
Genome-Wide Identification and Characterization of CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Foxtail Millet (Setaria italica L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of SiCLE Genes in Foxtail Millet
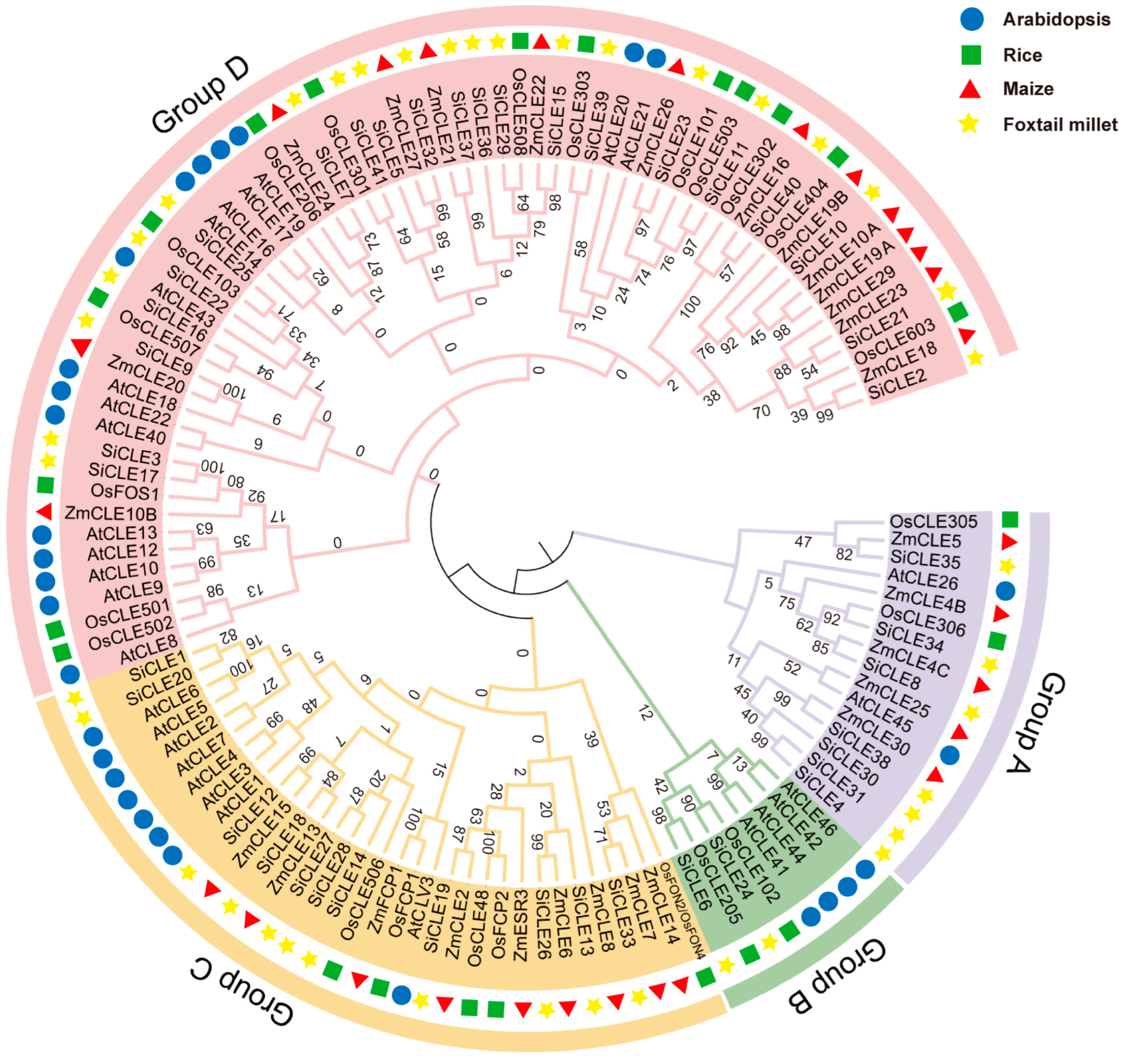
