
Predisposition of the Common MC4R rs17782313 Female Carriers to Elevated Obesity and Interaction with Eating Habits


Abstract
:1. Introduction

2. Materials and Methods
2.1. Participants
2.2. Anthropometric and Genetic Data
2.3. Eating Behaviors Variables
2.4. Statistical Analysis
3. Results
3.1. Participants
3.2. MC4R rs17782313 and Obesity Risk
3.3. Interactions of MC4R rs17782313 with Eating Behaviors on Obesity Risk
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 18 September 2023).
- Wardle, J.; Carnell, S.; Haworth, C.M.; Plomin, R. Evidence for a Strong Genetic Influence on Childhood Adiposity despite the Force of the Obesogenic Environment. Am. J. Clin. Nutr. 2008, 87, 398–404. [Google Scholar]
- Mahmoud, R.; Kimonis, V.; Butler, M.G. Genetics of Obesity in Humans: A Clinical Review. Int. J. Mol. Sci. 2022, 23, 11005. [Google Scholar]
- Nicolaidis, S. Environment and Obesity. Metabolism 2019, 100S, 153942. [Google Scholar] [CrossRef] [PubMed]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Impact of Insufficient Sleep on Total Daily Energy Expenditure, Food Intake, and Weight Gain. Proc. Natl. Acad. Sci. USA 2013, 110, 5695–5700. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Southerland, J.; Wang, K.; Bailey, B.A.; Alamian, A.; Stevens, M.A.; Wang, Y. Ethnic Differences in Risk Factors for Obesity among Adults in California, the United States. J. Obes. 2017, 2017, 2427483. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, F.Q. Body Mass Index. Nutr. Today 2015, 50, 117–128. [Google Scholar]
- Milaneschi, Y.; Simmons, W.K.; van Rossum, E.F.C.; Penninx, B.W. Depression and Obesity: Evidence of Shared Biological Mechanisms. Mol. Psychiatry 2019, 24, 18–33. [Google Scholar] [PubMed]
- Smith, K.B.; Smith, M.S. Obesity Statistics. Prim. Care 2016, 43, 121–135. [Google Scholar]
- Tremmel, M.; Gerdtham, U.-G.; Nilsson, P.M.; Saha, S. Economic Burden of Obesity: A Systematic Literature Review. Int. J. Environ. Res. Public Health 2017, 14, 435. [Google Scholar] [CrossRef]
- Okunogbe, A.; Nugent, R.; Spencer, G.; Ralston, J.; Wilding, J. Economic Impacts of Overweight and Obesity: Current and Future Estimates for Eight Countries. BMJ Glob. Health 2021, 6, e006351. [Google Scholar] [CrossRef]
- Dakanalis, A.; Mentzelou, M.; Papadopoulou, S.K.; Papandreou, D.; Spanoudaki, M.; Vasios, G.K.; Pavlidou, E.; Mantzorou, M.; Giaginis, C. The Association of Emotional Eating with Overweight/Obesity, Depression, Anxiety/Stress, and Dietary Patterns: A Review of the Current Clinical Evidence. Nutrients 2023, 15, 1173. [Google Scholar] [CrossRef]
