
FAAH rs324420 Polymorphism: Biological Pathways, Impact on Elite Athletic Performance and Insights for Sport Medicine
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. The Endocannabinoid System and the Biological Pathways of FAAH rs324420
2.1. The Endocannabinoid System Signalling
2.2. FAAH Protein Functioning and FAAH Gene Variation
2.3. FAAH rs324420 Polymorphism
3. Geographic Distribution of FAAH rs324420 Alleles
4. FAAH rs324420 Polymorphism and Elite Athletic Performance
4.1. Biological Evidence in Elite Athletic Performance
4.2. Biological Pathways, Elite Athletic Performance and Sport Medicine
5. FAAH rs324420 Polymorphism and Other Psychobiological Associations
5.1. FAAH rs324420 Polymorphism, Stress, Anxiety and Fear Extinction
5.2. FAAH rs324420 Polymorphism, Pain and Inflammation
5.3. FAAH rs324420 Polymorphism and Neural Dysfunctions
6. Implications for Sport Medicine
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, H.H.; Silva, M.G.; Cerqueira, F.; Tavares, V.; Medeiros, R. Genomic profile in association with sport-type, sex, ethnicity, psychological traits and sports injuries of elite athletes. J. Sports Med. Phys. Fit. 2022, 62, 418–434. [Google Scholar] [CrossRef] [PubMed]
- Peplonska, B.; Safranow, K.; Adamczyk, J.; Boguszewski, D.; Szymański, K.; Soltyszewski, I.; Barczak, A.; Siewierski, M.; Ploski, R.; Sozanski, H.; et al. Association of serotoninergic pathway gene variants with elite athletic status in the Polish population. J. Sports Sci. 2019, 37, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.-R.G.; Paiva, T.; Silva, H.-H. The elite athlete as a special risk traveller and the jet lag’s effect: Lessons learned from the past and how to be prepared for the next Olympic Games 2020 Tokyo? J. Sports Med. Phys. Fit. 2019, 59, 1420–1429. [Google Scholar] [CrossRef]
- Silva, M.G.; Silva, H.H. Comparison of body composition and nutrients’ deficiencies between Portuguese rink-hockey players. Eur. J. Pediatr. 2017, 176, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.H.; Tavares, V.; Silva, M.-R.G.; Neto, B.V.; Cerqueira, F.; Medeiros, R. FAAH rs324420 Polymorphism Is Associated with Performance in Elite Rink-Hockey Players. Biology 2022, 11, 1076. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.H.; Tavares, V.; Silva, M.-R.G.; Neto, B.V.; Cerqueira, F.; Medeiros, R. Association of FAAH rs324420 (C385A) Polymorphism with High-Level Performance in Volleyball Players. Genes 2023, 14, 1164. [Google Scholar] [CrossRef]
- Moscatelli, F.; Messina, G.; Valenzano, A.; Triggiani, A.I.; Sessa, F.; Carotenuto, M.; Tartaglia, N.; Ambrosi, A.; Cibelli, G.; Monda, V. Effects of twelve weeks’ aerobic training on motor cortex excitability. J. Sports Med. Phys. Fit. 2020, 60, 1383–1389. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Egorova, E.S.; Gabdrakhmanova, L.J.; Fedotovskaya, O.N. Genes and athletic performance: An Update. Med. Sport Sci. 2016, 61, 41–54. [Google Scholar] [CrossRef]
- Petito, A.; Altamura, M.; Iuso, S.; Padalino, F.A.; Sessa, F.; D’Andrea, G.; Margaglione, M.; Bellomo, A. The Relationship between Personality Traits, the 5HTT Polymorphisms, and the Occurrence of Anxiety and Depressive Symptoms in Elite Athletes. PLoS ONE 2016, 11, e0156601. [Google Scholar] [CrossRef]
- Kolla, N.J.; Mizrahi, R.; Karas, K.; Wang, C.; Bagby, R.M.; McMain, S.; Simpson, A.I.; Rusjan, P.M.; Tyndale, R.; Houle, S.; et al. Elevated fatty acid amide hydrolase in the prefrontal cortex of borderline personality disorder: A [11C]CURB positron emission tomography study. Neuropsychopharmacology 2020, 45, 1834–1841. [Google Scholar] [CrossRef]
- Best, L.M.; Wardell, J.D.; Tyndale, R.F.; McPhee, M.D.; Le Foll, B.; Kish, S.J.; Boileau, I.; Hendershot, C.S. Association of the Fatty Acid Amide Hydrolase C385A Polymorphism With Alcohol Use Severity and Coping Motives in Heavy-Drinking Youth. Alcohol. Clin. Exp. Res. 2021, 45, 507–517. [Google Scholar] [CrossRef]
- Hindocha, C.; Quattrone, D.; Freeman, T.P.; Murray, R.M.; Mondelli, V.; Breen, G.; Curtis, C.; Morgan, C.J.A.; Valerie Curran, H.; Di Forti, M. Do AKT1, COMT and FAAH influence reports of acute cannabis intoxication experiences in patients with first episode psychosis, controls and young adult cannabis users? Transl. Psychiatry 2020, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Storr, M.; Emmerdinger, D.; Diegelmann, J.; Yüce, B.; Pfennig, S.; Ochsenkühn, T.; Göke, B.; Lohse, P.; Brand, S. The role of fatty acid hydrolase gene variants in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2009, 29, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Peplonska, B.; Adamczyk, J.G.; Siewierski, M.; Safranow, K.; Maruszak, A.; Sozanski, H.; Gajewski, A.K.; Zekanowski, C. Genetic variants associated with physical and mental characteristics of the elite athletes in the Polish population. Scand. J. Med. Sci. Sports 2017, 27, 788–800. [Google Scholar] [CrossRef]
- Tejera, P.; Meyer, N.J.; Chen, F.; Feng, R.; Zhao, Y.; O’Mahony, D.S.; Li, L.; Sheu, C.C.; Zhai, R.; Wang, Z.; et al. Distinct and replicable genetic risk factors for acute respiratory distress syndrome of pulmonary or extrapulmonary origin. J. Med. Genet. 2012, 49, 671–680. [Google Scholar] [CrossRef]
- Best, L.M.; Hendershot, C.S.; Buckman, J.F.; Jagasar, S.; McPhee, M.D.; Muzumdar, N.; Tyndale, R.F.; Houle, S.; Logan, R.; Sanches, M.; et al. Association Between Fatty Acid Amide Hydrolase and Alcohol Response Phenotypes: A Positron Emission Tomography Imaging Study With [11C]CURB in Heavy-Drinking Youth. Biol. Psychiatry 2023, 94, 405–415. [Google Scholar] [CrossRef]
