
Genomic Selection for Dairy Cattle Behaviour Considering Novel Traits in a Changing Technical Production Environment


Abstract
:1. Introduction
2. Components of Dairy Cattle Behaviour
2.1. Feeding, Activity and Reproductive Behaviour
2.2. Behaviour Response to Human Handling: Cow–Human Interactions
2.3. Social and Maternal Behaviour: Cow–Cow and Cow–Calf Interactions

{kind=link}
{kind=link}
{kind=link}
| Behaviour Trait | Interaction | Score | Reference |
|---|---|---|---|
| Avoidance distance at the feeding barrier | Cow–human | Distance [cm] | [41] |
| Avoidance distance in the barn | Cow–human | Distance [cm] | [43] |
| Tolerance to tactile interaction | Cow–human | 1 to 5 | [9] |
| Release behaviour after opening the feeding gate | Cow–human | 1 to 5 | [9] |
| Agonistic behaviour | Cow–cow | Number of aggressive behaviours per animal and time period | [41] |
| Cohesive behaviour | Cow–cow | Number of cohesive behaviours per animal and time period | [41] |
| Early postpartum behaviour of cow and calf | Cow–calf | Duration and frequency | [60] |
2.4. Behaviour Response to the Technical Environment: Behaviour in (Automatic) Milking Systems and Learning Behaviour
3. The Relationship between Behaviour and Milk Production, Reproduction, and Health
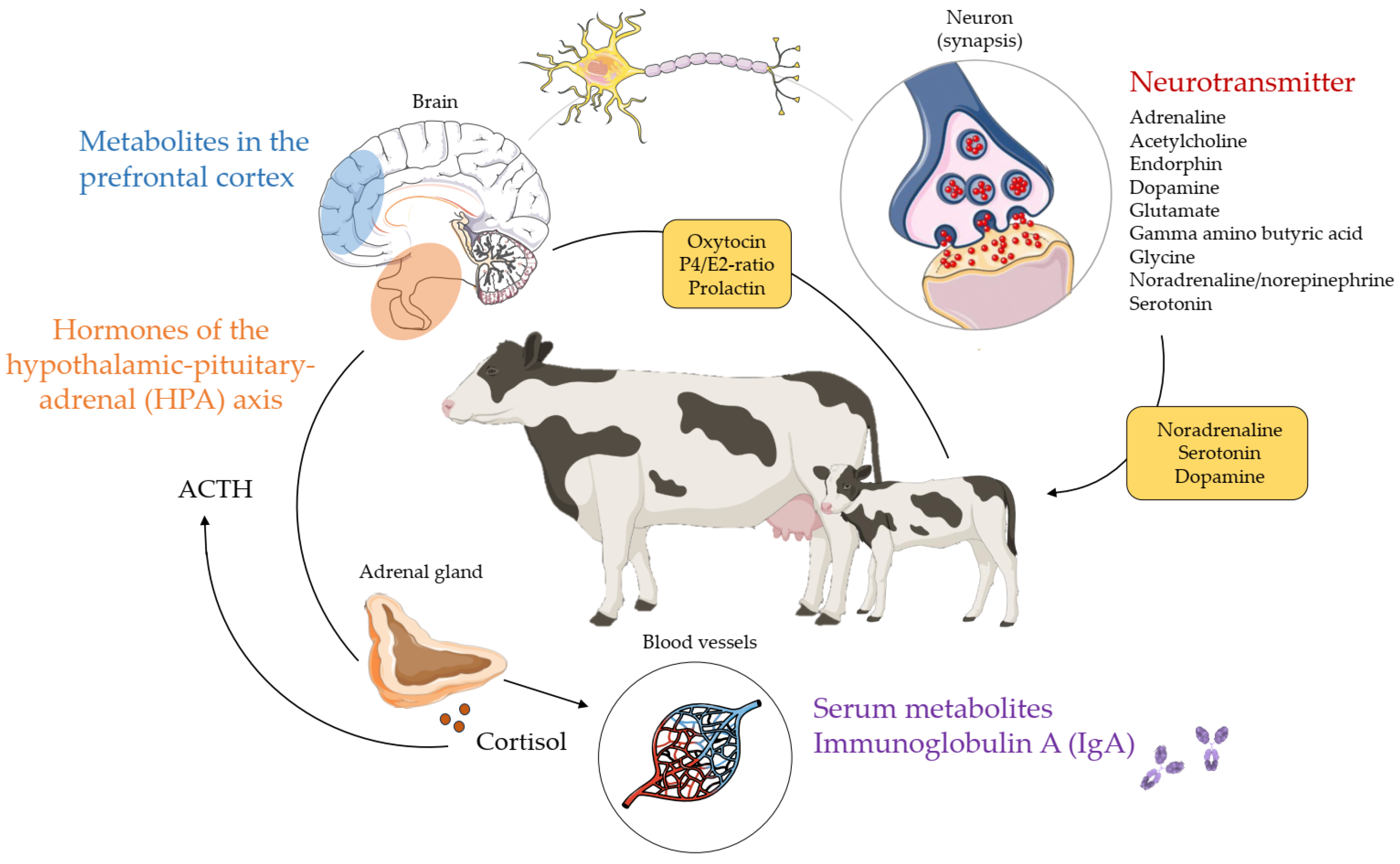
4. Biomarkers as Indicators for Cattle Behaviour
4.1. Physiological Biomarkers
4.2. Molecular Biomarkers
5. Genetic Parameter Estimates for Dairy Cattle Behaviour Traits and Genetic Correlations with Other Traits
5.1. Feeding, Activity, and Reproductive Behaviour
5.2. Behaviour Response to Human Handling
5.3. Social and Maternal Behaviour
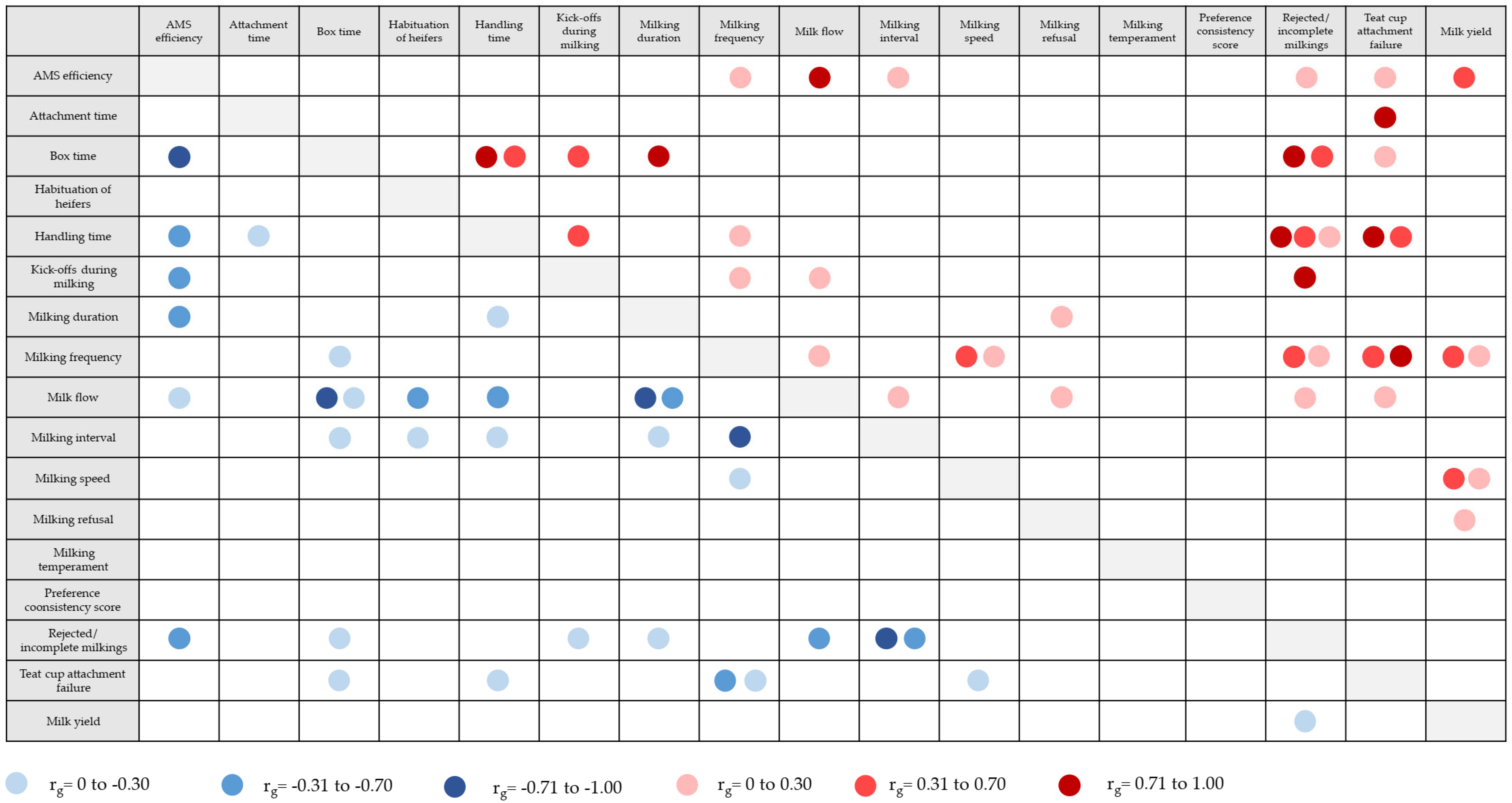
5.4. Behaviour Traits Derived from (Automatic) Milking Systems
| Trait | Definition | Link to Behaviour | Heritability | Reference |
|---|---|---|---|---|
| AMS efficiency | Milk production in kg milk produced per total box time | Cow robot performance | 0.29 | [148] |
| 0.33 | [144] | |||
| 0.40–0.50 | [149] | |||
| 0.45–0.56 | [63] | |||
| 0.23 | [147] | |||
| 0.29 | [150] | |||
| 0.22 | [64] | |||
| Attachment time | Time used for attaching the milking equipment | Cow robot performance | 0.26 | [10] |
| 0.36 | [146] | |||
| Box time | Time between entering and leaving the AMS | Milking behaviour | 0.41 | [144] |
| 0.06–0.33 | [151] | |||
| 0.27 | [64] | |||
| Habituation of heifers | Time period a heifer needs to become familiar with the AMS | Learning/habituation behaviour | 0.07 | [147] |
| Handling time | Time in AMS before and after milking | Temperament | 0.05–0.15 | [145] |
| 0.05 | [64] | |||
| Kick-offs during milking | Number of knocking off the milking device | Temperament, udder health | 0.06 | [152] |
| 0.03 | [138] | |||
| 0.06 | [64] | |||
| Milking duration/ milking time | Milking duration of each AMS visit in minutes | Milking behaviour | 0.39 | [153] |
| 0.19 | [138] | |||
| 0.22–0.28 | [63] | |||
| 0.32 | [10] | |||
| Milking frequency | AMS visits per cow per day | Cow robot performance, milking behaviour, learning behaviour | 0.23 | [154] |
| 0.02–0.07 | [141] | |||
| 0.26 | [144] | |||
| 0.16–0.27 | [142] | |||
| 0.12–0.28 | [151] | |||
| 0.02–0.08 | [143] | |||
| 0.51 | [10] | |||
| 0.05 | [64] | |||
| Milk flow | Average milk flow in kg min−1 | Milking behaviour, temperament | 0.43–0.52 | [63] |
| 0.25 | [138] | |||
| 0.48 | [64] | |||
| Milking interval | Interval between milking sessions | Milking behaviour, Temperament, social dominance | 0.09–0.26 | [141] |
| 0.17 | [152] | |||
| 0.07 | [138] | |||
| 0.08 | [147] | |||
| 0.02 | [64] | |||
| Milking speed | Milk yield per milk time | Milking behaviour | 0.42 | [154] |
| 0.43 | [10] | |||
| 0.46 | [150] | |||
| Milking refusal | The cow is not allowed to be milked because the expected milk yield is too low | Temperament, learning behaviour | 0.02 | [63] |
| Milking temperament | Aggressive or docile behaviour during milking | Temperament | 0.14 | [152] |
| Preference consistency score | Milking box preference consistency/frequency of access to each milking unit in a given time period | Learning/habituation behaviour | 0.05–0.13 | [151] |
| Rejected/incomplete milkings | Number of unsuccessful milkings | Temperament | 0.02–0.06 | [145] |
| 0.01–0.02 | [64] | |||
| Teat cup attachment failure/ number of attachments per teat | Teat cup attachment failure | Cow robot performance | 0.21–0.31 | [145] |
| 0.06 | [152] | |||
| 0.26 | [146] | |||
| 0.002 | [64] |
6. Genomic Regions Associated with Dairy Cattle Behaviour
6.1. Feeding, Activity and Reproductive Behaviour
6.2. Behaviour Response to Human Handling
6.3. Social and Maternal Behaviour
| BTA | Trait | Chromosome Position | Associated Gene/QTL 1 | Breed | Reference |
|---|---|---|---|---|---|
| 1 | Rank order in the herd | 126.5 Mbp | SLC9A9 | Brown Swiss | [135] |
| Standing alert in response to social separation | 0.4–0.6 Mbp | BM6438 | HF × Charolais crosses | [169] | |
| Vocalisation in response to social separation | 95.7–95.9 Mbp | MBMS4044 | HF × Charolais crosses | [169] | |
| 3 | Refusals per day in AMS | 73.5–73.9 Mbp | NEGR1, PTGER3 | HF | [175] |
| 4 | General temperament | 14.9 bp | TAC1 | Brown Swiss | [135] |
| Standing alert in response to social separation | 51.3–86.3 Mbp | MAF50-DIK026 | HF × Charolais crosses | [169] | |
| Vocalisation in response to social separation | 51.3–86.3 Mbp | MAF50-DIK026 | HF × Charolais crosses | [169] | |
| 6 | Rank order in the herd | 8.4 Mbp | - | Brown Swiss | [135] |
| Walking in response to social separation | 4.0–31.0 Mbp | DIK5076-BM1329 | HF × Charolais crosses | [169] | |
| 7 | Vocalisation in response to social separation | 62.0–62.2 Mbp | INRA053 | HF × Charolais crosses | [169] |
| 8 | Aggressiveness | 58.5 Mbp | - | Brown Swiss | [135] |
| Walking in response to social separation | 51.9–52.0 Mbp | CSSM047 | HF × Charolais crosses | [169] | |
| General temperament | 101.5 bp | AKAP2, TXN, TXNDC8 | Brown Swiss | [135] | |
| 9 | Vocalisation in response to social separation | 28.1–45.4 Mbp | BM2504-UWCA9 | HF × Charolais crosses | [169] |
| 10 | Vocalisation in response to social separation | 20.9–38.0 Mbp | BMS528-TGLA378 | HF x Charolais crosses | [169] |
| Walking in response to social separation | 52.2–67.7 Mbp | BM888-CSRM60 | HF × Charolais crosses | [169] | |
| 11 | Rumination | 55,229,674 bp | - | HF | [133] |
| Standing alert in response to social separation | 48.3–66.9 Mbp | ILSTS100-IDVGA-3 | HF × Charolais crosses | [169] | |
| 12 | Refusals per day in AMS | 53.0–55.0 Mbp | SPRY2, POU4F1 | HF | [175] |
| 84.3 Mbp | MYO16, CARS2, IRS2 | HF | [175] | ||
| 13 | Not active | 79,178,395 bp | - | Multi-breed | [133] |
| 16 | Standing alert in response to social separation | 87.7–101.8 Mbp | HUJ625 | HF × Charolais crosses | [169] |
| Vocalisation in response to social separation | 46.4–66.5 Mbp | ETH11-BM719 | HF × Charolais crosses | [169] | |
| Walking in response to social separation | 7.6–7.8 Mbp | BM121 | HF × Charolais crosses | [169] | |
| 17 | Rumination | 68,187,177 bp | - | Multi-breed | [133] |
| 18 | Rank order in the herd | 29.4 Mbp | - | Brown Swiss | [135] |
| Refusals per day in AMS | 25.4–26.2 Mbp | COQ9, CNOT1 | HF | [175] | |
| 55.7–55.9. Mbp | SLC6A16, PTH2, ALDH16A1 | HF | [175] | ||
| 60.0–63.2 Mbp | MBOAT7, CNOT3 | HF | [175] | ||
| Vocalisation in response to social separation | 0–15.8 cM | IDVGA-31-ABS013 | HF × Charolais crosses | [169] | |