- Salman, E.J.; Kabir, R. Night Eating Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ljubičić, M.; Matek Sarić, M.; Klarin, I.; Rumbak, I.; Colić Barić, I.; Ranilović, J.; Dželalija, B.; Sarić, A.; Nakić, D.; Djekic, I.; et al. Emotions and Food Consumption: Emotional Eating Behavior in a European Population. Foods 2023, 12, 872. [Google Scholar] [CrossRef]
- Loos, R.J.F.; Yeo, G.S.H. The Genetics of Obesity: From Discovery to Biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [PubMed]
- Chami, N.; Preuss, M.; Walker, R.W.; Moscati, A.; Loos, R.J.F. The Role of Polygenic Susceptibility to Obesity among Carriers of Pathogenic Mutations in MC4R in the UK Biobank Population. PLoS Med. 2020, 17, e1003196. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Lelliott, C.J.; Vidal-Puig, A. Hypothalamic fatty acid metabolism: A housekeeping pathway that regulates food intake. Bioessays 2007, 29, 248–261. [Google Scholar] [CrossRef]
- Yeo, G.S.H.; Chao, D.H.M.; Siegert, A.-M.; Koerperich, Z.M.; Ericson, M.D.; Simonds, S.E.; Larson, C.M.; Luquet, S.; Clarke, I.; Sharma, S.; et al. The Melanocortin Pathway and Energy Homeostasis: From Discovery to Obesity Therapy. Mol. Metab. 2021, 48, 101206. [Google Scholar] [PubMed]
- Gonçalves, J.P.L.; Palmer, D.; Meldal, M. MC4R Agonists: Structural Overview on Antiobesity Therapeutics. Trends Pharmacol. Sci. 2018, 39, 402–423. [Google Scholar] [CrossRef] [PubMed]
- Ju, S.H.; Cho, G.-B.; Sohn, J.-W. Understanding Melanocortin-4 Receptor Control of Neuronal Circuits: Toward Novel Therapeutics for Obesity Syndrome. Pharmacol. Res. 2018, 129, 10–19. [Google Scholar] [CrossRef]
- Sharma, S.; Garfield, A.S.; Shah, B.; Kleyn, P.; Ichetovkin, I.; Moeller, I.H.; Mowrey, W.R.; Van der Ploeg, L.H.T. Current Mechanistic and Pharmacodynamic Understanding of Melanocortin-4 Receptor Activation. Molecules 2019, 24, 1892. [Google Scholar] [CrossRef]
- Baldini, G.; Phelan, K.D. The Melanocortin Pathway and Control of Appetite-Progress and Therapeutic Implications. J. Endocrinol. 2019, 241, R1–R33. [Google Scholar]
- Kühnen, P.; Krude, H.; Biebermann, H. Melanocortin-4 Receptor Signalling: Importance for Weight Regulation and Obesity Treatment. Trends Mol. Med. 2019, 25, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.B.; Harlan, S.M.; Morgan, D.A.; Cui, H.; Rahmouni, K. Differential Contribution of POMC and AgRP Neurons to the Regulation of Regional Autonomic Nerve Activity by Leptin. Mol. Metab. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Szalanczy, A.M.; Key, C.-C.C.; Solberg Woods, L.C. Genetic Variation in Satiety Signaling and Hypothalamic Inflammation: Merging Fields for the Study of Obesity. J. Nutr. Biochem. 2022, 101, 108928. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, U.; Kidd, E.; Malagamuwa, T.; Walley, A. Genetics of Severe Obesity. Curr. Diab. Rep. 2018, 18, 85. [Google Scholar] [CrossRef]