- Alsaafin, A.; Chenoweth, M.J.; Sylvestre, M.P.; O’Loughlin, J.; Tyndale, R.F. Genetic variation in fatty acid amide hydrolase (FAAH): Associations with early drinking and smoking behaviors. Addict. Behav. 2023, 137, 107545. [Google Scholar] [CrossRef]
- Legaki, E.; Tsaklakidou, D.; Hatzimanolis, A.; Segredou, E.; Petalotis, M.; Moularogiorgou, G.; Mouchtouri, V.; Lykouras, L.; Stefanis, N.C.; Gazouli, M. Association of Alcohol Use Disorder Risk with ADH1B, DRD2, FAAH, SLC39A8, GCKR, and PDYN Genetic Polymorphisms. In Vivo 2022, 36, 2092–2104. [Google Scholar] [CrossRef]
- Best, L.M.; Williams, B.; Le Foll, B.; Mansouri, E.; Bazinet, R.P.; Lin, L.; De Luca, V.; Lagzdins, D.; Rusjan, P.; Tyndale, R.F.; et al. Lower brain fatty acid amide hydrolase in treatment-seeking patients with alcohol use disorder: A positron emission tomography study with [C-11]CURB. Neuropsychopharmacology 2020, 45, 1289–1296. [Google Scholar] [CrossRef]
- Sloan, M.E.; Gowin, J.L.; Yan, J.; Schwandt, M.L.; Spagnolo, P.A.; Sun, H.; Hodgkinson, C.A.; Goldman, D.; Ramchandani, V.A. Severity of alcohol dependence is associated with the fatty acid amide hydrolase Pro129Thr missense variant. Addict. Biol. 2018, 23, 474–484. [Google Scholar] [CrossRef]
- Bühler, K.M.; Huertas, E.; Echeverry-Alzate, V.; Giné, E.; Moltó, E.; Montoliu, L.; López-Moreno, J.A. Risky alcohol consumption in young people is associated with the fatty acid amide hydrolase gene polymorphism C385A and affective rating of drug pictures. Mol. Genet. Genom. 2014, 289, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Spohrs, J.; Ulrich, M.; Grön, G.; Plener, P.L.; Abler, B. FAAH polymorphism (rs324420) modulates extinction recall in healthy humans: An fMRI study. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Lazary, J.; Eszlari, N.; Juhasz, G.; Bagdy, G. Genetically reduced FAAH activity may be a risk for the development of anxiety and depression in persons with repetitive childhood trauma. Neuropsychopharmacology 2016, 26, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Demers, C.H.; Drabant Conley, E.; Bogdan, R.; Hariri, A.R. Interactions Between Anandamide and Corticotropin-Releasing Factor Signaling Modulate Human Amygdala Function and Risk for Anxiety Disorders: An Imaging Genetics Strategy for Modeling Molecular Interactions. Biol. Psychiatry 2016, 80, 356–362. [Google Scholar] [CrossRef]
- Boileau, I.; Tyndale, R.F.; Williams, B.; Mansouri, E.; Westwood, D.J.; Le Foll, B.; Rusjan, P.M.; Mizrahi, R.; De Luca, V.; Zhou, Q.; et al. The fatty acid amide hydrolase C385A variant affects brain binding of the positron emission tomography tracer [11C]CURB. J. Cereb. Blood Flow Metab. 2015, 35, 1237–1240. [Google Scholar] [CrossRef]
- Anvar, L.H.; Alejafar, A.; Moosavi, S.E.; Charsouei, S.; Zeynalzadeh, N.; Fanid, L.M.; Emamalizadeh, B.; Aydinlou, Z.H.; Vaezi, H.; Kashefi, A.; et al. The study of rs324420 (C385A) polymorphism of the FAAH gene of the endocannabinoid system in patients with epilepsy and ADHD. Epilepsy Res. 2023, 192, 107100. [Google Scholar] [CrossRef]
- Horcajadas, F.A.; Piriz, J.R.D.; González, A.P.; Romero, S.S.; Sánchez-Morla, E.; Sánchez, I.A.; Atance, J.A.R. Cannabinoid receptor type 2 gene is associated with comorbidity of schizophrenia and cannabis dependence and fatty acid amide hydrolase gene is associated with cannabis dependence in the Spanish population. Adicciones 2023, 35, 33–46. [Google Scholar] [CrossRef]
- Jacobson, M.R.; Watts, J.J.; Da Silva, T.; Tyndale, R.F.; Rusjan, P.M.; Houle, S.; Wilson, A.A.; Ross, R.A.; Boileau, I.; Mizrahi, R. Fatty acid amide hydrolase is lower in young cannabis users. Addict. Biol. 2021, 26, e12872. [Google Scholar] [CrossRef]
- Hindocha, C.; Freeman, T.P.; Schafer, G.; Gardner, C.; Bloomfield, M.A.P.; Bramon, E.; Morgan, C.J.A.; Curran, H.V. Acute effects of cannabinoids on addiction endophenotypes are moderated by genes encoding the CB1 receptor and FAAH enzyme. Addict. Biol. 2020, 25, e12762. [Google Scholar] [CrossRef]
- Melroy-Greif, W.E.; Wilhelmsen, K.C.; Ehlers, C.L. Genetic variation in FAAH is associated with cannabis use disorders in a young adult sample of Mexican Americans. Drug Alcohol Depend. 2016, 166, 249–253. [Google Scholar] [CrossRef]
- Boileau, I.; Mansouri, E.; Williams, B.; Le Foll, B.; Rusjan, P.; Mizrahi, R.; Tyndale, R.F.; Huestis, M.A.; Payer, D.E.; Wilson, A.A.; et al. Fatty Acid Amide Hydrolase Binding in Brain of Cannabis Users: Imaging with the Novel Radiotracer [11C]CURB. Biol. Psychiatry 2016, 80, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Maple, K.E.; McDaniel, K.A.; Shollenbarger, S.G.; Lisdahl, K.M. Dose-dependent cannabis use, depressive symptoms, and FAAH genotype predict sleep quality in emerging adults: A pilot study. Am. J. Drug Alcohol Abus. 2016, 42, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Shollenbarger, S.G.; Price, J.; Wieser, J.; Lisdahl, K. Poorer frontolimbic white matter integrity is associated with chronic cannabis use, FAAH genotype, and increased depressive and apathy symptoms in adolescents and young adults. NeuroImage Clin. 2015, 8, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M.; Nielsen, D.A.; Kosten, T.R.; De La Garza, R., 2nd; Newton, T.F.; Verrico, C.D. FAAH variant Pro129Thr modulates subjective effects produced by cocaine administration. Am. J. Addict. 2018, 27, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.H.C.; McGeary, J.E.; Knopik, V.S.; Bidwell, L.C.; Metrik, J.M. CNR1 and FAAH variation and affective states induced by marijuana smoking. Am. J. Drug Alcohol Abus. 2019, 45, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Filbey, F.M.; Schacht, J.P.; Myers, U.S.; Chavez, R.S.; Hutchison, K.E. Individual and additive effects of the CNR1 and FAAH genes on brain response to marijuana cues. Neuropsychopharmacology 2010, 35, 967–975. [Google Scholar] [CrossRef]