| 19 | Rank order in the herd | 14.1 Mbp | ACACA, TADA2A, DUSP14, SYNRG | Brown Swiss | [135] |
| Refusals per day in AMS | 50.5–50.6 Mbp | TBCD, ACTG1, RAC3 | HF | [175] | |
| Standing alert in response to social separation | 40.4–52.1 Mbp | CSSM065-ETH3 | HF × Charolais crosses | [169] | |
| Vocalisation in response to social separation | 40.4–52.1 Mbp | CSSM065-ETH3 | HF × Charolais crosses | [169] | |
| Walking in response to social separation | 25.1–40.4 Mbp | BMS2142-CSSM065 | HF × Charolais crosses | [169] | |
| 20 | Aggressiveness | 65.7 Mbp | ADCY | Brown Swiss | [135] |
| Flight from feeder | 45.6–62.3 Mbp | DIK15-BM5004 | HF × Charolais crosses | [169] | |
| 23 | Feeding | 19,834,215 bp | SLC25A27 | Multi-breed | [133] |
| 25 | Flight from feeder | 19.6–33.0 Mbp | BM737-INRA222 | HF × Charolais crosses | [169] |
| Vocalisation in response to social separation | 19.6–33.0 Mbp | BM737-INRA222 | HF × Charolais crosses | [169] | |
| 26 | Vocalisation in response to social separation | 1.6–19.6 Mbp | ABS12-HEL11 | HF × Charolais crosses | [169] |
| 27 | Rumination | 37,283,994 bp | THAP1, RNF170, | Multi-breed | [133] |
| 28 | Flight from feeder | 6.2–6.4 Mbp | BP23 | HF × Charolais crosses | [169] |
| 29 | Flight from feeder | 29.4–51.1 Mbp | DIK094-MNB101 | HF × Charolais crosses | [169] |
| Rumination | 46,014,507 bp | RPS6KB2, PTPRCAP, CORO1B, GPR152, CaBP4, TMEM134, AIP, PITPNM1 | Multi-breed | [133] | |
| 49,036,680 bp | ENSBTAG00000000776, MRGPRG | Multi-breed | [133] | ||
| Vocalisation in response to social separation | 17.9–24.6 Mbp | RM044-MNB-166 | HF × Charolais crosses | [169] |
6.4. Behaviour Traits Derived from (Automatic) Milking Systems
7. Genetics and Genomics of Biomarkers as Indicators for Cattle Behaviour
8. Inclusion of Behaviour Traits into Genomic Selection Indices
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bikker, J.P.; van Laar, H.; Rump, P.; Doorenbos, J.; van Meurs, K.; Griffioen, G.M.; Dijkstra, J. Technical note: Evaluation of an ear-attached movement sensor to record cow feeding behavior and activity. J. Dairy Sci. 2014, 97, 2974–2979. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, M.A.; Rogers, A.R.; Verkerk, G.A. The effect of temperament and responsiveness towards humans on the behavior, physiology and milk production of multi-parous dairy cows in a familiar and novel milking environment. Physiol. Behav. 2012, 107, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Haskell, M.J.; Simm, G.; Turner, S.P. Genetic selection for temperament traits in dairy and beef cattle. Front. Genet. 2014, 5, 368. [Google Scholar] [CrossRef] [PubMed]
- Burrow, H.M. Measurements of temperament and their relationships with performance traits of beef cattle. Anim. Breed. Abstr. 1997, 65, 477–495. [Google Scholar]
- Hemsworth, P.H. Human-animal interactions in livestock production. Appl. Anim. Behav. Sci. 2003, 81, 185–198. [Google Scholar] [CrossRef]
- de Koning, C.J.A.M. Automatic milking—Common practice on dairy farms. In Proceedings of the First North American Conference on Precision Dairy Management, Toronto, ON, Canada, 2–5 March 2010; pp. 1–16. [Google Scholar]
- Hiendleder, S.; Thomsen, H.; Reinsch, N.; Bennewitz, J.; Leyhe-Horn, B.; Looft, C.; Xu, N.; Medjugorac, I.; Russ, I.; Kühn, C.; et al. Mapping of QTL for body conformation and behavior in cattle. J. Hered. 2003, 94, 496–506. [Google Scholar] [CrossRef]
- Chang, Y.; Brito, L.F.; Alvarenga, A.B.; Wang, Y. Incorporating temperament traits in dairy cattle breeding programs: Challenges and opportunities in the phenomics era. Anim. Front. 2020, 10, 29–36. [Google Scholar] [CrossRef]
- Ebinghaus, A.; Ivemeyer, S.; Rupp, J.; Knierim, U. Identification and development of measures suitable as potential breeding traits regarding dairy cows’ reactivity towards humans. Anim. Behav. Sci. 2016, 185, 30–38. [Google Scholar] [CrossRef]
- Piwczyński, D.; Sitkowska, B.; Ptak, E. Genetic relationship among somatic cell score and some milking traits in Holstein-Friesian primiparous cows milked bay an automated milking system. Animal 2021, 15, 100094. [Google Scholar] [CrossRef]
- Titterington, F.M.; Knox, R.; Morrison, S.J.; Shirali, M. Behavioural traits in Bos taurus cattle, their heritability, potential genetic markers, and associations with production traits. Animals 2022, 12, 2602. [Google Scholar] [CrossRef]
- Titterington, F.M.; Knox, R.; Buijs, S.; Lowe, D.E.; Morrison, S.J.; Lively, F.O.; Shirali, M. Human-animal interactions with Bos taurus cattle and their impacts on On-Farm safety: A systematic review. Animals 2022, 12, 776. [Google Scholar] [CrossRef] [PubMed]
- Sahu, B.K.; Parganiha, A.; Pati, A.K. Behavior and foraging ecology of cattle. J. Vet. Behav. 2020, 40, 50–74. [Google Scholar] [CrossRef]
- Albright, J.L. Feeding Behavior of Dairy Cattle. Nutr. Feed. Calves 1993, 76, 485–498. [Google Scholar] [CrossRef]
- de Carvalho, E.R.; Schmelz-Roberts, N.S.; White, H.M.; Wilcox, S.; Eicher, S.; Donkin, S.S. Eating, resting and rumination activities of transition dairy cows fed with glycerol. Glob. Sci. Technol. 2014, 7, 130–141. [Google Scholar] [CrossRef]
- Custido, S.A.S.; Tomaz, M.P.P.; da Silva, D.A.L.; de Oliviera Goulart, R.; Dias, K.M.; de Carvalho, E.R. Feeding behaviour of beef cattle fed different forages and housed in individual or collective pens. J. Anim. Behav. Biometeorol. 2017, 5, 20–28. [Google Scholar] [CrossRef]
- von Keyserlingk, M.A.G.; Weary, D.M. Review: Feeding behaviour of dairy cattle: Measures and applications. Can. J. Anim. Sci. 2010, 90, 303–309. [Google Scholar] [CrossRef]
- Bozkurt, Y.; Ozkaya, S.; Dewi, I.A. Association between aggressive behaviour and high-energy feeding level in beef cattle. J. Anim. Sci. 2006, 51, 151–156. [Google Scholar] [CrossRef]
- Grant, R.J.; Albright, J.L. Feeding behaviour and management factors during the transition period in dairy cattle. J. Anim. Sci. 1995, 73, 2791–2803. [Google Scholar] [CrossRef]
- Strickling, W.R.; Gonyou, H.W. Dominance and eating behaviour of beef cattle fed from single stall. Appl. Anim. Ethol. 1981, 7, 135–140. [Google Scholar] [CrossRef]
- Grant, R.J.; Albright, J.L. Effect of animal grouping on feeding behaviour and intake of dairy cattle. J. Dairy. Sci. 2001, 84, 156–163. [Google Scholar] [CrossRef]
- DeVries, T.J.; von Keyserlingk, M.A.G.; Weary, D.M. Effect of feeding space on the inter-cow distance, aggression, and feeding behaviour of free-stall housed lactating dairy cows. J. Dairy Sci. 2004, 87, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Huzzey, J.M.; DeVries, T.J.; Valois, P.; von Keyserlingk, M.A.G. Stocking Density and Feed Barrier Design Affect the Feeding and Social Behavor of Dairy Cattle. J. Dairy Sci. 2006, 89, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, M.; Brügemann, K.; Brandt, H.; König, S. Associations between precision sensor data with productivity, health and welfare indicator traits in native black and white dual-purpose cattle under grazing conditions. Appl. Anim. Behav. Sci. 2019, 212, 9–18. [Google Scholar] [CrossRef]
- Cavani, L.; Brown, W.E.; Parker Gaddis, K.L.; Tempelman, R.J.; VandeHaar, M.J.; White, H.M.; Peñagaricano, F.; Weigel, K.A. Estimates of genetic parameters for feeding behavior traits and their associations with feed efficiency in Holstein cows. J. Dairy Sci. 2022, 105, 7564–7574. [Google Scholar] [CrossRef] [PubMed]
