziziphus nummularia Genotypes by Phenotypic Traits and SSR Markers with Special Reference to Geographic Distribution" Genes 14, no. 1: 155. https://doi.org/10.3390/genes14010155
APA StyleUddin, N., Muhammad, N., Ali, S. S., Ullah, R., Bari, A., Hussain, H., & Zhu, D. (2023). Characterization of the Genetic Variability within Ziziphus nummularia Genotypes by Phenotypic Traits and SSR Markers with Special Reference to Geographic Distribution. Genes, 14(1), 155. https://doi.org/10.3390/genes14010155