- Schacht, J.P.; Selling, R.E.; Hutchison, K.E. Intermediate cannabis dependence phenotypes and the FAAH C385A variant: An exploratory analysis. Psychopharmacology 2009, 203, 511–517. [Google Scholar] [CrossRef]
- Haughey, H.M.; Marshall, E.; Schacht, J.P.; Louis, A.; Hutchison, K.E. Marijuana withdrawal and craving: Influence of the cannabinoid receptor 1 (CNR1) and fatty acid amide hydrolase (FAAH) genes. Addiction 2018, 103, 1678–1686. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.; Deng, X.D.; Ma, Y.; Liu, Y. FAAH levels and its genetic polymorphism association with susceptibility to methamphetamine dependence. Ann. Hum. Genet. 2020, 84, 259–270. [Google Scholar] [CrossRef]
- Pu, S.; Eck, P.; Jenkins, D.J.; Connelly, P.W.; Lamarche, B.; Kris-Etherton, P.M.; West, S.G.; Liu, X.; Jones, P.J. Interactions between dietary oil treatments and genetic variants modulate fatty acid ethanolamides in plasma and body weight composition. Br. J. Nutr. 2016, 115, 1012–1023. [Google Scholar] [CrossRef]
- de Luis, D.A.; Sagrado, M.G.; Aller, R.; Izaola, O.; Conde, R. Effects of C358A missense polymorphism of the endocannabinoid degrading enzyme fatty acid amide hydrolase on weight loss after a hypocaloric diet. Metab. Clin. Exp. 2011, 60, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Milano, W.; Petrella, C.; Canestrelli, B.; Maj, M. Endocannabinoid Pro129Thr FAAH functional polymorphism but not 1359G/A CNR1 polymorphism is associated with antipsychotic-induced weight gain. J. Clin. Psychopharmacol. 2010, 30, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Bifulco, M.; Di Filippo, C.; Gazzerro, P.; Canestrelli, B.; Monteleone, F.; Proto, M.C.; Di Genio, M.; Grimaldi, C.; Maj, M. Association of CNR1 and FAAH endocannabinoid gene polymorphisms with anorexia nervosa and bulimia nervosa: Evidence for synergistic effects. Genes Brain Behav. 2009, 8, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Crombie, K.M.; Privratsky, A.A.; Schomaker, C.M.; Heilicher, M.; Ross, M.C.; Sartin-Tarm, A.; Sellnow, K.; Binder, E.B.; Andrew James, G.; Cisler, J.M. The influence of FAAH genetic variation on physiological, cognitive, and neural signatures of fear acquisition and extinction learning in women with PTSD. NeuroImage Clin. 2022, 33, 102922. [Google Scholar] [CrossRef]
- Ney, L.J.; Matthews, A.; Hsu, C.K.; Zuj, D.V.; Nicholson, E.; Steward, T.; Nichols, D.; Graham, B.; Harrison, B.; Bruno, R.; et al. Cannabinoid polymorphisms interact with plasma endocannabinoid levels to predict fear extinction learning. Depress. Anxiety 2021, 38, 1087–1099. [Google Scholar] [CrossRef]
- Zabik, N.L.; Iadipaolo, A.S.; Marusak, H.A.; Peters, C.; Burghardt, K.; Rabinak, C.A. A common genetic variant in fatty acid amide hydrolase is linked to alterations in fear extinction neural circuitry in a racially diverse, nonclinical sample of adults. J. Neurosci. Res. 2022, 100, 744–761. [Google Scholar] [CrossRef]
- Green, D.G.J.; Kim, J.; Kish, S.J.; Tyndale, R.F.; Hill, M.N.; Strafella, A.P.; Tong, J.; McCluskey, T.; Westwood, D.J.; Houle, S.; et al. Fatty acid amide hydrolase binding is inversely correlated with amygdalar functional connectivity: A combined positron emission tomography and magnetic resonance imaging study in healthy individuals. J. Psychiatry Neurosci. 2021, 46, E238–E246. [Google Scholar] [CrossRef]
- Spohrs, J.; Ulrich, M.; Grön, G.; Prost, M.; Plener, P.L.; Fegert, J.M.; Bindila, L.; Abler, B. Fear extinction learning and anandamide: An fMRI study in healthy humans. Transl. Psychiatry 2021, 11, 161. [Google Scholar] [CrossRef]
- Jiang, Y.; Nie, Y.; Li, Y.; Zhang, L. Association of cannabinoid type 1 receptor and fatty acid amide hydrolase genetic polymorphisms in Chinese patients with irritable bowel syndrome. J. Gastroenterol. Hepatol. 2014, 29, 1186–1191. [Google Scholar] [CrossRef]
- Wong, B.S.; Camilleri, M.; Busciglio, I.; Carlson, P.; Szarka, L.A.; Burton, D.; Zinsmeister, A.R. Pharmacogenetic trial of a cannabinoid agonist shows reduced fasting colonic motility in patients with nonconstipated irritable bowel syndrome. Gastroenterology 2011, 141, 1638–1647.e477. [Google Scholar] [CrossRef]