- Legerwood, D.N.; Winckler, C.; Tucker, C.B. Evaluation of data loggers, sampling intervals, and editing techniques for measuring the lying behavior of dairy cattle. J. Dairy Sci. 2010, 93, 5129–5139. [Google Scholar] [CrossRef] [PubMed]
- Büchel, S.; Sundrum, A. Evaluation of a new system for measuring feeding behavior of dairy cows. Comput. Electron. Agric. 2014, 108, 12–16. [Google Scholar] [CrossRef]
- Kuan, C.Y.; Tsai, Y.C.; Hsu, J.T.; Ding, S.T.; Te Lin, T. An imaging system based on deep learning for monitoring the feeding behavior of dairy cows. In Proceedings of the ASABE Annual International Meeting, Boston, MA, USA, 7–10 July 2019; American Society of Agricultural and Biological Engineers: Saint Joseph, MI, USA, 2019. [Google Scholar]
- Drissler, M.; Gaworski, M.; Tucker, C.B.; Weary, D.M. Effects on lying behavior of dairy cattle. J. Dairy Sci. 2005, 88, 2381–2387. [Google Scholar] [CrossRef]
- Alsaaod, M.; Fadul, M.; Steiner, A. Automatic lameness detection in cattle. Vet. J. 2019, 246, 34–44. [Google Scholar] [CrossRef]
- Quiao, Y.; Kong, H.; Clark, C.; Lomax, S.; Su, D.; Eiffert, S.; Sukkarieh, S. Intelligent Perception-Based Cattle Lameness Detection and Behaviour Recognition: A Review. Animals 2021, 11, 3033. [Google Scholar] [CrossRef]
- Aungier, S.P.M.; Roche, J.F.; Sheehy, M.; Crowe, M.A. Effects of management and health on the use of activity monitoring for estrus detection in dairy cows. J. Dairy Sci. 2012, 95, 2452–2466. [Google Scholar] [CrossRef]
- Løvendahl, P.; Chagunda, M.G.G. On the use of physical activity monitoring for estrus detection in dairy cows. J. Dairy Sci. 2010, 93, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Lucy, M.C. Symposium review: Selection for fertility in the modern dairy cow—Current status and future direction for genetic selection. J. Dairy Sci. 2019, 102, 3706–3721. [Google Scholar] [CrossRef] [PubMed]
- Dutt, T.; Sinha, R.R.K.; Singh, M.; Bharti, P.K.; Patel, B.H.M.; Gaur, G.K. Effect of temperament on production, reproduction and milking behaviour of Vrindavani cows in an organized herd. Indian J. Anim. Sci. 2016, 86, 950–952. [Google Scholar] [CrossRef]
- Boissy, A.; Fisher, A.D.; Bouix, J.; Hinch, G.N.; Le Neindre, P. Genetics of fear in ruminant livestock. Livest. Prod. Sci. 2005, 93, 23–32. [Google Scholar] [CrossRef]
- Ivemeyer, S.; Knierim, U.; Waiblinger, S. Effect of human-animal relationship and management on udder health in Swiss dairy herds. J. Dairy Sci. 2011, 94, 5890–5902. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Coleman, G.J.; Barnett, J.L.; Borg, S. Relationship between human-animal interactions and productivity of commercial dairy cows. J. Anim. Sci. 2000, 78, 2821–2831. [Google Scholar] [CrossRef]
- Chesterton, R.N.; Pfeiffer, D.U.; Morris, R.S.; Tanner, C.M. Environmental and behavioural factors affecting the prevalence of foot lameness in New Zealand dairy herds—A case-control study. N. Z. Vet. J. 1989, 37, 135–142. [Google Scholar] [CrossRef]
- Waiblinger, S.; Boivin, X.; Pedersen, V.; Tosi, M.V.; Janczak, A.M.; Visser, E.K.; Jones, R.B. Assessing the human-animal relationship in farmed species: A critical review. Appl. Anim. Behav. Sci. 2006, 101, 185–242. [Google Scholar] [CrossRef]
- Welfare Quality®. Welfare Quality® Assessment Protocol for Cattle; Welfare Quality® Consortium: Lelystad, The Netherlands, 2009. [Google Scholar]
- Nicol, C. Methods of welfare assessment-indices based on animal behaviour. In Proceedings of the International Conference “Welfare in Farm Animals”, Jastrzębiec, Poland, 7–8 June 2011; pp. 17–28. [Google Scholar]
- Waiblinger, S.; Menke, C.; Coleman, G. The relationship between attitudes, personal characteristics and behaviour of stockpeople and subsequent behaviour and production of dairy cows. Appl. Anim. Behav. Sci. 2002, 79, 195–219. [Google Scholar] [CrossRef]
- Waiblinger, S.; Menke, C.; Fölsch, D.W. Influences on the avoidance and approach behaviour of dairy cows towards humans on 35 farms. Appl. Anim. Behav. Sci. 2003, 84, 23–39. [Google Scholar] [CrossRef]
- Hoppe, S.; Brandt, H.R.; König, S.; Erhardt, G.; Gauly, M. Temperament traits of beef calves measured under field conditions and their relationships to performance. J. Anim. Sci. 2010, 88, 1982–1989. [Google Scholar] [CrossRef]
- Gargiulo, J.I.; Eastwood, C.R.; Garcia, S.C.; Lyons, N.A. Dairy farmers with larger herd sizes adopt more precision dairy technologies. J. Dairy Sci. 2018, 101, 5466–5473. [Google Scholar] [CrossRef]
- Raussi, S. Human-cattle interactions in group housing. Appl. Anim. Behav. Sci. 2003, 80, 245–262. [Google Scholar] [CrossRef]
- Jacobs, J.A.; Siegford, J.M. Invited review: The impact of automatic milking systems on dairy cow management, behavior, health, and welfare. J. Dairy Sci. 2012, 95, 2227–2247. [Google Scholar] [CrossRef] [PubMed]
- Breuer, K.; Hemsworth, P.H.; Barnett, J.L.; Matthews, L.R.; Coleman, G.J. Behavioural response to humans and the productivity of commercial dairy cows. Appl. Anim. Behav. Sci. 2000, 66, 273–288. [Google Scholar] [CrossRef] [PubMed]
- BØe, K.E.; Færevik, G. Grouping and social preferences in calves, heifers and cows. Appl. Anim. Behav. Sci. 2003, 80, 175–190. [Google Scholar] [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Burdick, N.C.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Willard, S.T.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Relationships between temperament and transportation with rectal temperature and serum concentrations of cortisol and epinephrine in bulls. Livest. Sci. 2010, 129, 166–172. [Google Scholar] [CrossRef]
- Foris, B.; Zebunke, M.; Langbein, J.; Melzer, N. Comprehensive analysis of affiliative and agonistic social networks in lactating dairy cattle groups. Appl. Anim. Behav. Sci. 2019, 210, 60–67. [Google Scholar] [CrossRef]
- Edwards, S.A. The behaviour of dairy cows and their newborn calves in individual or group housing. Appl. Anim. Ethol. 1983, 10, 191–198. [Google Scholar] [CrossRef]
- Hasegawa, N.; Nishiwaki, A.; Sugawara, K.; Ito, I. The effects of social exchange between two groups of lactating primiparous heifers on milk production, dominance order, behaviour and adrenocortical response. Appl. Anim. Behav. Sci. 1997, 51, 15–27. [Google Scholar] [CrossRef]
- Landaeta-Hernández, A.J.; Palomares-Naveda, R.; Soto-Castillo, G.; Atencio, A.; Chase, C.C., Jr.; Chenoweth, P.J. Social and Breed Effects on the Expression of a PGF2α Induced Oestrus in Beef Cows. Reprod. Dom. Anim. 2004, 39, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.M.; Lawrence, A.B.; Haskell, M.J. Measuring sociability in dairy cows. Appl. Anim. Behav. Sci. 2010, 122, 84–91. [Google Scholar] [CrossRef]
- Kondo, S.; Hurnik, J.F. Stabilization of social hierarchy in dairy cows. Appl. Anim Behav. Sci. 1990, 27, 287–297. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Herrera-Covarrubias, D.; García, L.I.; Toledo, R.; Hernández, M.E.; Paredes-Ramos, P.; Corona-Morales, A.A.; Manzo, J. Neurobiology of Maternal Behavior in Nonhuman Mammals: Acceptance, Recognition, Motivation, and Rejection. Animals 2022, 12, 3589. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B. The early behaviour of cow and calf in an individual calving pen. Appl. Anim. Behav. Sci. 2011, 134, 92–99. [Google Scholar] [CrossRef]