- Yu, K.; Li, L.; Zhang, L.; Guo, L.; Wang, C. Association between MC4R Rs17782313 Genotype and Obesity: A Meta-Analysis. Gene 2020, 733, 144372. [Google Scholar] [CrossRef]
- Namjou, B.; Stanaway, I.B.; Lingren, T.; Mentch, F.D.; Benoit, B.; Dikilitas, O.; Niu, X.; Shang, N.; Shoemaker, A.H.; Carey, D.J.; et al. Evaluation of the MC4R Gene across eMERGE Network Identifies Many Unreported Obesity-Associated Variants. Int. J. Obes. 2021, 45, 155–169. [Google Scholar] [CrossRef]
- Farooqi, I.S.; Keogh, J.M.; Yeo, G.S.H.; Lank, E.J.; Cheetham, T.; O’Rahilly, S. Clinical Spectrum of Obesity and Mutations in the Melanocortin 4 Receptor Gene. N. Engl. J. Med. 2003, 348, 1085–1095. [Google Scholar] [CrossRef]
- Drabkin, M.; Birk, O.S.; Birk, R. Heterozygous versus Homozygous Phenotype Caused by the Same MC4R Mutation: Novel Mutation Affecting a Large Consanguineous Kindred. BMC Med. Genet. 2018, 19, 135. [Google Scholar] [CrossRef]
- Novoselova, T.V.; Chan, L.F.; Clark, A.J.L. Pathophysiology of Melanocortin Receptors and Their Accessory Proteins. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 93–106. [Google Scholar] [CrossRef]
- Ranadive, S.A.; Vaisse, C. Lessons from Extreme Human Obesity: Monogenic Disorders. Endocrinol. Metab. Clin. N. Am. 2008, 37, 733–751. [Google Scholar] [CrossRef]
- Lotta, L.A.; Mokrosinski, J.; Mendes de Oliveira, E.; Li, C.; Sharp, S.J.; Luan, J.; Brouwers, B.; Ayinampudi, V.; Bowker, N.; Kerrison, N.; et al. Human Gain-of-Function MC4R Variants Show Signaling Bias and Protect against Obesity. Cell 2019, 177, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-Wide Association Yields New Sequence Variants at Seven Loci That Associate with Measures of Obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Willer, C.J.; Speliotes, E.K.; Loos, R.J.F.; Li, S.; Lindgren, C.M.; Heid, I.M.; Berndt, S.I.; Elliott, A.L.; Jackson, A.U.; Lamina, C.; et al. Six New Loci Associated with Body Mass Index Highlight a Neuronal Influence on Body Weight Regulation. Nat. Genet. 2009, 41, 25–34. [Google Scholar] [PubMed]
- Loos, R.J.F.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common Variants near MC4R Are Associated with Fat Mass, Weight and Risk of Obesity. Nat. Genet. 2008, 40, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Meyre, D.; Delplanque, J.; Chèvre, J.-C.; Lecoeur, C.; Lobbens, S.; Gallina, S.; Durand, E.; Vatin, V.; Degraeve, F.; Proença, C.; et al. Genome-Wide Association Study for Early-Onset and Morbid Adult Obesity Identifies Three New Risk Loci in European Populations. Nat. Genet. 2009, 41, 157–159. [Google Scholar] [CrossRef]