- Doris, J.M.; Millar, S.A.; Idris, I.; O’Sullivan, S.E. Genetic polymorphisms of the endocannabinoid system in obesity and diabetes. Diabetes Obes. Metab. 2019, 21, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Brönner, G.; Wandolski, M.; Carrie, J.; Nguyen, T.T.; Greene, B.H.; Scherag, A.; Grallert, H.; Vogel, C.I.; Scherag, S.; et al. Mutation screen and association studies for the fatty acid amide hydrolase (FAAH) gene and early onset and adult obesity. BMC Med. Genet. 2010, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Durand, E.; Lecoeur, C.; Delplanque, J.; Benzinou, M.; Degraeve, F.; Boutin, P.; Marre, M.; Balkau, B.; Charpentier, G.; Froguel, P.; et al. Evaluating the association of FAAH common gene variation with childhood, adult severe obesity and type 2 diabetes in the French population. Obes. Facts 2008, 1, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Chidambaran, V.; Pilipenko, V.; Spruance, K.; Venkatasubramanian, R.; Niu, J.; Fukuda, T.; Mizuno, T.; Zhang, K.; Kaufman, K.; Vinks, A.A.; et al. Fatty acid amide hydrolase-morphine interaction influences ventilatory response to hypercapnia and postoperative opioid outcomes in children. Pharmacogenomics 2017, 18, 143–156. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Zhang, X.; Chidambaran, V.; Mavi, J.; Pilipenko, V.; Mersha, T.B.; Meller, J.; Kaufman, K.M.; Martin, L.J.; McAuliffe, J. Novel associations between FAAH genetic variants and postoperative central opioid-related adverse effects. Pharmacogenom. J. 2015, 15, 436–442. [Google Scholar] [CrossRef]
- Dlugos, A.M.; Hamidovic, A.; Hodgkinson, C.A.; Goldman, D.; Palmer, A.A.; de Wit, H. More aroused, less fatigued: Fatty acid amide hydrolase gene polymorphisms influence acute response to amphetamine. Neuropsychopharmacology 2010, 35, 613–622. [Google Scholar] [CrossRef]
- Chmelikova, M.; Pacal, L.; Spinarova, L.; Vasku, A. Association of polymorphisms in the endocannabinoid system genes with myocardial infarction and plasma cholesterol levels. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc. Czech. 2015, 159, 535–539. [Google Scholar] [CrossRef]
- Presciuttini, S.; Carli, G.; Santarcangelo, E.L. Hypnotizability-related FAAH C385A polymorphism: Possible endocannabinoid contribution to suggestion-induced analgesia. Int. J. Clin. Exp. Hypn. 2020, 68, 29–37. [Google Scholar] [CrossRef]
- Colloca, L.; Wang, Y.; Martinez, P.E.; Chang, Y.C.; Ryan, K.A.; Hodgkinson, C.; Goldman, D.; Dorsey, S.G. OPRM1 rs1799971, COMT rs4680, and FAAH rs324420 genes interact with placebo procedures to induce hypoalgesia. Pain 2019, 160, 1824–1834. [Google Scholar] [CrossRef]
- Cajanus, K.; Holmström, E.J.; Wessman, M.; Anttila, V.; Kaunisto, M.A.; Kalso, E. Effect of endocannabinoid degradation on pain: Role of FAAH polymorphisms in experimental and postoperative pain in women treated for breast cancer. Pain 2016, 157, 361–369. [Google Scholar] [CrossRef]
- Vazquez-Roque, M.I.; Camilleri, M.; Vella, A.; Carlson, P.; Laugen, J.; Zinsmeister, A.R. Association of genetic variation in cannabinoid mechanisms and gastric motor functions and satiation in overweight and obesity. Neurogastroenterol. Motil. 2011, 23, 637-e257. [Google Scholar] [CrossRef] [PubMed]
- Knoll, N.; Volckmar, A.L.; Pütter, C.; Scherag, A.; Kleber, M.; Hebebrand, J.; Hinney, A.; Reinehr, T. The fatty acid amide hydrolase (FAAH) gene variant rs324420 AA/AC is not associated with weight loss in a 1-year lifestyle intervention for obese children and adolescents. Horm. Metab. Res. 2012, 44, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Atieh, J.; Maselli, D.; Breen-Lyles, M.; Torres, M.; Katzka, D.; Ryks, M.; Busciglio, I.; Burton, D.; Carlson, P.; Harmsen, W.S.; et al. Cannabidiol for Functional Dyspepsia With Normal Gastric Emptying: A Randomized Controlled Trial. Am. J. Gastroenterol. 2022, 117, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sonnenberg, G.E.; Baye, T.M.; Littrell, J.; Gunnell, J.; DeLaForest, A.; MacKinney, E.; Hillard, C.J.; Kissebah, A.H.; Olivier, M.; et al. Obesity-related dyslipidemia associated with FAAH, independent of insulin response, in multigenerational families of Northern European descent. Pharmacogenomics 2009, 10, 1929–1939. [Google Scholar] [CrossRef]
- Wong, B.S.; Camilleri, M.; Eckert, D.; Carlson, P.; Ryks, M.; Burton, D.; Zinsmeister, A.R. Randomized pharmacodynamic and pharmacogenetic trial of dronabinol effects on colon transit in irritable bowel syndrome-diarrhea. Neurogastroenterol. Motil. 2012, 24, 358-e169. [Google Scholar] [CrossRef]
- Jensen, D.P.; Andreasen, C.H.; Andersen, M.K.; Hansen, L.; Eiberg, H.; Borch-Johnsen, K.; Jørgensen, T.; Hansen, T.; Pedersen, O. The functional Pro129Thr variant of the FAAH gene is not associated with various fat accumulation phenotypes in a population-based cohort of 5801 whites. J. Mol. Med. 2007, 85, 445–449. [Google Scholar] [CrossRef]
- Demaili, A.; Portugalov, A.; Dudai, M.; Maroun, M.; Akirav, I.; Braun, K.; Bock, J. Epigenetic (re)programming of gene expression changes of CB1R and FAAH in the medial prefrontal cortex in response to early life and adolescence stress exposure. Front. Cell. Neurosci. 2023, 17, 1129946. [Google Scholar] [CrossRef]
- Massa, F.; Marsicano, G.; Hermann, H.; Cannich, A.; Monory, K.; Cravatt, B.F.; Ferri, G.L.; Sibaev, A.; Storr, M.; Lutz, B. The endogenous cannabinoid system protects against colonic inflammation. J. Clin. Investig. 2004, 113, 1202–1209. [Google Scholar] [CrossRef]
- van der Stelt, M.; Di Marzo, V. The endocannabinoid system in the basal ganglia and in the mesolimbic reward system: Implications for neurological and psychiatric disorders. Eur. J. Pharmacol. 2003, 480, 133–150. [Google Scholar] [CrossRef]
- Schmidt, M.E.; Liebowitz, M.R.; Stein, M.B.; Grunfeld, J.; Van Hove, I.; Simmons, W.K.; Van Der Ark, P.; Palmer, J.A.; Saad, Z.S.; Pemberton, D.J.; et al. The effects of inhibition of fatty acid amide hydrolase (FAAH) by JNJ-42165279 in social anxiety disorder: A double-blind, randomized, placebo-controlled proof-of-concept study. Neuropsychopharmacology 2021, 46, 1004–1010. [Google Scholar] [CrossRef]