- Meagher, R.K.; Beaver, A.; Weary, D.M.; von Keyserlingk, A.G. Invited review: A systematic review of the effects of prolonged cow-calf contact on behavior, welfare, and productivity. J. Dairy Sci. 2019, 102, 5765–5783. [Google Scholar] [CrossRef]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behaviour. Net. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Pedrosa, V.B.; Boerman, J.P.; Gloria, L.S.; Chen, S.-Y.; Montes, M.E.; Doucette, J.S.; Brito, L.F. Genomic-based genetic parameters for milkability traits derived from automatic milking systems in North American Holstein cattle. J. Dairy Sci. 2023, 106, 2613–2629. [Google Scholar] [CrossRef]
- Wethal, K.B.; Heringstad, B. Genetic analyses of novel temperament and milkability traits in Norwegian Red cattle based on data from automatic milking systems. J. Dairy Sci. 2019, 102, 8221–8233. [Google Scholar] [CrossRef]
- Kilgour, R. The open-field test as an assessment of the temperament of dairy cows. Anim. Behav. 1975, 23, 615–624. [Google Scholar] [CrossRef]
- Bach, A.; Devant, M.; Igleasias, C.; Ferrer, A. Forced traffic in automatic milking systems effectively reduces the need to get cows, but alters eating behaviour and does not improve milk yield of dairy cattle. J. Dairy Sci. 2009, 92, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Melin, M.; Pettersson, G.; Svennersten-Sjaunja, K.; Wiktorsson, H. The effects of restricted feed access and social rank on feeding behavior, ruminating and intake for cows managed in automated milking systems. Appl. Anim. Behav. Sci. 2007, 197, 13–21. [Google Scholar] [CrossRef]
- Marino, L.; Allen, K. The psychology of cows. Anim. Behav. Cognit. 2017, 4, 474–498. [Google Scholar] [CrossRef]
- Hagen, K.; Broom, D.M. Cattle discriminate between individual familiar herd members in a learning experiment. Appl. Anim. Behav. Sci. 2003, 82, 13–28. [Google Scholar] [CrossRef]
- Hagen, K.; Broom, D.M. Emotional reactions to learning in cattle. Appl. Anim. Behav. Sci. 2004, 85, 203–213. [Google Scholar] [CrossRef]
- Meagher, R.; Strazhnik, E.; von Keyserlingk, M.A.; Weary, D.M. Assessing the motivation to learn in cattle. Sci. Rep. 2020, 10, 6847. [Google Scholar] [CrossRef]
- Cziszter, L.T.; Gavojdian, D.; Neamt, R.; Neciu, F.; Kusza, S.; Ilie, D.-E. Effects of temperament on production and reproductive performances in Simmental dual-purpose cows. J. Vet. Behav. 2016, 15, 50–55. [Google Scholar] [CrossRef]
- Mincu, M.; Gavojdian, D.; Nicolae, I.; Olteanu, A.-C.; Vlagioiu, C. Effects of milking temperament of dairy cows on production and reproductive efficiency under tied stall housing. J. Vet. Behav. 2021, 44, 12–17. [Google Scholar] [CrossRef]
- Marçal-Pedroza, M.G.; Campos, M.M.; Pereira, L.G.R.; Machado, F.S.; Tomich, T.R.; Paranhos da Costa, M.J.R.; Sant Anna, A.C. Consistency of temperament traits and their relationship with milk yield in lactating primiparous F1 Holstein—Gyr cows. Appl. Anim. Behav. Sci. 2020, 222, 104881. [Google Scholar] [CrossRef]
- Sawa, A.; Bogucki, M.; Neja, W.; Krężel-Czopek, S. Effect of temperament on performance of primiparous dairy cows. Ann. Anim. Sci. 2017, 17, 863–872. [Google Scholar] [CrossRef]
- Ebinghaus, A.; Ivemeyer, S.; Lauks, V.; Santos, L.; Brügemann, K.; König, S.; Knierim, U. How to measure dairy cows’ responsiveness towards humans in breeding and welfare assessment? A comparison of selected behavioural measures and existing breeding traits. Appl. Anim. Behav. Sci. 2017, 196, 22–29. [Google Scholar] [CrossRef]
- Dittrich, I.; Gertz, M.; Krieter, J. Alterations in sick dairy cows‘ daily behavioural patterns. Heliyon 2019, 5, e02902. [Google Scholar] [CrossRef] [PubMed]
- Siivonen, J.; Taponen, S.; Hovinen, M.; Pastell, M.; Lensink, B.J.; Pyörälä, S.; Hänninen, L. Impact of acute clinical mastitis on cow behavior. Appl. Anim. Behav. Sci. 2011, 132, 101–106. [Google Scholar] [CrossRef]
- Medrano-Galarza, C.; Gibbons, J.; Wagner, S.; de Passillé, A.M.; Rushen, J. Behavioral changes in dairy cows with mastitis. J. Dairy Sci. 2012, 95, 6994–7002. [Google Scholar] [CrossRef]
- Calderon, D.F.; Cook, N.B. The effect of lameness on the resting behavior and metabolic status of dairy cattle during the transition period in a freestall-housed dairy herd. J. Dairy Sci. 2011, 94, 2883–2894. [Google Scholar] [CrossRef]
- Beer, G.; Alsaaod, M.; Starke, A.; Schuepbach-Regula, G.; Müller, H.; Kohler, P.; Steiner, A. Use of extended characteristic of locomotion and feeding behavior for automated identification of lame dairy cows. PLoS ONE 2016, 11, e0155796. [Google Scholar] [CrossRef]
- Galindo, F.; Broom, D.M. The effects of lameness on social and individual behavior of dairy cows. J. Appl. Anim. Welf. Sci. 2002, 5, 193–201. [Google Scholar] [CrossRef]
- King, M.T.M.; DeVries, T.J. Graduate Student Literature Review: Detecting health disorders using data from automatic milking systems and associated technologies. J. Dairy Sci. 2018, 101, 8605–8614. [Google Scholar] [CrossRef]
- Miguel-Pacheco, G.G.; Kaler, J.; Remnant, J.; Cheyne, L.; Abbott, C.; French, A.P.; Pridmore, T.P.; Huxley, J.N. Behavioural changes in dairy cows with lameness in an automatic milking system. Appl. Anim. Behav. Sci. 2014, 150, 1–8. [Google Scholar] [CrossRef]
- Garcia, E.; Klaas, I.; Amigo, J.M.; Bro, R.; Enevoldsen, C. Lameness detection challenges in automated milking systems addressed with partial least squares discriminant analysis. J. Dairy Sci. 2014, 97, 7476–7486. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.L. Grazing behaviour and helminthic disease. Brit. J. Anim. Behav. 1954, 2, 61–62. [Google Scholar] [CrossRef]
- Burdick, N.C.; Randel, R.D.; Carroll, J.A.; Welsh, T.H., Jr. Interactions between Temperament, Stress, and Immune Function in Cattle. Int. J. Zool. 2011, 2011, 373197. [Google Scholar] [CrossRef]
- Sarkar, A.; Harty, S.; Johnson, K.V.-A.; Moeller, A.H.; Carmody, R.N.; Lehto, S.M.; Erdman, S.E.; Dunbar, R.I.M.; Burnet, P.W.J. The role of microbiome in the neurobiology of social behaviour. Biol. Rev. 2020, 95, 1131–1166. [Google Scholar] [CrossRef]
- Von Holst, D. The concept of stress and its relevance for animal behavior. In Advances in the Study of Behaviour; Moller, A.P., Milinski, M., Slater, P.J.B., Eds.; Academic Press: Cambridge, MA, USA, 1998; Volume 27, pp. 1–131. [Google Scholar]
- Wascher, C.A.F. Heart rate as a measure of emotional arousal in evolutionary biology. Phil. Trans. R. Soc. 2021, 376, 1831. [Google Scholar] [CrossRef]
- Kovaćs, L.; Jurkovich, V.; Bakony, M.; Szenci, O.; Póti, P.; Tözsér, J. Welfare implication of measuring heart rate and heart rate variability in dairy cattle: Literature review and conclusions for future research. Animal 2014, 8, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Hopster, H.; O’Connell, J.M.; Blokhuis, H.J. Acute effects of cow-calf separation on heart rate, plasma cortisol and behaviour in multiparous dairy cows. Appl. Anim. Behav. Sci. 1995, 44, 1–8. [Google Scholar] [CrossRef]
- Laister, S.; Stockinger, B.; Regner, A.-M.; Zenger, K.; Knierim, U.; Winckler, C. Social licking in dairy cattle—Effects on heart rate in performers and receivers. Appl. Anim. Behav. Sci. 2011, 130, 81–90. [Google Scholar] [CrossRef]
- Geburt, K.; Friedrich, M.; Piechotta, M.; Gauly, M.; König von Borstel, U. Validity of physiological biomarkers for maternal behavior in cows—A comparison of beef and dairy cattle. Physiol. Behav. 2015, 139, 361–368. [Google Scholar] [CrossRef]