- Kring, S.I.I.; Holst, C.; Toubro, S.; Astrup, A.; Hansen, T.; Pedersen, O.; Sørensen, T.I.A. Common Variants near MC4R in Relation to Body Fat, Body Fat Distribution, Metabolic Traits and Energy Expenditure. Int. J. Obes. 2010, 34, 182–189. [Google Scholar] [CrossRef]
- Stutzmann, F.; Cauchi, S.; Durand, E.; Calvacanti-Proença, C.; Pigeyre, M.; Hartikainen, A.-L.; Sovio, U.; Tichet, J.; Marre, M.; Weill, J.; et al. Common Genetic Variation near MC4R Is Associated with Eating Behaviour Patterns in European Populations. Int. J. Obes. 2009, 33, 373–378. [Google Scholar] [CrossRef]
- Dastgheib, S.A.; Bahrami, R.; Setayesh, S.; Salari, S.; Mirjalili, S.R.; Noorishadkam, M.; Sadeghizadeh-Yazdi, J.; Akbarian, E.; Neamatzadeh, H. Evidence from a Meta-Analysis for Association of MC4R Rs17782313 and FTO Rs9939609 Polymorphisms with Susceptibility to Obesity in Children. Diabetes Metab. Syndr. 2021, 15, 102234. [Google Scholar] [CrossRef]
- Hammad, M.M.; Abu-Farha, M.; Hebbar, P.; Cherian, P.; Al Khairi, I.; Melhem, M.; Alkayal, F.; Alsmadi, O.; Thanaraj, T.A.; Al-Mulla, F.; et al. MC4R Variant Rs17782313 Associates with Increased Levels of DNAJC27, Ghrelin, and Visfatin and Correlates with Obesity and Hypertension in a Kuwaiti Cohort. Front. Endocrinol. 2020, 11, 437. [Google Scholar] [CrossRef]
- Tao, L.; Zhang, Z.; Chen, Z.; Zhou, D.; Li, W.; Kan, M.; Zhang, D.; He, L.; Huang, G.; Liu, Y. A Common Variant near the Melanocortin 4 Receptor Is Associated with Low-Density Lipoprotein Cholesterol and Total Cholesterol in the Chinese Han Population. Mol. Biol. Rep. 2012, 39, 6487–6493. [Google Scholar] [CrossRef]
- Illangasekera, Y.A.; Kumarasiri, R.P.V.; Fernando, D.J.; Dalton, C.F. Association of FTO and near MC4R Variants with Obesity Measures in Urban and Rural Dwelling Sri Lankans. Obes. Res. Clin. Pract. 2016, 10 (Suppl. S1), S117–S124. [Google Scholar] [CrossRef] [PubMed]
- Sull, J.W.; Kim, G.; Jee, S.H. Association of MC4R (Rs17782313) with Diabetes and Cardiovascular Disease in Korean Men and Women. BMC Med. Genet. 2020, 21, 160. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.R.; Peter, J.-C.; Lecourt, A.-C.; Barde, Y.-A.; Hofbauer, K.G. Melanocortin-4 Receptor Activation Stimulates Hypothalamic Brain-Derived Neurotrophic Factor Release to Regulate Food Intake, Body Temperature and Cardiovascular Function. J. Neuroendocrinol. 2007, 19, 974–982. [Google Scholar] [CrossRef]