- Mendizábal, V.E.; Adler-Graschinsky, E. Cannabinoid system as a potential target for drug development in the treatment of cardiovascular disease. Curr. Vasc. Pharmacol. 2003, 1, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Hampson, A.J.; Baler, R.D. Don’t Worry, Be Happy: Endocannabinoids and Cannabis at the Intersection of Stress and Reward. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Wotjak, C.T.; Azad, S.C.; Bisogno, T.; Rammes, G.; Cascio, M.G.; Hermann, H.; Tang, J.; Hofmann, C.; Zieglgänsberger, W.; et al. The endogenous cannabinoid system controls extinction of aversive memories. Nature 2002, 418, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dey, S.K.; Maccarrone, M. Jekyll and hyde: Two faces of cannabinoid signaling in male and female fertility. Endocr. Rev. 2006, 27, 427–448. [Google Scholar] [CrossRef] [PubMed]
- Viveros, M.P.; de Fonseca, F.R.; Bermudez-Silva, F.J.; McPartland, J.M. Critical role of the endocannabinoid system in the regulation of food intake and energy metabolism, with phylogenetic, developmental, and pathophysiological implications. Endocr. Metab. Immune Disord. Drug Targets 2008, 8, 220–230. [Google Scholar] [CrossRef]
- Maccarrone, M.; De Petrocellis, L.; Bari, M.; Fezza, F.; Salvati, S.; Di Marzo, V.; Finazzi-Agrò, A. Lipopolysaccharide downregulates fatty acid amide hydrolase expression and increases anandamide levels in human peripheral lymphocytes. Arch. Biochem. Biophys. 2001, 393, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Zaplatic, E.; Micioni Di Bonaventura, M.V.; Micioni Di Bonaventura, E.; De Cristofaro, P.; Maccarrone, M.; Cifani, C.; D’Addario, C. On the Role of Central Type-1 Cannabinoid Receptor Gene Regulation in Food Intake and Eating Behaviors. Int. J. Mol. Sci. 2021, 22, 398. [Google Scholar] [CrossRef]
- Gunduz-Cinar, O.; MacPherson, K.P.; Cinar, R.; Gamble-George, J.; Sugden, K.; Williams, B.; Godlewski, G.; Ramikie, T.S.; Gorka, A.X.; Alapafuja, S.O.; et al. Convergent translational evidence of a role for anandamide in amygdala-mediated fear extinction, threat processing and stress-reactivity. Mol. Psychiatry 2013, 18, 813–823. [Google Scholar] [CrossRef]
- Freund, T.F.; Katona, I.; Piomelli, D. Role of endogenous cannabinoids in synaptic signaling. Physiol. Rev. 2003, 83, 1017–1066. [Google Scholar] [CrossRef]
- Difede, J.; Rothbaum, B.O.; Rizzo, A.A.; Wyka, K.; Spielman, L.; Reist, C.; Roy, M.J.; Jovanovic, T.; Norrholm, S.D.; Cukor, J.; et al. Enhancing exposure therapy for posttraumatic stress disorder (PTSD): A randomized clinical trial of virtual reality and imaginal exposure with a cognitive enhancer. Transl. Psychiatry 2022, 12, 299. [Google Scholar] [CrossRef]
- Ahmadalipour, A.; Fanid, L.M.; Zeinalzadeh, N.; Alizadeh, M.; Vaezi, H.; Aydinlou, Z.H.; Noorazar, S.G. The first evidence of an association between a polymorphism in the endocannabinoid-degrading enzyme FAAH (FAAH rs2295633) with attention deficit hyperactivity disorder. Genomics 2020, 112, 1330–1334. [Google Scholar] [CrossRef] [PubMed]
- Yagin, N.L.; Aliasgari, F.; Alizadeh, M.; Aliasgharzadeh, S.; Mahdavi, R. Comparison of endocannabinoids levels, FAAH gene polymorphisms, and appetite regulatory substances in women with and without binge eating disorder: A crosssectional study. Nutr. Res. 2020, 83, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological interactions between stress and the endocannabinoid system. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Chadha, S.; Sachdeva, M.; Sehgal, A.; Kumar, A.; Venkatachalam, T.; Hafeez, A.; Aleya, L.; Arora, S.; Batiha, G.E.; et al. Understanding the possible role of endocannabinoid system in obesity. Prostaglandins Other Lipid Mediat. 2021, 152, 106520. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Lecue, I.; Pilar-Cuéllar, F.; Muguruza, C.; Florensa-Zanuy, E.; Díaz, Á.; Urigüen, L.; Castro, E.; Pazos, A.; Callado, L.F. The endocannabinoid system in mental disorders: Evidence from human brain studies. Biochem. Pharmacol. 2018, 157, 97–107. [Google Scholar] [CrossRef]
- Woodhams, S.G.; Sagar, D.R.; Burston, J.J.; Chapman, V. The role of the endocannabinoid system in pain. Handb. Exp. Pharmacol. 2015, 227, 119–143. [Google Scholar] [CrossRef]
- Laredo, S.A.; Marrs, W.R.; Parsons, L.H. Endocannabinoid signaling in reward and addiction: From homeostasis to pathology. In Endocannabinoids and Lipid Mediators in Brain Functions; Melis, M., Ed.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Monteleone, P.; Tortorella, A.; Martiadis, V.; Di Filippo, C.; Canestrelli, B.; Maj, M. The cDNA 385C to A missense polymorphism of the endocannabinoid degrading enzyme fatty acid amide hydrolase (FAAH) is associated with overweight/obesity but not with binge eating disorder in overweight/obese women. Psychoneuroendocrinology 2008, 33, 546–550. [Google Scholar] [CrossRef]
- Lutz, B.; Marsicano, G.; Maldonado, R.; Hillard, C.J. The endocannabinoid system in guarding against fear, anxiety and stress. Nat. Rev. Neurosci. 2015, 16, 705–718. [Google Scholar] [CrossRef]
- Ensembl Database. Available online: https://www.ensembl.org/index.html (accessed on 24 August 2023).
- Ensembl Home. rs324420 (SNP)—Population Genetics—Homo Sapiens—Ensembl Genome Browser 89. Available online: http://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=1:46404589-46405589;v=rs324420;vdb=variation;vf=200953 (accessed on 24 August 2023).