- Stewart, R.A.; Willis, R.M.; Panuwatwanich, K.; Sahin, O. Showering behavioural response to alarming visual display monitors: Longitudinal mixed method study. Behav. Inform. Technol. 2013, 32, 695–711. [Google Scholar] [CrossRef]
- Rushen, J.; de Passillé, A.M.B.; Munksgaard, L. Fear of People by Cows and Effects on Milk Yield, Behavior, and Heart Rate at Milking. J. Dairy Sci. 1999, 82, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Hagen, K.; Langbein, J.; Schmied, C.; Lexer, D.; Waiblinger, S. Heart rate variability in dairy cows—Influences of breed and milking system. Physiol. Behav. 2005, 85, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Gygax, L.; Neuffer, I.; Kaufmann, C.; Hauser, R.; Wechsler, B. Restlessness behaviour, heart rate and heart-rate variability of dairy cows milked in two types of automatic milking systems and auto-tandem milking parlours. Appl. Anim. Behav. Sci. 2008, 109, 167–179. [Google Scholar] [CrossRef]
- Friedrich, J.; Brand, B.; Schwerin, M. Genetics of cattle temperament and its impact on livestock production and breeding—A review. Arch. Anim. Breed. 2015, 58, 13–21. [Google Scholar] [CrossRef]
- Wenzel, C.; Schönreiter-Fischer, S.; Unshelm, J. Studies on step-kick behavior and stress of cows during milking in an automatic milking system. Livest. Prod. Sci. 2003, 83, 237–246. [Google Scholar] [CrossRef]
- Jorquera-Chavez, M.; Fuentes, S.; Dunshea, F.R.; Warner, R.D.; Poblete, T.; Jongman, E.C. Modelling and validation of computer vision techniques to assess heart rate, eye temperature, ear-base temperature and respiration rate in cattle. Animals 2019, 9, 1089. [Google Scholar] [CrossRef]
- O’Neill, H.A. A review on the involvement of catecholamines in animal behaviour. S. Afr. J. Anim. Sci. 2019, 49, 1–8. [Google Scholar] [CrossRef]
- Chen, Q.; Qu, K.; Ma, Z.; Zhan, J.; Zhang, F.; Shen, J.; Ning, Q.; Jia, P.; Zhang, J.; Chen, N.; et al. Genome-wide association study identifies genomic loci associated with neurotransmitter concentration in cattle. Front. Genet. 2020, 11, 139. [Google Scholar] [CrossRef]
- Friedrich, J.; Brand, B.; Ponsuksili, S.; Graunke, K.L.; Langbein, J.; Knaust, J.; Kühn, C.; Schwerin, M. Detection of genetic variants affecting cattle behaviour and their impact on milk production: A genome-wide association study. Anim. Genet. 2015, 47, 12–18. [Google Scholar] [CrossRef]
- Hopster, H.; Bruckmaier, R.M.; Van der Werf, J.T.N.; Korte, S.M.; Macuhova, J.; Korte-Bouws, G.; van Reenen, C.G. Stress responses during milking; Comparing conventional and automatic milking in primiparous dairy cows. J. Dairy Sci. 2002, 85, 3206–3216. [Google Scholar] [CrossRef]
- Marino, M.D.; Bourdélat-Parks, B.N.; Liles, L.C.; Weinshenker, D. Genetic reduction of noradrenergic function alters social memory and reduces aggression in mice. Behav. Brain Res. 2005, 161, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Garza-Brenner, E.; Sifuentes-Rincón, A.M.; Randel, R.D.; Paredes-Sánchez, F.A.; Parra-Bracamonte, G.M.; Arellano Vera, W.; Rodríguez Almeida, F.A.; Segura Cabrera, A. Association of SNPs in dopamine and serotonin pathway genes and their interacting genes with temperament traits in Charolais cows. J. Appl. Genet. 2016, 58, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Radkowska, I.; Godyn, D.; Fic, K. Stereotypic behaviour in cattle, pigs and horses—A review. Anim. Sci. Papers Rep. 2020, 38, 303–319. [Google Scholar]
- Bacqué-Cazenave, J.; Bharatiya, R.; Barriére, G.; Delbecque, J.-P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdére, P. Serotonin in animal cognition and behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef]
- Marrero, M.G.; Field, S.L.; Skibiel, A.L.; Dado-Senn, B.; Driver, J.P.; Laporta, J. Increasing serotonin bioavailability alters gene expression in peripheral leukocytes and lymphoid tissues of dairy calves. Sci. Rep. 2020, 10, 9712. [Google Scholar] [CrossRef]
- O’Neill, H.A. The Influence of Catecholamines on Energy Metabolism and Selected Meat Quality Attributes of Three Commercial Beef Breeds. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2016. [Google Scholar]
- Nevard, R.P.; Pan, S.D.; Broster, J.C.; Norman, S.T.; Stephen, C.P. Maternal behavior in beef cattle: The physiology, assessment and future directions—A review. Vet. Sci. 2023, 10, 10. [Google Scholar] [CrossRef]
- Cooke, R.F.; Moriel, P.; Cappellozza, B.I.; Miranda, V.F.B.; Batista, L.F.D.; Colombo, E.A.; Ferreira, V.S.M.; Miranda, M.F.; Marques, R.S.; Vasconcelos, J.L.M. Effects of temperament on growth, plasma cortisol concentrations and puberty attainment in Nelore beef heifers. Animal 2019, 13, 1209–1213. [Google Scholar] [CrossRef]
- Knights, M.; Smith, G.W. Decreased ACTH secretion during prolonged transportation stress is associated with reduced pituitary responsiveness to tropic hormone stimulation in cattle. Domest. Anim. Endocrin. 2007, 33, 442–450. [Google Scholar] [CrossRef]
- Zulkifli, I.; Najafi, P.; Nurfarahin, A.J.; Soleimani, A.F.; Kumari, S.; Aryani, A.A.; O’Reilly, E.L.; Eckersall, P.D. Acute phase proteins, interleukin 6, and heat shock protein 70 in broiler chickens administered with corticosterone. Poult. Sci. 2014, 93, 3112–3118. [Google Scholar] [CrossRef]
- Boissy, A.; La Neindre, P. Behavioral, cardiac and cortisol responses to brief peer separation and reunion in cattle. Physiol. Behav. 1997, 61, 693–699. [Google Scholar] [CrossRef]
- Kendrick, K.M. Oxytocin, motherhood and bonding. Exp. Physiol. 2000, 85, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Vandenheede, M.; Bouissou, M.F. Effect of Androgen Treatment on Fear Reactions in Ewes. Horm. Behav. 1993, 27, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Staley, M.; Conners, M.G.; Hall, K.; Miller, L.J. Linking stress and immunity: Immunoglobulin A as a non-invasive physiological biomarker in animal welfare studies. Horm. Behav. 2018, 102, 55–68. [Google Scholar] [CrossRef]
- Brand, B.; Hadlich, F.; Brandt, B.; Schauer, N.; Graunke, K.L.; Langbein, J.; Repsilber, D.; Ponsuksili, S.; Schwerin, M. Temperament type specific metabolite profiles of the prefrontal cortex and serum in cattle. PLoS ONE 2015, 10, e0125044. [Google Scholar] [CrossRef] [PubMed]
- Akimoto, H.; Oshima, S.; Sugiyama, T.; Negishi, A.; Nemoto, T.; Kobayashi, D. Changes in brain metabolites related to stress resilience: Metabolomic analysis of the hippocampus in a rat model of depression. Behav. Brain Res. 2019, 359, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.M.; Mancano, G.; Kashofer, K.; Fröhlich, E.E.; Matak, A.; Mayerhofer, R.; Reichmann, F.; Olivares, M.; Neyrinck, A.M.; Delzenne, N.M.; et al. High-fat diet induces depression-like behaviour in mice associated with changes in microbiome, neuropeptide Y, and brain metabolome. Nutr. Neurosci. 2019, 22, 877–893. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Bai, J.; Du, J.; Wu, X.; Thomsen, B.; Gao, H.; Su, G.; Wang, X. Overview of metabolomic analysis and the integration with multi-omics for economic traits in cattle. Metabolites 2021, 11, 753. [Google Scholar] [CrossRef]
- Serova, L.I.; Gueorguiev, V.; Cheng, S.-Y.; Sbban, E.L. ACTH elevates gene expression for catecholamine biosynthesis in rat superior cervical ganglia and locus coeruleus by an adrenal independent mechanism. Neuroscience 2008, 153, 1380–1389. [Google Scholar] [CrossRef]
- Li, Y.H.; Li, F.; Liu, M.; Yin, J.J.; Cheng, B.J.; Shi, B.M.; Shan, A.S. Effect of γ-aminobutyric acid on growth performance, behavior and plasma hormones in weaned pigs. Can. J. Anim. Sci. 2015, 95, 165–171. [Google Scholar] [CrossRef]