2.2. Phylogenetic Analysis and Classification of SiCLE Genes in Foxtail Millet
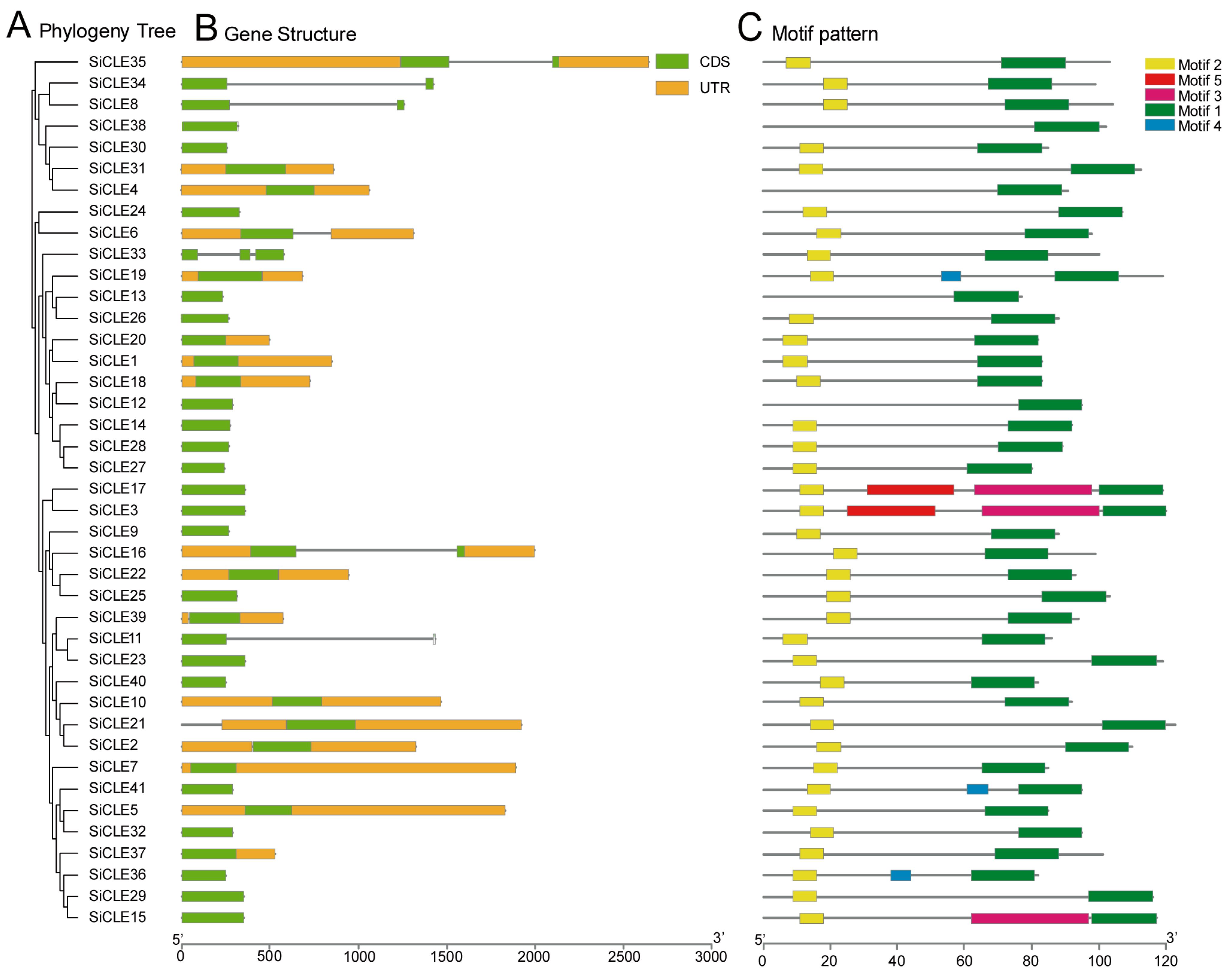
2.3. Gene Structure and Conserved Motif Analysis of SiCLE Genes in Foxtail Millet
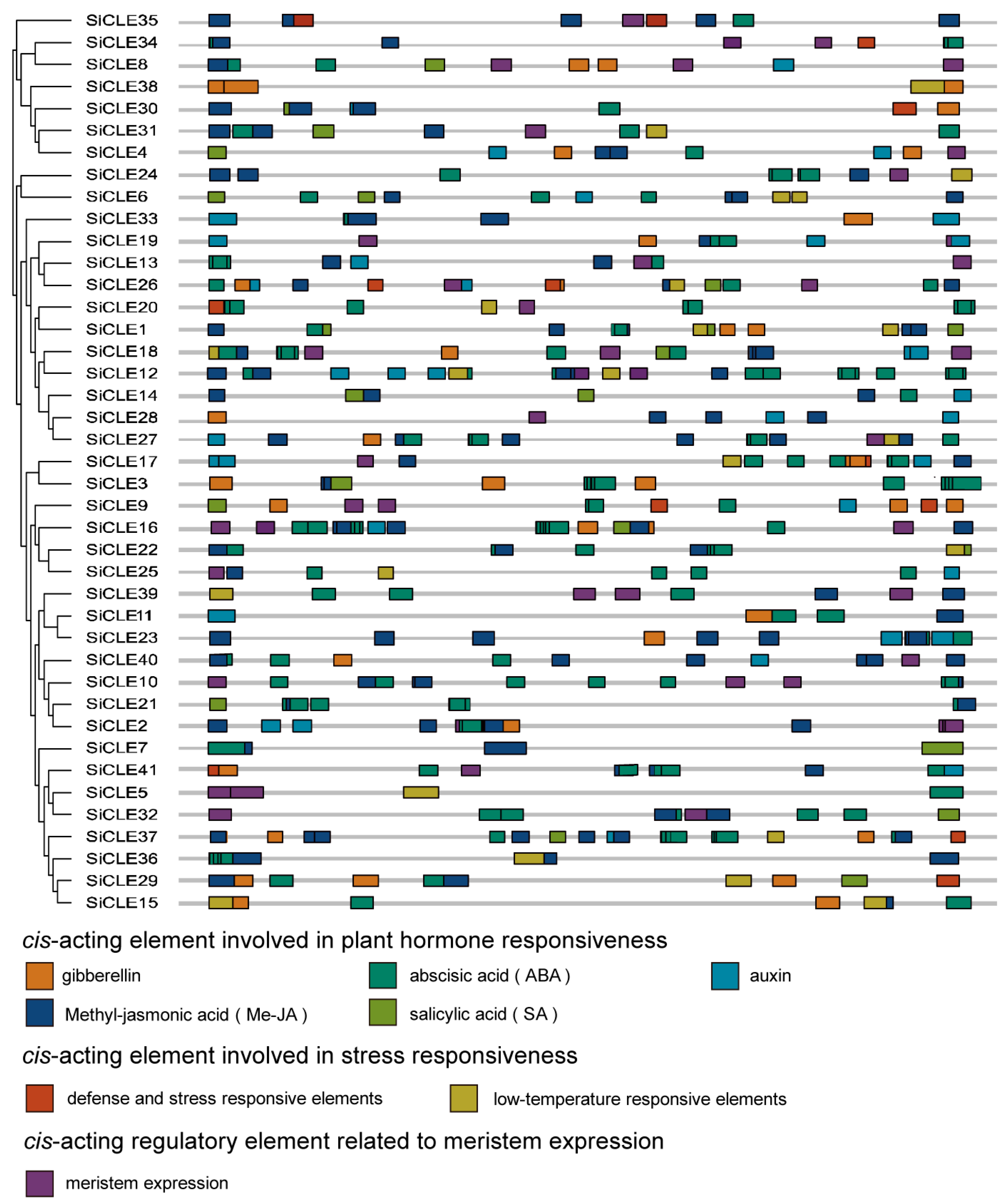