- Woodhams, S.G.; Chapman, V.; Finn, D.P.; Hohmann, A.G.; Neugebauer, V. The cannabinoid system and pain. Neuropharmacology 2017, 124, 105–120. [Google Scholar] [CrossRef]
- Medeiros, D.C.; Cota, V.R.; Oliveira, A.C.P.; Moreira, F.A.; Moraes, M.F.D. The Endocannabinoid System Activation as a Neural Network Desynchronizing Mediator for Seizure Suppression. Front. Behav. Neurosci. 2020, 14, 603245. [Google Scholar] [CrossRef]
- Ginevičienė, V.; Utkus, A.; Pranckevičienė, E.; Semenova, E.A.; Hall, E.; Ahmetov, I.I. Perspectives in Sports Genomics. Biomedicines 2022, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; Karacabeyli, E.S.; Gorzalka, B.B. Estrogen recruits the endocannabinoid system to modulate emotionality. Psychoneuroendocrinology 2007, 32, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Di, S.; Malcher-Lopes, R.; Marcheselli, V.L.; Bazan, N.G.; Tasker, J.G. Rapid glucocorticoid-mediated endocannabinoid release and opposing regulation of glutamate and γ-aminobutyric acid inputs to hypothalamic magnocellular neurons. Endocrinology 2005, 146, 4292–4301. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; McLaughlin, R.J.; Pan, B.; Fitzgerald, M.L.; Roberts, C.J.; Lee, T.T.; Karatsoreos, I.N.; Mackie, K.; Viau, V.; Pickel, V.M.; et al. Recruitment of prefrontal cortical endocannabinoid signaling by glucocorticoids contributes to termination of the stress response. J. Neurosci. 2011, 31, 10506–10515. [Google Scholar] [CrossRef]
- Evanson, N.K.; Tasker, J.G.; Hill, M.N.; Hillard, C.J.; Herman, J.P. Fast feedback inhibition of the HPA axis by glucocorticoids is mediated by endocannabinoid signaling. Endocrinology 2010, 151, 4811–4819. [Google Scholar] [CrossRef]
- Russo, S.J.; Murrough, J.W.; Han, M.H.; Charney, D.S.; Nestler, E.J. Neurobiology of resilience. Nat. Neurosci. 2012, 15, 1475–1484. [Google Scholar] [CrossRef]
- Hill, M.N.; McLaughlin, R.J.; Morrish, A.C.; Viau, V.; Floresco, S.B.; Hillard, C.J.; Gorzalka, B.B. Suppression of amygdalar endocannabinoid signaling by stress contributes to activation of the hypothalamic–pituitary–adrenal axis. Neuropsychopharmacology 2009, 34, 2733–2745. [Google Scholar] [CrossRef]
- Gray, J.M.; Vecchiarelli, H.A.; Morena, M.; Lee, T.T.; Hermanson, D.J.; Kim, A.B.; McLaughlin, R.J.; Hassan, K.I.; Kühne, C.; Wotjak, C.T.; et al. Corticotropin-releasing hormone drives anandamide hydrolysis in the amygdala to promote anxiety. J. Neurosci. 2015, 35, 3879–3892. [Google Scholar] [CrossRef]
- Hariri, A.R.; Gorka, A.; Hyde, L.W.; Kimak, M.; Halder, I.; Ducci, F.; Ferrell, R.E.; Goldman, D.; Manuck, S.B. Divergent ef-fects of genetic variation in endocannabinoid signaling on human threat- and reward-related brain function. Biol. Psychiatry 2009, 66, 9–16. [Google Scholar] [CrossRef]
- Dincheva, I.; Drysdale, A.T.; Hartley, C.A.; Johnson, D.C.; Jing, D.; King, E.C.; Ra, S.; Gray, J.M.; Yang, R.; DeGruccio, A.M.; et al. FAAH genetic variation enhances fronto-amygdala function in mouse and human. Nat. Commun. 2015, 6, 6395. [Google Scholar] [CrossRef]
- Gee, D.G.; Fetcho, R.N.; Jing, D.; Li, A.; Glatt, C.E.; Drysdale, A.T.; Cohen, A.O.; Dellarco, D.V.; Yang, R.R.; Dale, A.M.; et al. Individual differences in frontolimbic circuitry and anxiety emerge with adolescent changes in endocannabinoid signaling across species. Proc. Natl. Acad. Sci. USA 2016, 113, 4500–4505. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, P.A.; Ramchandani, V.A.; Schwandt, M.L.; Kwako, L.E.; George, D.T.; Mayo, L.M.; Hillard, C.J.; Heilig, M. FAAH gene variation moderates stress response and symptom severity in patients with posttraumatic stress disorder and comorbid alcohol dependence. Alcohol. Clin. Exp. Res. 2016, 40, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Mayo, L.M.; Asratian, A.; Linde, J.; Holm, L.; Natt, D.; Augier, G.; Stensson, N.; Vecchiarelli, H.A.; Balsevich, G.; Aukema, R.J.; et al. Protective effects of elevated anandamide on stress and fear-related behaviors: Translational evidence from humans and mice. Mol. Psychiatry 2018, 25, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Roques, B.P.; Fournie-Zaluski, M.C.; Wurm, M. Inhibiting the breakdown of endogenous opioids and cannabinoids to alleviate pain. Nat. Rev. Drug Discov. 2012, 11, 292–310. [Google Scholar] [CrossRef]