- Purnell, J.Q. Definitions, Classification, and Epidemiology of Obesity. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Rahati, S.; Qorbani, M.; Naghavi, A.; Pishva, H. Association and Interaction of the MC4R Rs17782313 Polymorphism with Plasma Ghrelin, GLP-1, Cortisol, Food Intake and Eating Behaviors in Overweight/Obese Iranian Adults. BMC Endocr. Disord. 2022, 22, 234. [Google Scholar] [CrossRef]
- Farooq, S.; Rana, S.; Siddiqui, A.J.; Iqbal, A.; Musharraf, S.G. Association of Metabolites with Obesity Based on Two Gene Variants, MC4R Rs17782313 and BDNF Rs6265. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166144. [Google Scholar] [CrossRef]
- MacArthur, J.; Bowler, E.; Cerezo, M.; Gil, L.; Hall, P.; Hastings, E.; Junkins, H.; McMahon, A.; Milano, A.; Morales, J.; et al. The New NHGRI-EBI Catalog of Published Genome-Wide Association Studies (GWAS Catalog). Nucleic Acids Res. 2017, 45, D896–D901. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.R.; Lal, N.; Giridharan, N.V. Genetic & Epigenetic Approach to Human Obesity. Indian J. Med. Res. 2014, 140, 589–603. [Google Scholar] [PubMed]
- Bora, J.K.; Saikia, N. Neonatal and Under-Five Mortality Rate in Indian Districts with Reference to Sustainable Development Goal 3: An Analysis of the National Family Health Survey of India (NFHS), 2015–2016. PLoS ONE 2018, 13, e0201125. [Google Scholar] [CrossRef]
- Kshatriya, G.K.; Acharya, S.K. Triple Burden of Obesity, Undernutrition, and Cardiovascular Disease Risk among Indian Tribes. PLoS ONE 2016, 11, e0147934. [Google Scholar] [CrossRef]
- Castro, G.V.; Latorre, A.F.S.; Korndorfer, F.P.; de Carlos Back, L.K.; Lofgren, S.E. The Impact of Variants in Four Genes: MC4R, FTO, PPARG and PPARGC1A in Overweight and Obesity in a Large Sample of the Brazilian Population. Biochem. Genet. 2021, 59, 1666–1679. [Google Scholar] [CrossRef]
- Wei, B.-L.; Yin, R.-X.; Liu, C.-X.; Deng, G.-X.; Guan, Y.-Z.; Zheng, P.-F. The MC4R SNPs, Their Haplotypes and Gene-Environment Interactions on the Risk of Obesity. Mol. Med. 2020, 26, 77. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.C.Y.; Tam, C.H.T.; So, W.Y.; Ho, J.S.K.; Chan, A.W.; Lee, H.M.; Wang, Y.; Lam, V.K.L.; Chan, J.C.N.; Ma, R.C.W. Implication of Genetic Variants Near NEGR1, SEC16B, TMEM18, ETV5/DGKG, GNPDA2, LIN7C/BDNF, MTCH2, BCDIN3D/FAIM2, SH2B1, FTO, MC4R, and KCTD15 with Obesity and Type 2 Diabetes in 7705 Chinese. J. Clin. Endocrinol. Metab. 2010, 95, 2418–2425. [Google Scholar] [CrossRef] [PubMed]