- Wang, D.M.; Wang, C.; Li, H.Y.; Liu, J.X.; Ferguson, J.D. Effects of rumen-protected γ-aminobutyric acid on feed intake, lactation performance, and antioxidative status in early lactating dairy cows. J. Dairy Sci. 2013, 96, 3222–3227. [Google Scholar] [CrossRef]
- Brand, W.; Wells, A.T.; Smith, S.L.; Denholm, S.J.; Wall, E.; Coffey, M.P. Predicting pregnancy status from mid-infrared spectroscopy in dairy cow milk using deep learning. J. Dairy Sci. 2021, 104, 4980–4990. [Google Scholar] [CrossRef] [PubMed]
- Denholm, S.J.; Brand, W.; Mitchell, A.P.; Wells, A.T.; Kryzelewski, T.; Smith, S.L. Predicting bovine tuberculosis status of dairy cows from mid-infrared spectral data using deep learning. J. Dairy Sci. 2020, 103, 9355–9367. [Google Scholar] [CrossRef] [PubMed]
- Benfica, L.F.; Sakamoto, L.S.; Magalhães, A.F.B.; de Oliviera, M.H.V.; de Albuquerque, L.G.; Cavalheiro, R.; Branco, R.H.; Cyrillo, J.N.S.G.; Mercadante, M.E.Z. Genetic association among feeding behavior, feed efficiency, and growth traits in growing indicine cattle. J. Anim. Sci. 2020, 98, skaa350. [Google Scholar] [CrossRef]
- Nkrumah, J.D.; Crews, D.H., Jr.; Basarab, J.A.; Price, M.A.; Okine, E.K.; Wang, Z.; Li, C.; Moore, S.S. Genetic and phenotypic relationship of feeding behavior and temperament with performance, feed efficiency, ultrasound, and carcass merit of beef cattle. J. Anim. Sci. 2007, 85, 2382–2390. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.L.; Oddy, V.H. Genetic parameters for feed efficiency, fatness, muscle area and feeding behaviour of feedlot finished beef cattle. Livest. Prod. Sci. 2004, 90, 255–270. [Google Scholar] [CrossRef]
- Lin, Z.; Macloed, I.; Pryce, J.E. Short communication: Estimation of genetic parameters for residual feed intake and feeding behavior traits in dairy heifers. J. Dairy Sci. 2013, 96, 2654–2656. [Google Scholar] [CrossRef]
- Yin, T.; Jaeger, M.; Scheper, C.; Grodkowski, G.; Sakowski, T.; Klopčič, M.; Bapst, B.; König, S. Multi-breed genome-wide association studies across countries for electronically recorded behavior traits in local dual-purpose cows. PLoS ONE 2019, 14, e0221973. [Google Scholar] [CrossRef]
- Dickson, D.P.; Barr, G.R.; Johnson, L.P.; Wieckert, D.A. Social Dominance and Temperament of Holstein Cows. J. Dairy Sci. 1970, 53, 904–907. [Google Scholar] [CrossRef]
- Kramer, M.; Erbe, M.; Seefried, F.R.; Gredler, B.; Bapst, B.; Bieber, A.; Simianer, H. Accuracy of direct genomic values for functional traits in Brown Swiss cattle. J. Dairy Sci. 2014, 97, 1774–1781. [Google Scholar] [CrossRef]
- Jakimowicz, M.; Szyda, J.; Zarnecki, A.; Jagusiak, W.; Morek-Kopeć, M.; Kosińska-Selbi, B.; Suchocki, T. Genome-Wide Genomic and Functional Association Study for Workability and Calving Traits in Holstein Cattle. Animals 2022, 12, 1127. [Google Scholar] [CrossRef]
- Marete, A.; Sahana, G.; Fritz, S.; Lefebvre, R.; Barbat, A.; Lund, M.S.; Guldbrandtsen, B.; Boichard, D. Genome-wide association study for milking speed in French Holstein cows. J. Dairy Sci. 2018, 101, 6205–6219. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.V.; Brügemann, K.; Ebinghaus, A.; König, S. Genetic parameters for longitudinal behavior and health indicator traits generated in automatic milking systems. Arch. Anim. Breed. 2018, 61, 161–171. [Google Scholar] [CrossRef]
- Kramer, M.; Erbe, M.; Bapst, B.; Bieber, A.; Simianer, H. Estimation of genetic parameters for novel functional traits in Brown Swiss cattle. J. Dairy Sci. 2013, 96, 5954–5964. [Google Scholar] [CrossRef]
- Le Neindre, P.; Grignard, L.; Trillat, G.; Boissy, A.; Ménissier, F.; Sapa, F.; Boivin, X. Docile Limousine cows are not poor mothers. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002. [Google Scholar]
- Carlström, C.; Pettersson, G.; Johansson, K.; Strandberg, E.; Stålhammar, H.; Philipsson, J. Feasibility of using automatic milking system data from commercial herds for genetic analysis of milkability. J. Dairy Sci. 2013, 96, 5324–5332. [Google Scholar] [CrossRef] [PubMed]
- König, S.; Köhn, F.; Kuwan, K.; Simianer, H.; Gauly, M. Use of repeated measures analysis for evaluation of genetic background of dairy cattle behavior in automatic milking systems. J. Dairy Sci. 2006, 89, 3636–3644. [Google Scholar] [CrossRef]
- Nixon, M.; Bohmanova, J.; Jamrozik, J.; Schaeffer, L.R.; Hand, K.; Miglior, F. Genetic parameters of milking frequency and milk production traits in Canadian Holsteins milked by an automated milking system. J. Dairy Sci. 2009, 92, 3422–3430. [Google Scholar] [CrossRef]
- Enghardt, T.; Swalve, H.H. Genetisch-statistische Untersuchungen zum Verhalten von Milchkühen unter AMS-Bedingungen. In Proceedings of the Tagung der Deutschen Gesellschaft für Züchtungskunde (DGfZ) und der Gesellschaft für Tierzuchtwissenschaften (GfT), Giessen, Germany, 11–12 September 2019. [Google Scholar]
- Carlström, C.; Strandberg, E.; Petterson, G.; Johansson, K.; Stalhammar, H.; Philipsson, J. Genetic associations of teat cup attachment failures, incomplete milkings, and handling time in automatic milking systems with milkability, temperament, and udder conformation. Acta Agric. Scand. Sect. A Anim. Sci. 2016, 66, 75–83. [Google Scholar] [CrossRef]
- Stephansen, R.S.; Fogh, A.; Norberg, E. Genetic parameters for handling and milking temperament in Danish first-parity Holstein cows. J. Dairy Sci. 2018, 101, 11033–11039. [Google Scholar] [CrossRef]
- Vosman, J.J.; de Jon, G.; Eding, H. Breeding of cows suitable for an automatic milking system. In Proceedings of the INTERBULL BULLETIN No. 48, Berlin, Germany, 20–21 May 2014. [Google Scholar]
- Bakke, K.A.; Heringstad, B. Data from automatic milking systems used in genetic evaluations of temperament and milkability. In Proceedings of the 66th Annual Meeting of the European Association for Animal Production, Warsaw, Poland, 31 August–4 September 2015. [Google Scholar]
- Løvendahl, P.; Lassen, J.; Chagunda, M.G.G. Milking efficiency—A milkability trait for automatically milked cowds. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014. [Google Scholar]
- Vosman, J.J.; Poppe, H.W.M.; Mulder, H.A.; Assen, A.; Ducro, B.J.; Gerrits, A.; Vesseur, C.; Boes, J.H.; Eding, H.; de Jong, G. Automatic milking system, a source for novel phenotypes as base for new genetic selection tools. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018. [Google Scholar]
- Løvendahl, P.; Buitenhuis, A.J. Genetic and phenotypic variation and consistency in cow preference and circadian use of robotic milking units. J. Dairy Sci. 2022, 105, 5283–5295. [Google Scholar] [CrossRef]
- Rinell, E. A Genetic Analysis of Traits Recorded by Automatic Milking Systems—The Possibility for a New Method to Evaluate Temperament of Dairy Cows. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2013. [Google Scholar]
- Gäde, S.; Stamer, E.; Junge, W.; Kalm, E. Estimates of genetic parameters for milkability from automatic milking. Livest. Sci. 2006, 104, 135–146. [Google Scholar] [CrossRef]
- Aerts, J.; Piwczyński, D.; Ghiasi, H.; Sitkowska, B.; Kolenda, M.; Önder, H. Genetic parameters estimation of milking traits in Polish Holstein-Friesians based on automatic milking system data. Animals 2021, 11, 1943. [Google Scholar] [CrossRef] [PubMed]
- CattleQTLdb. Available online: https://www.animalgenome.org/cgi-bin/QTLdb/BT/index (accessed on 11 September 2023).