- Clapper, J.R.; Moreno-Sanz, G.; Russo, R.; Guijarro, A.; Vacondio, F.; Duranti, A.; Tontini, A.; Sanchini, S.; Sciolino, N.R.; Spradley, J.M.; et al. Anandamide suppresses pain initiation through a peripheral endocannabinoid mechanism. Nat. Neurosci. 2010, 13, 1265–1270. [Google Scholar] [CrossRef]
- Greenbaum, L.; Tegeder, I.; Barhum, Y.; Melamed, E.; Roditi, Y.; Djaldetti, R. Contribution of genetic variants to pain susceptibility in Parkinson disease. Eur. J. Pain 2012, 16, 1243–1250. [Google Scholar] [CrossRef]
- Khasabova, I.A.; Holman, M.; Morse, T.; Burlakova, N.; Coicou, L.; Harding Rose, C.; Simone, D.A.; Seybold, V.S. Increased anandamide uptake by sensory neurons contributes to hyperalgesia in a model of cancer pain. Neurobiol. Dis. 2013, 58, 19–28. [Google Scholar] [CrossRef]
- Pini, A.; Mannaioni, G.; Pellegrini-Giampietro, D.; Passani, M.B.; Mastroianni, R.; Bani, D.; Masini, E. The role of cannabinoids in inflammatory modulation of allergic respiratory disorders, inflammatory pain and ischemic stroke. Curr. Drug Targets 2012, 13, 984–993. [Google Scholar] [CrossRef]
- Hohmann, A.G.; Suplita, R.L.; Bolton, N.M.; Neely, M.H.; Fegley, D.; Mangieri, R.; Krey, J.F.; Walker, J.M.; Holmes, P.V.; Crystal, J.D.; et al. An endocannabinoid mechanism for stress-induced analgesia. Nature 2005, 435, 1108–1112. [Google Scholar] [CrossRef]
- Grillo, A.; Fezza, F.; Chemi, G.; Colangeli, R.; Brogi, S.; Fazio, D.; Federico, S.; Papa, A.; Relitti, N.; Di Maio, R.; et al. Selective fatty acid amide hydrolase inhibitors as potential novel antiepileptic agents. ACS Chem. Neurosci. 2021, 12, 1716–1736. [Google Scholar] [CrossRef]
- Centonze, D.; Bari, M.; Di Michele, B.; Rossi, S.; Gasperi, V.; Pasini, A.; Battista, N.; Bernardi, G.; Curatolo, P.; Maccarrone, M. Altered anandamide degradation in attention-deficit/hyperactivity disorder. Neurology 2009, 72, 1526–1527. [Google Scholar] [CrossRef] [PubMed]
- Beghi, E. The epidemiology of epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Buono, R.J.; Bradfield, J.P.; Wei, Z.; Sperling, M.R.; Dlugos, D.J.; Privitera, M.D.; French, J.A.; Lo, W.; Cossette, P.; Schachter, S.C.; et al. Genetic variation in PADI6-PADI4 on 1p36.13 is associated with common forms of human generalized epilepsy. Genes 2021, 12, 1441. [Google Scholar] [CrossRef] [PubMed]
- Sayal, K.; Prasad, V.; Daley, D.; Ford, T.; Coghill, D. ADHD in children and young people: Prevalence, care pathways, and service provision. Lancet Psychiatry 2018, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.E.; Williams, E.; Seegobin, S.; Tye, C.; Kuntsi, J.; Asherson, P. Cannabinoids in attention-deficit/hyperactivity disorder: A randomised-controlled trial. Eur. Neuropsychopharmacol. 2017, 27, 795–808. [Google Scholar] [CrossRef]
- De Caro, C.; Leo, A.; Citraro, R.; De Sarro, C.; Russo, R.; Calignano, A.; Russo, E. The potential role of cannabinoids in epilepsy treatment. Expert Rev. Neurother. 2017, 17, 1069–1079. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef]
- Niemela, G.; Terry, G.E. Contribution of Fatty Acid Amide Hydrolase to Alcohol Use Disorder: A Systematic Review. Cannabis Cannabinoid Res. 2021, 6, 105–118. [Google Scholar] [CrossRef]
- Parsons, L.H.; Hurd, Y.L. Endocannabinoid signalling in reward and addiction. Nat. Rev. Neurosci. 2015, 16, 579–594. [Google Scholar] [CrossRef]
- Sipe, J.C.; Chiang, K.; Gerber, A.L.; Beutler, E.; Cravatt, B.F. A missense mutation in human fatty acid amide hydrolase associated with problem drug use. Proc. Natl. Acad. Sci. USA 2002, 99, 8394–8399. [Google Scholar] [CrossRef]
- Dongdem, J.T.; Helegbe, G.K.; Opare-Asamoah, K.; Wezena, C.A.; Ocloo, A. Assessment of NSAIDs as potential inhibitors of the fatty acid amide hydrolase I (FAAH-1) using three different primary fatty acid amide substrates in vitro. BMC Pharmacol. Toxicol. 2022, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.G.; Wackerhage, H. Genetic testing of athletes. Med. Sport Sci. 2009, 54, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, I.D.; Lucia, A.; Pitsiladis, Y.P.; Pushkarev, V.P.; Dyatlov, D.A.; Orekhov, E.F.; Artioli, G.G.; Guilherme, J.P.; Lancha, A.H., Jr.; Ginevičienė, V.; et al. ACTN3 R577X and ACE I/D gene variants influence performance in elite sprinters: A multi-cohort study. BMC Genom. 2016, 17, 285. [Google Scholar] [CrossRef]