- Brodowski, J.; Szkup, M.; Jurczak, A.; Wieder-Huszla, S.; Brodowska, A.; Laszczyńska, M.; Karakiewicz, B.; Kęcka, K.; Grochans, E. Searching for the Relationship between the Parameters of Metabolic Syndrome and the Rs17782313 (T>C) Polymorphism of the MC4R gene in Postmenopausal Women. Clin. Interv. Aging 2017, 12, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Rahmani, S.; Mirza, S. MC4R Variant Rs17782313 and Manifestation of Obese Phenotype in Pakistani Females. RSC Adv. 2018, 8, 16957–16972. [Google Scholar] [CrossRef] [PubMed]
- Bjørnland, T.; Langaas, M.; Grill, V.; Mostad, I.L. Assessing Gene-Environment Interaction Effects of FTO, MC4R and Lifestyle Factors on Obesity Using an Extreme Phenotype Sampling Design: Results from the HUNT Study. PLoS ONE 2017, 12, e0175071. [Google Scholar] [CrossRef]
- Horstmann, A.; Kovacs, P.; Kabisch, S.; Boettcher, Y.; Schloegl, H.; Tönjes, A.; Stumvoll, M.; Pleger, B.; Villringer, A. Common Genetic Variation near MC4R Has a Sex-Specific Impact on Human Brain Structure and Eating Behavior. PLoS ONE 2013, 8, e74362. [Google Scholar] [CrossRef]
- Qi, L.; Kraft, P.; Hunter, D.J.; Hu, F.B. The Common Obesity Variant near MC4R Gene Is Associated with Higher Intakes of Total Energy and Dietary Fat, Weight Change and Diabetes Risk in Women. Hum. Mol. Genet. 2008, 17, 3502–3508. [Google Scholar] [CrossRef]
- Kapoor, N.; Arora, S.; Kalra, S. Gender Disparities in People Living with Obesity—An Unchartered Territory. J. Mid-Life Health 2021, 12, 103–107. [Google Scholar]
- Yusuf, F.M.; San Sebastián, M.; Vaezghasemi, M. Explaining Gender Inequalities in Overweight People: A Blinder-Oaxaca Decomposition Analysis in Northern Sweden. Int. J. Equity Health 2023, 22, 159. [Google Scholar] [CrossRef]
- Kroll, D.S.; Feldman, D.E.; Biesecker, C.L.; McPherson, K.L.; Manza, P.; Joseph, P.V.; Volkow, N.D.; Wang, G.-J. Neuroimaging of Sex/Gender Differences in Obesity: A Review of Structure, Function, and Neurotransmission. Nutrients 2020, 12, 1942. [Google Scholar] [CrossRef]

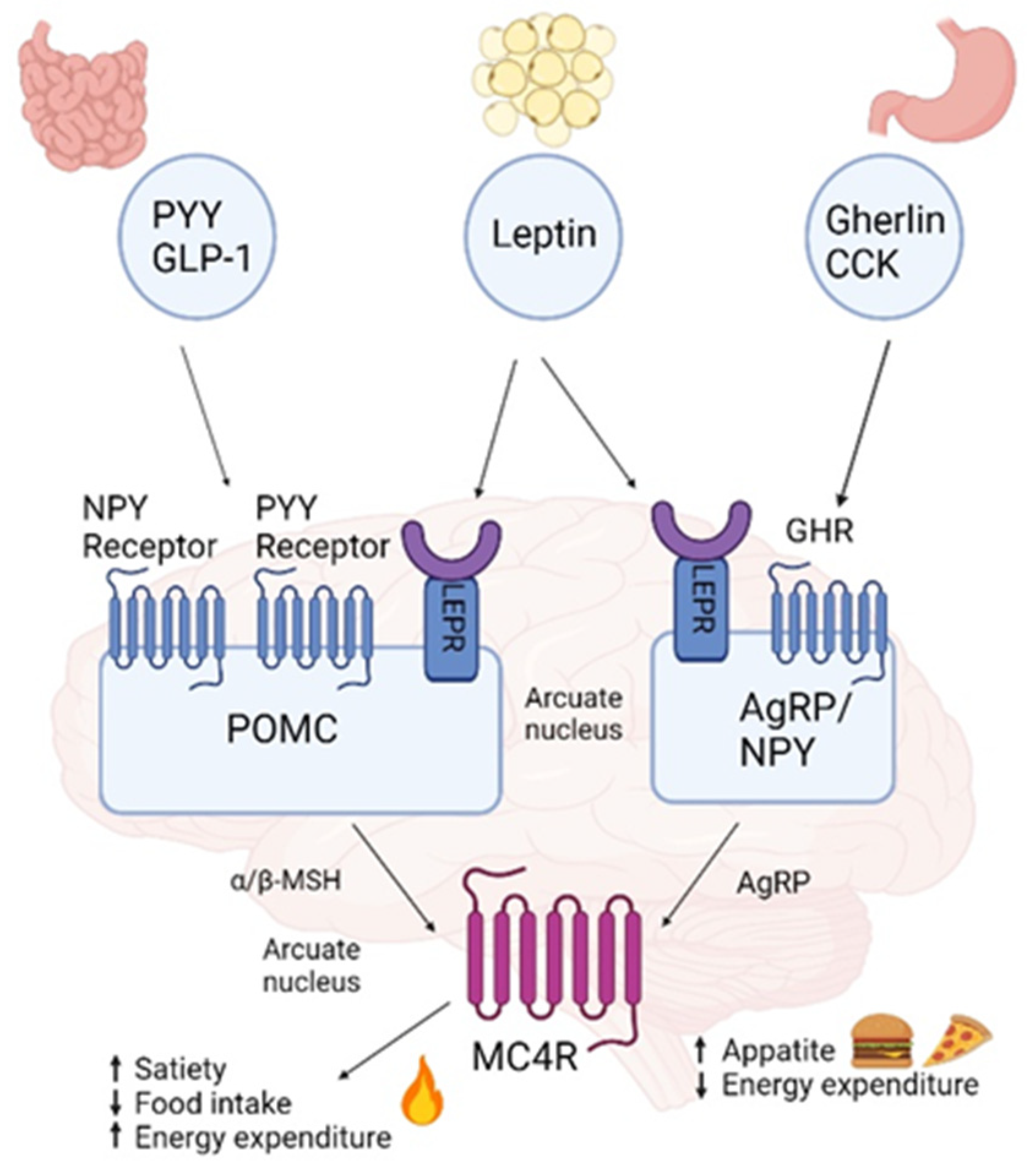{kind=link}
| All Population n = 5785 | With Obesity (BMI ≥ 30) n = 3157 | Non-Obese (BMI < 30) n = 2628 | p-Value | |
|---|---|---|---|---|
| Sex (women, %) | 4023 (69.5%) | 2120 (67.2%) | 1903 (72.4%) | <0.001 |
| Age (mean ± SD) | 56.47 ± 14.48 | 56.61 ± 14.66 | 56.31 ± 14.26 | 0.22 |
| Weight (mean ± SD) | 87.1 ± 19.4 | 98.34 ± 17.7 | 73.61 ± 10.93 | <0.001 |
| Height (mean ± SD) | 166.64 ± 8.85 | 166.69 ± 9.12 | 166.58 ± 8.52 | 0.86 |
| BMI (mean ± SD) | 31.27 ± 6.04 | 35.29 ± 5.05 | 26.44 ± 2.6 | <0.001 |
| T1DM (n, %) | 100 (1.73%) | 62 (1.96%) | 38 (1.45%) | 0.27 |
| T2DM (n, %) | 449 (7.76%) | 284 (9%) | 165 (6.3%) | <0.001 |
| Genotype Frequency (%) | p-Value OR ± 95%(CI) | ||||||
|---|---|---|---|---|---|---|---|