- Kelly, A.K.; McGee, M.; Crews, D.H., Jr.; Fahey, A.G.; Wylie, A.R.; Kenny, D.A. Effect of divergence in residual feed intake on feeding behavior, blood metabolic variables, and body composition traits in growing beef heifers. J. Anim. Sci. 2010, 88, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fang, L.; Null, D.J.; Hutchison, J.L.; Connor, E.E.; VanRaden, P.M.; VandeHaar, M.J.; Tempelman, R.J.; Weigel, K.A.; Cole, J.B. High-density genome-wide association study for residual feed intake in Holstein dairy cattle. J. Dairy Sci. 2019, 102, 11067–11080. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gondro, C.; Quinn, K.; Herd, R.M.; Parnell, P.F.; Vanselow, B. Global gene expression profiling reveals genes expresses differentially in cattle with high and low residual feed intake. Anim. Genet. 2011, 42, 475–490. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Kuehn, L.A.; Oliver, W.T.; Kern, R.J.; Cushman, R.A.; Miles, J.R.; McNeel, A.K.; Freetly, H.C. DNA polymorphisms and transcript abundance of PRKAG2 and phosphorylated AMP-activated protein kinase in the rumen are associated with gain and feed intake in beef steers. Anim. Genet. 2014, 45, 461–472. [Google Scholar] [CrossRef]
- Doherty, A.; Smith-Byrne, K.; Ferreira, T.; Holmes, M.V.; Holmes, C.; Pulit, S.L.; Lindgren, C.M. GWAS identifies 14 loci for device-measured physical activity and sleep duration. Nat. Commun. 2018, 9, 5257. [Google Scholar] [CrossRef]
- Kommadath, A.; Woelders, H.; Beerda, B.; Mulder, H.A.; de Wit, A.A.C.; Veerkamp, R.F.; te Pas, M.F.W.; Smits, M.A. Gene expression patterns in four brain areas associate with quantitative measure of estrous behavior in dairy cows. Genomics 2011, 12, 200. [Google Scholar] [CrossRef]
- Woelders, H.; van der Lende, T.; Kommadath, A.; te Pas, M.F.W.; Smits, M.A.; Kaal, L.M.T.E. Central genomic regulation of the expression of the oestrus behaviour in dairy cows: A review. Animal 2014, 8, 754–764. [Google Scholar] [CrossRef]
- Imran, S.; Maryam, J.; Nadeem, A.; Iqbal, M. Pretentious genomic selection signatures in CYP19A1 gene associated with silent estrous behaviour in water buffalo in Pakistan. Electron. J. Biotechnol. 2018, 32, 35–40. [Google Scholar] [CrossRef]
- Guillemin, N.; Dufort, I.; Sirard, M.-A. A genetical genomics methodology to identify genetic markers of a bovine fertility phenotype based on CYP19A1 gene expression. Gene Expr. Genet. Genom. 2015, 8, 9–18. [Google Scholar]
- Abo-Ismail, M.K.; Miller, S.P.; Sargolzaei, M.; Grossi, D.A.; Nayeri, S.; Moore, S.S.; Plastow, G.; Stothard, P.; Schenkel, F. Genome wide association analyses identify new loci for milking speed and temperament in North American Holsteins. In Proceedings of the 10th World Congress of Genetics Applied to Livestock Production, Volume Genetics of Trait Complexes: Lactation, Vancouer, BC, Canada, 17–22 August 2014. [Google Scholar]
- Kolbehdari, D.; Wang, Z.; Grant, J.R.; Murdoch, B.; Prasad, A.; Xiu, Z.; Marques, E.; Stothard, P.; Moore, S.S. A whole-genome scan to map quantitative trait loci for conformation and functional traits in Canadian Holstein bulls. J. Dairy Sci. 2008, 91, 2844–2856. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Oliveaira, H.R.; Schenkel, F.S.; Pedrosa, V.B.; Melka, M.G.; Brito, L.F. Using whole-genome sequence variants to uncover candidate mutations and genes affecting milking speed and temperament in Holstein cattle. J. Dairy Sci. 2020, 103, 10383–10398. [Google Scholar] [CrossRef] [PubMed]
- Ensembl Genome Browser. Available online: https://www.ensembl.org (accessed on 6 September 2023).
- Gutiérrez-Gil, B.; Ball, N.; Burton, D.; Haskell, M.; Williams, J.L.; Wiener, P. Identification of quantitative trait loci affecting cattle temperament. J. Hered. 2008, 99, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Glenske, K.; Brandt, H.; Gauly, M.; Erhardt, G. Analysis of association of two SNP in cholecystokinin B receptor gene with behaviour scores in German Angus and German Simmental cattle (Brief report). Archiv Tierzucht. 2010, 53, 494–496. [Google Scholar] [CrossRef]
- Glenske, K.; Prinzenberg, E.-M.; Brandt, H.; Gauly, M.; Erhardt, G. A chromosome-wide QTL study on BTA29 affecting temperament traits in German Angus beef cattle and mapping of DRD4. Animal 2011, 5, 195–197. [Google Scholar] [CrossRef]
- Paredes-Sánchez, F.A.; Sifuentes-Rincón, A.M.; Casas, E.; Arellano-Vera, W.; Parra-Bracamonte, G.M.; Riley, D.G.; Welsh, T.H., Jr.; Randel, R.D. Novel genes involved in the genetic architecture of temperament in Brahman cattle. PLoS ONE 2020, 15, e0237825. [Google Scholar] [CrossRef]
- Urwin, R.E.; Bennetts, B.H.; Wilcken, B.; Lampropoulos, B.; Beumont, P.J.V.; Russell, J.D.; Tanner, S.L.; Nunn, K.P. Gene-gene interaction between the monoamine oxidase A gene and solute carrier family 6 (neurotransmitter transporter, noradrenalin) member 2 gene in anorexia nervosa (restrictive subtype). Eur. J. Hum. Genet. 2003, 11, 945–950. [Google Scholar] [CrossRef]
- Michenet, A.; Saintilan, R.; Venot, E.; Phocas, F. Insights into the genetic variation of maternal behaviour and suckling performance of continental beef cows. Genet. Sel. Evol. 2016, 48, 45. [Google Scholar] [CrossRef]
- Schafberg, R.; Rosner, F.; Oelschlägel, D.; Swalve, H.H. Learning behaviour of dairy cows in automated milking systems: Genetic parameters and suggested candidate genes. In Proceedings of the 12th World Congress on Genetics Applied to Livestock Production, Rotterdam, The Netherlands, 3–8 July 2022. [Google Scholar]
- Corbetta, S.; Gualdoni, S.; Albertinazzi, C.; Paris, S.; Croci, L.; Consalez, G.G.; de Curtis, I. Generation and characterization of Rac3 knockout mice. Mol. Cell. Biol. 2005, 25, 5763–5776. [Google Scholar] [CrossRef]
- Al-Kanaan, A.J.J. Heat Stress Response for Physiological Traits in Dairy and Dual-Purpose Cattle Populations on Phenotypic and Genetic Scales. Ph.D. Thesis, University of Kassel, Kassel, Germany, 2016. [Google Scholar]
- Luo, H.; Hu, L.; Brito, L.F.; Dou, J.; Sammad, A.; Chang, Y.; Ma, L.; Guo, G.; Liu, L.; Zhai, L.; et al. Weighted single-step GWAS and RNA sequencing reveals key candidate genes associated with physiological indicators of heat stress on Holstein cattle. J. Anim. Sci. Biotechnol. 2022, 13, 108. [Google Scholar] [CrossRef]
- Shen, J.-F.; Chen, Q.-M.; Zhang, F.-Z.; Hanif, Q.; Huang, B.-Z.; Chen, N.-B.; Qu, K.-X.; Zhan, J.-X.; Chen, H.; Jiang, Y.; et al. Genome-wide association study identifies quantitative trait loci affecting cattle temperament. Zool. Res. 2022, 43, 14–25. [Google Scholar] [CrossRef]
- Marsh, D.J.; Weingarth, D.T.; Novi, D.E.; Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Guan, X.M.; Jiang, M.M.; Feng, Y.; Camacho, R.E.; et al. Melanin-concentrating hormone 1 receptor-deficient mice are lean, hyperactive, and hyperphagic and have altered metabolism. Proc. Natl. Acad. Sci. USA 2002, 99, 3240–3245. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; David, N.K.; Danao, J.V.; Baribault, H.; Tian, H.; Giorgetti, M. Genetic Inactivation of Melanin-Concentrating Hormone Receptor Subtype 1 (MCHR1) in Mice Exerts Anxiolytic-Like Behavioral Effects. Neuropsychopharmacology 2006, 31, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Strekalova, T.; Svirin, E.; Waider, J.; Gorlova, A.; Cespuglio, R.; Kaleff, A.; Pomytkin, I.; Schmitt-Boehrer, A.G.; Lesch, K.P.; Anthony, D.C. Altered behaviour, dopamine and norepinephrine regulation in stressed mice heterozygous in TPH2 gene. Prog. Neuro-Psychopharm. 2021, 108, 110155. [Google Scholar] [CrossRef] [PubMed]