- Ben-Zaken, S.; Meckel, Y.; Nemet, D.; Kassem, E.; Eliakim, A. The combined frequencies of the IL-6 G-174C and IGFBP3 A-202C polymorphisms among swimmers and runners. Growth Horm. IGF Res. 2020, 51, 17–21. [Google Scholar] [CrossRef]
- Ben-Zaken, S.; Meckel, Y.; Nemet, D.; Kassem, E.; Eliakim, A. Genetic basis for the dominance of Israeli long-distance runners of Ethiopian origin. J. Strength Cond. Res. 2019, 35, 1885–1896. [Google Scholar] [CrossRef] [PubMed]
- Amorim, T.; Durães, C.; Machado, J.C.; Metsios, G.S.; Wyon, M.; Maia, J.; Flouris, A.D.; Marques, F.; Nogueira, L.; Adubeiro, N.; et al. Genetic variation in Wnt/β-catenin and ER signalling pathways in female and male elite dancers and its associations with low bone mineral density: A cross-section and longitudinal study. Osteoporos. Int. 2018, 29, 2261–2274. [Google Scholar] [CrossRef]
- Salles, J.I.; Lopes, L.R.; Duarte, M.; Morrissey, D.; Martins, M.B.; Machado, D.E.; Guimarães, J.; Perini, J.A. Fc receptor-like 3 (-169T>C) polymorphism increases the risk of tendinopathy in volleyball athletes: A case control study. BMC Med. Genet. 2018, 19, 119. [Google Scholar] [CrossRef]
- Vitale, J.A.; Castellini, G.; Gianola, S.; Banfi, G. Analysis of the Christiania stop in roller hockey players with and without previous groin pain: A prospective case series study. Sports Sci. Health 2019, 15, 641–646. [Google Scholar] [CrossRef]





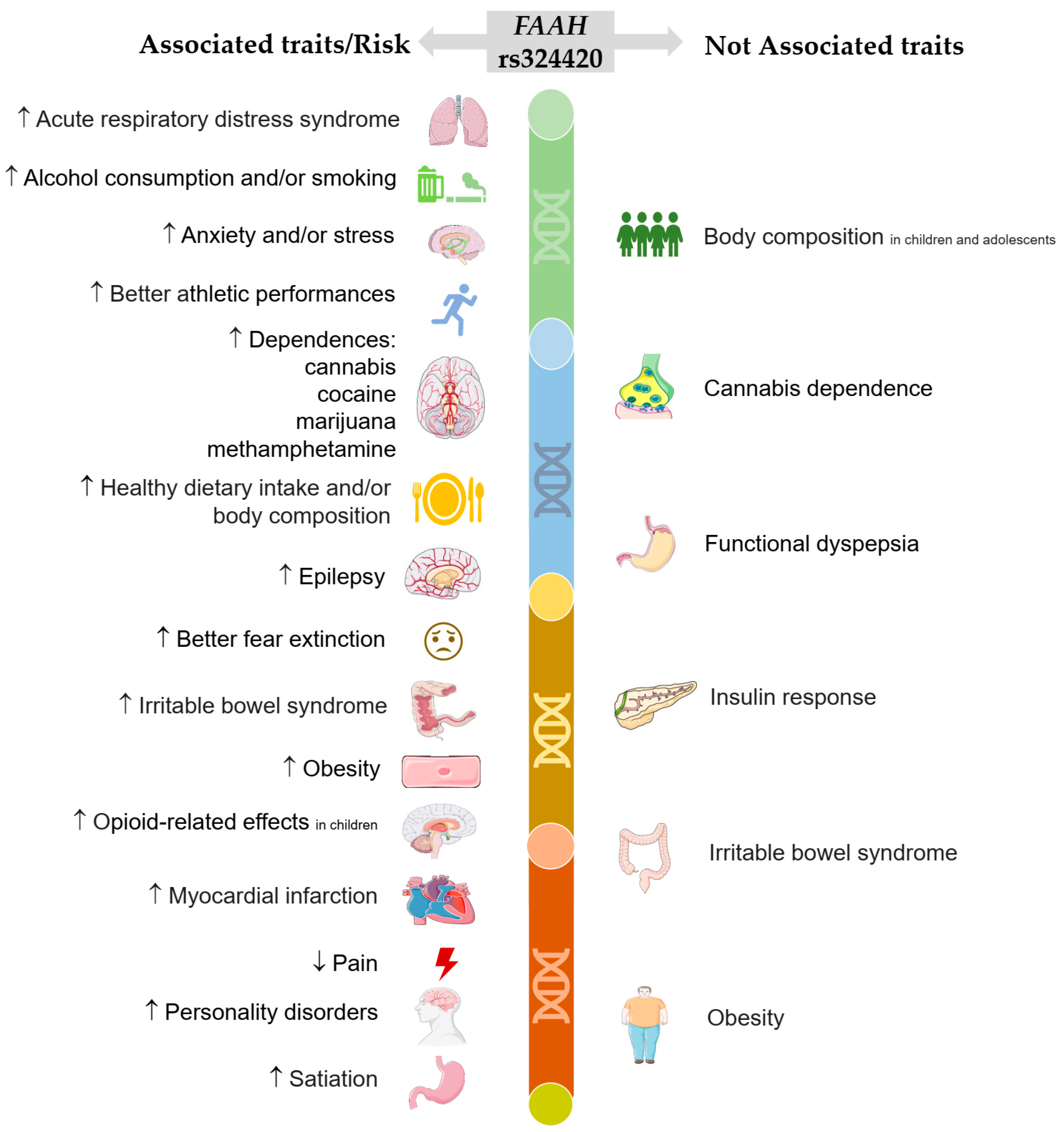{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Population | Athletes’ Profile | Genotypes, n (%) | ||
|---|---|---|---|---|---|
| AA | AC | CC | |||
| Silva et al. [6] | 75.1% Portuguese and 24.9% Others | Elite volleyball players (n = 219) | 11 (5.0) | 74 (33.8) | 134 (61.2) |
| 75.8% Portuguese and 24.2% Others | Female (n = 66) | 3 (5.0) | 20 (33.3) | 37 (61.7) | |
| 74.8% Portuguese and 25.2% Others | Male (n = 162) | 8 (5.0) | 54 (34.0) | 97 (61.0) | |
| Silva et al. [5] | 82.8% Portuguese and 17.2% Others | Elite rink-hockey players (n = 116) | 4 (3.4) | 34 (29.3) | 78 (67.2) |
| All Portuguese | Female (n = 18) | 1 (5.6) | 6 (33.3) | 11 (61.1) | |
| 79.6% Portuguese and 20.4% Others | Male (n = 98) | 3 (3.1) | 28 (28.6) | 67 (68.4) | |
| Peplonska et al. [2] | All Caucasians of Polish origin | Elite athletes (n = 621, 29.3% females) | 27 (4.3) | 259 (41.7) | 335 (54.0) |
| Power (n = 183) | 6 (3.3) | 79 (43.2) | 98 (53.5) | ||
| Endurance (n = 212) | 8 (3.8) | 84 (39.6) | 120 (56.6) | ||
| Combat (n = 226) | 13 (5.8) | 96 (42.4) | 117 (51.8) | ||
| Peplonska et al. [14] | All Caucasians of Polish origin | Elite athletes (n = 413, 36.3% females) | 16 (3.9) | 169 (40.9) | 228 (55.2) |
| Power (n = 188) | 6 (3.2) | 80 (42.6) | 102 (54.2) | ||
| Endurance (n = 225) | 10 (4.4) | 89 (39.6) | 126 (56.0) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, H.-H.; Tavares, V.; Neto, B.V.; Cerqueira, F.; Medeiros, R.; Silva, M.-R.G. FAAH rs324420 Polymorphism: Biological Pathways, Impact on Elite Athletic Performance and Insights for Sport Medicine. Genes 2023, 14, 1946. https://doi.org/10.3390/genes14101946
Silva H-H, Tavares V, Neto BV, Cerqueira F, Medeiros R, Silva M-RG. FAAH rs324420 Polymorphism: Biological Pathways, Impact on Elite Athletic Performance and Insights for Sport Medicine. Genes. 2023; 14(10):1946. https://doi.org/10.3390/genes14101946
Chicago/Turabian StyleSilva, Hugo-Henrique, Valéria Tavares, Beatriz Vieira Neto, Fátima Cerqueira, Rui Medeiros, and Maria-Raquel G. Silva. 2023. "FAAH rs324420 Polymorphism: Biological Pathways, Impact on Elite Athletic Performance and Insights for Sport Medicine" Genes 14, no. 10: 1946. https://doi.org/10.3390/genes14101946
APA StyleSilva, H. -H., Tavares, V., Neto, B. V., Cerqueira, F., Medeiros, R., & Silva, M. -R. G. (2023). FAAH rs324420 Polymorphism: Biological Pathways, Impact on Elite Athletic Performance and Insights for Sport Medicine. Genes, 14(10), 1946. https://doi.org/10.3390/genes14101946