2.4. Cis-Regulatory Elements Analysis of SiCLE Genes in Foxtail Millet
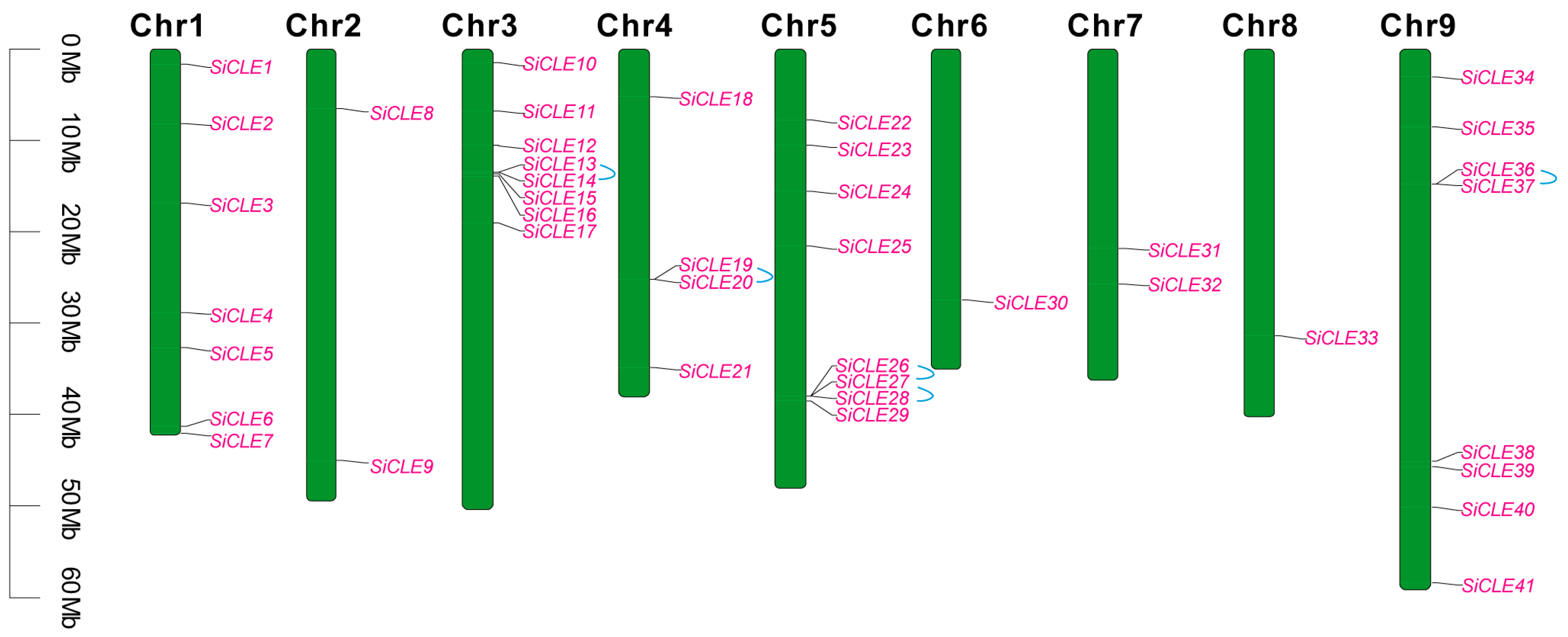
2.5. Cene Duplication and Collinearity Analysis of the SiCLEs in Foxtail Millet
2.6. Expression Pattern Analysis of SiCLE Genes in Foxtail Millet