| Sample | Genotype | Overall Population | With Obesity (BMI ≥ 30) | Non-Obese (BMI < 30) | Dominant Model | Recessive Model | Additive Model |
| All sample * | TT | 3375 (58.3%) | 1818 (57.6%) | 1557 (59.2%) | 0.16 1.08 (0.97–1.2) | 0.005 1.38 (1.1–1.72) | 0.03 1.1 (1.01–1.2) |
| TC | 2062 (35.6%) | 1125 (35.6%) | 937 (35.6%) | ||||
| CC | 348 (6%) | 214 (6.8%) | 134 (5.1%) | ||||
| Females (n = 4023) | TT | 2328 (35.6%) | 1211 (57.1%) | 1117 (58.7%) | 0.28 1.07 (0.95–1.22) | 0.01 1.41 (1.09–1.82) | 0.06 1.62 (1.24–2.11) |
| TC | 1434 (35.6%) | 751 (35.4%) | 683 (35.9%) | ||||
| CC | 261 (6.5%) | 158 (7.4%) | 103 (5.4%) | ||||
| Males (n = 1762) | TT | 1047 (59.4%) | 607 (58.5%) | 440 (60.6%) | 0.34 1.1 (0.9–1.34) | 0.28 1.28 (0.82–1.01) | 0.23 1.11 (0.94–1.23) |
| TC | 628 (35.6%) | 374 (36.1%) | 254 (35.0%) | ||||
| CC | 87 (4.9%) | 56 (5.4%) | 31 (4.3%) | ||||
| Eating Behavior | BMI ≥ 30 | BMI < 30 | β | OR ± 95% CI | p-Value |
|---|---|---|---|---|---|
| Sweets desire | 91 (57.6%) | 56 (54.4%) | 0.387 | 1.47 (1.05–2.07) | 0.03 |
| Emotional eater | 83 (52.5%) | 23 (22.3%) | 1.2 | 3.32 (2.08–5.3) | <0.001 |
| Overfullness feeling | 54 (34.2%) | 21 (20.4%) | 0.84 | 2.32 (1.39–3.87) | 0.001 |
| Junk food ≥ 1/week | 26 (16.5%) | 8 (7.8%) | 1.1 | 3 (1.36–6.67) | 0.007 |
| Fast eater | 57 (36.1%) | 32 (31.1%) | 0.495 | 1.64 (1.06–2.54) | 0.027 |
| Late night eater | 60 (38%) | 23 (22.3%) | 0.853 | 2.35 (1.44–3.82) | <0.001 |
| Non-tableside eater | 48 (65.8%) | 25 (34.2%) | 0.541 | 1.72 (1.05–2.8) | 0.03 |
| Non-hunger-driven eater | 75 (47.5%) | 39 (37.9%) | 0.565 | 1.76 (1.19–2.6) | 0.005 |
| Eat while standing | 23 (14.6%) | 10 (9.7%) | 0.736 | 2.09 (1–4.4) | 0.05 |
| Distracted eater | 98 (62.0%) | 54 (52.4%) | 0.519 | 1.68 (1.2–2.36) | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chermon, D.; Birk, R. Predisposition of the Common MC4R rs17782313 Female Carriers to Elevated Obesity and Interaction with Eating Habits. Genes 2023, 14, 1996. https://doi.org/10.3390/genes14111996
Chermon D, Birk R. Predisposition of the Common MC4R rs17782313 Female Carriers to Elevated Obesity and Interaction with Eating Habits. Genes. 2023; 14(11):1996. https://doi.org/10.3390/genes14111996
Chicago/Turabian StyleChermon, Danyel, and Ruth Birk. 2023. "Predisposition of the Common MC4R rs17782313 Female Carriers to Elevated Obesity and Interaction with Eating Habits" Genes 14, no. 11: 1996. https://doi.org/10.3390/genes14111996
APA StyleChermon, D., & Birk, R. (2023). Predisposition of the Common MC4R rs17782313 Female Carriers to Elevated Obesity and Interaction with Eating Habits. Genes, 14(11), 1996. https://doi.org/10.3390/genes14111996