- Kopečková, M.; Paclt, I.; Goetz, P. Polymorphisms of Dopamine-ß-Hydroxylase in ADHD Children. Fol. Biol. 2006, 52, 194–201. [Google Scholar]
- Polasik, D.; Konieczna, A.; Terman, A.; Dybus, A. The association of C789A polymorphism in the dopamine beta-hydroxylase gene (DBH) and aggressive behaviour in dogs. Acta Vet. Brno 2021, 90, 295–299. [Google Scholar] [CrossRef]
- Lourenco-Jaramillo, D.L.; Sifuentes-Rincón, A.M.; Parra-Bracamonte, G.M.; de la Rosa-Reyna, X.F.; Segura-Cabrera, A.; Arellano-Vera, W. Genetic diversity of tyrosine hydroxylase (TH) and dopamine ß-hydroxylase (DBH) genes in cattle breeds. Genet. Mol. Biol. 2012, 35, 435–440. [Google Scholar] [CrossRef]
- Sifuentes-Rincón, A.M.; Trejo-Tapia, A.G.; Randel, R.D.; Abriz-Morales, P.; Parra-Bracamonte, G.N. Bovine dopamine receptors DRD1, DRD4 and DRD5: Genetic polymorphisms and diversities among ten cattle breeds. Genet. Mol. Res. 2016, 15, 12017725. [Google Scholar] [CrossRef]
- Fleming, A.S.; Kraemer, G.W. Molecular and Genetic Bases of Mammalian Maternal Behavior. Gend. Genome 2019, 3, 1–14. [Google Scholar] [CrossRef]
- Wang, L.P. mRNA expression of prolactin receptor in sheep pituitary and its effect on maternal behavior. Genet. Mol. Res. 2015, 14, 8650–8657. [Google Scholar] [CrossRef]
- Moreno García, C.A.; Zhou, H.; Altimira, D.; Dynes, R.; Gregorini, P.; Jayathunga, S.; Maxwll, T.N.R.; Hickford, J. The glutamate metabotropic receptor 5 (GRM5) gene is associated with beef cattle home range and movement tortuosity. J. Anim. Sci. Biotechnol. 2022, 13, 111. [Google Scholar] [CrossRef] [PubMed]
- Eusebi, P.G.; Cortés, O.; Carleos, C.; Dunner, S.; Cañon, J. Detection of selection signatures for agonistic behaviour in cattle. J. Anim. Breed. Genet. 2018, 135, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.S.; Suh, G.H.; Kwon, E.G. Estimation of Environmental Effects and Genetic Parameters for Somatic Cell Score, Stress and Immunological Traits in Holstein Cattle. J. Anim. Sci. Technol. 2006, 48, 9–14. [Google Scholar]
- Cockrum, R.R.; Speidel, S.E.; Salak-Johnson, J.L.; Chase, C.C.L.; Peel, R.K.; Weaber, R.L.; Loneagan, G.H.; Wagner, J.J.; Boddhireddy, P.; Thomas, M.G.; et al. Genetic parameters estimated at receiving for circulating cortisol, immunoglobulin G, interleukin 8, and incidence of bovine respiratory disease in feedlot beef steers. J. Anim. Sci. 2016, 94, 2770–2778. [Google Scholar] [CrossRef]
- Liu, S.; Vallejo, R.L.; Gao, G.; Palti, Y.; Weber, G.M.; Herandez, A.; Rexroad, C.E., III. Identification of single-nucleotide polymorphism markers associated with cortisol response to crowding in rainbow trout. Mar. Biotechnol. 2015, 17, 328–337. [Google Scholar] [CrossRef]
- Neumann, A.; Direk, N.; Crawford, A.A.; Mirza, S.; Adams, H.; Bolton, J.; Hayward, C.; Strachan, D.P.; Payne, E.K.; Smith, J.A.; et al. The low single nucleotide polymorphism heritability of plasma and saliva cortisol levels. Psychoneuroendocrinology 2017, 85, 88–95. [Google Scholar] [CrossRef]
- Byrne, T.J.; Santos, B.F.S.; Amer, P.R.; Martin-Collado, D.; Pryce, J.E.; Axford, M. New breeding objectives and selection indices for the Australian dairy industry. J. Dairy Sci. 2016, 99, 8146–8167. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, F.C.; Peixoto, M.G.C.D.; Fonseca, P.A.d.S.; Pires, M.d.F.A.; Ventura, R.V.; Rosse, I.d.C.; Bruneli, F.A.T.; Machado, M.A.; Carvalho, M.R.S. Identification of candidate genes for reactivity in Guzerat (Bos indicus) Cattle: A genome-wide association study. PLoS ONE 2017, 12, e0169163. [Google Scholar] [CrossRef]
- Rancelis, T.; Domarkiene, I.; Ambrozaityte, L.; Utkus, A. Implementing core genes and an omnigenic model for behaviour traits prediction in genomics. Genes 2023, 14, 1630. [Google Scholar] [CrossRef]
- González-Recio, O.; Martínez-Álvaro, M.; Tiezzi, F.; Saborío-Montero, A.; Maltecca, C.; Roehe, R. Invited review: Novel methods and perspectives for modulating the rumen microbiome through selective breeding as a means to improve complex traits: Implications for methane emissions in cattle. Livest. Sci. 2023, 269, 105171. [Google Scholar] [CrossRef]
- Neumann, G.B.; Korkuć, P.; Arends, D.; Wolf, M.J.; May, K.; König, S.; Brockmann, G.A. Genomic diversity and relationship analyses of endangered German Black Pied cattle (DSN) to 68 other taurine breeds based on whole-genome sequencing. Front. Genet. 2023, 13, 993959. [Google Scholar] [CrossRef] [PubMed]
- Mathis, M.W.; Mathis, A. Deep learning tools for the measurement of animal behaviour in neuroscience. Curr. Opin. Neurobiol. 2020, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.M.; Warner, R.D. Have we underestimated the impact of pre-slaughter stress on meat quality in ruminants? Meat Sci. 2008, 80, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Van der Laak, M.; van Pelt, M.L.; de Jong, G.; Mulder, H.A. Genotype by environment interaction for production, somatic cell score, workability, and conformation traits in Dutch Holstein-Friesian cows between farms with or without grazing. J. Dairy Sci. 2016, 99, 4496–4503. [Google Scholar] [CrossRef] [PubMed]
- Ketelaar-de Lauwere, C.C.; Ipema, A.H.; van Ouwerkerk, E.N.J.; Hendriks, M.M.W.B.; Metz, J.H.M.; Noordhuizen, J.P.T.M.; Schouten, W.G.P. Voluntary automatic milking in combination with grazing of dairy cows Milking frequency and effects on behaviour. Appl. Anim. Behav. Sci. 1999, 64, 91–109. [Google Scholar] [CrossRef]
- Guan, J.-S.; Xie, H.; Ding, X. The role of epigenetic regulation in learning and memory. Exp. Neurol. 2015, 268, 30–36. [Google Scholar] [CrossRef]
- Dias, B.G.; Maddox, S.A.; Klengel, T.; Ressler, K.J. Epigenetic mechanisms underlying learning and the inheritance of learned behaviors. Trends Neurosci. 2015, 38, 2. [Google Scholar] [CrossRef]
- Weigel, K.A.; VanRaden, P.M.; Norman, H.D.; Grosu, H. A 100-Year Review: Methods and impact of genetic selection in dairy cattle—From daughter-dam comparisons to deep learning algorithms. J. Dairy Sci. 2017, 100, 10234–10250. [Google Scholar] [CrossRef]
- Yang, Y.-I.; Zhou, R.; Li, K. Future livestock breeding: Precision breeding based on multi-omics information and population personalization. J. Integr. Agric. 2017, 16, 2784–2791. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behren, L.E.; König, S.; May, K. Genomic Selection for Dairy Cattle Behaviour Considering Novel Traits in a Changing Technical Production Environment. Genes 2023, 14, 1933. https://doi.org/10.3390/genes14101933
Behren LE, König S, May K. Genomic Selection for Dairy Cattle Behaviour Considering Novel Traits in a Changing Technical Production Environment. Genes. 2023; 14(10):1933. https://doi.org/10.3390/genes14101933
Chicago/Turabian StyleBehren, Larissa Elisabeth, Sven König, and Katharina May. 2023. "Genomic Selection for Dairy Cattle Behaviour Considering Novel Traits in a Changing Technical Production Environment" Genes 14, no. 10: 1933. https://doi.org/10.3390/genes14101933
APA StyleBehren, L. E., König, S., & May, K. (2023). Genomic Selection for Dairy Cattle Behaviour Considering Novel Traits in a Changing Technical Production Environment. Genes, 14(10), 1933. https://doi.org/10.3390/genes14101933