2.7. Response of SiCLE Genes to Phytohormones in Foxtail Millet
3. Results
3.1. Identification of the SiCLE Genes in Foxtail Millet
3.2. Phylogenetic Analysis and Classification of SiCLE Peptides in Foxtail Millet
3.3. Gene Structure and Conserved Motif Analysis of SiCLE Peptides in Foxtail Millet
3.4. Cis-Regulatory Elements in SiCLE Gene Promoters
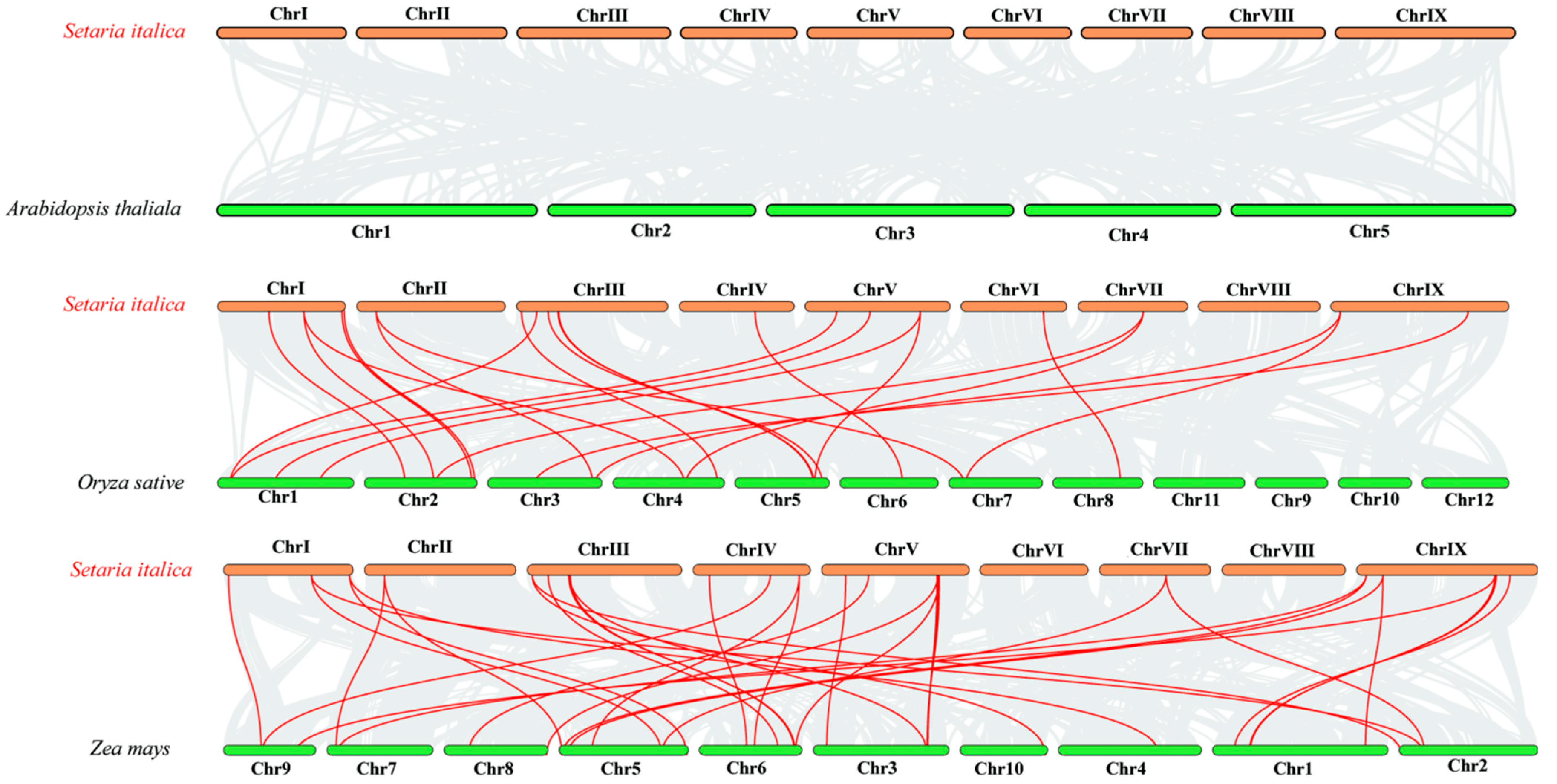
3.5. Chromosomal Distribution, Gene Duplication, and Synteny Analysis of the SiCLE Genes
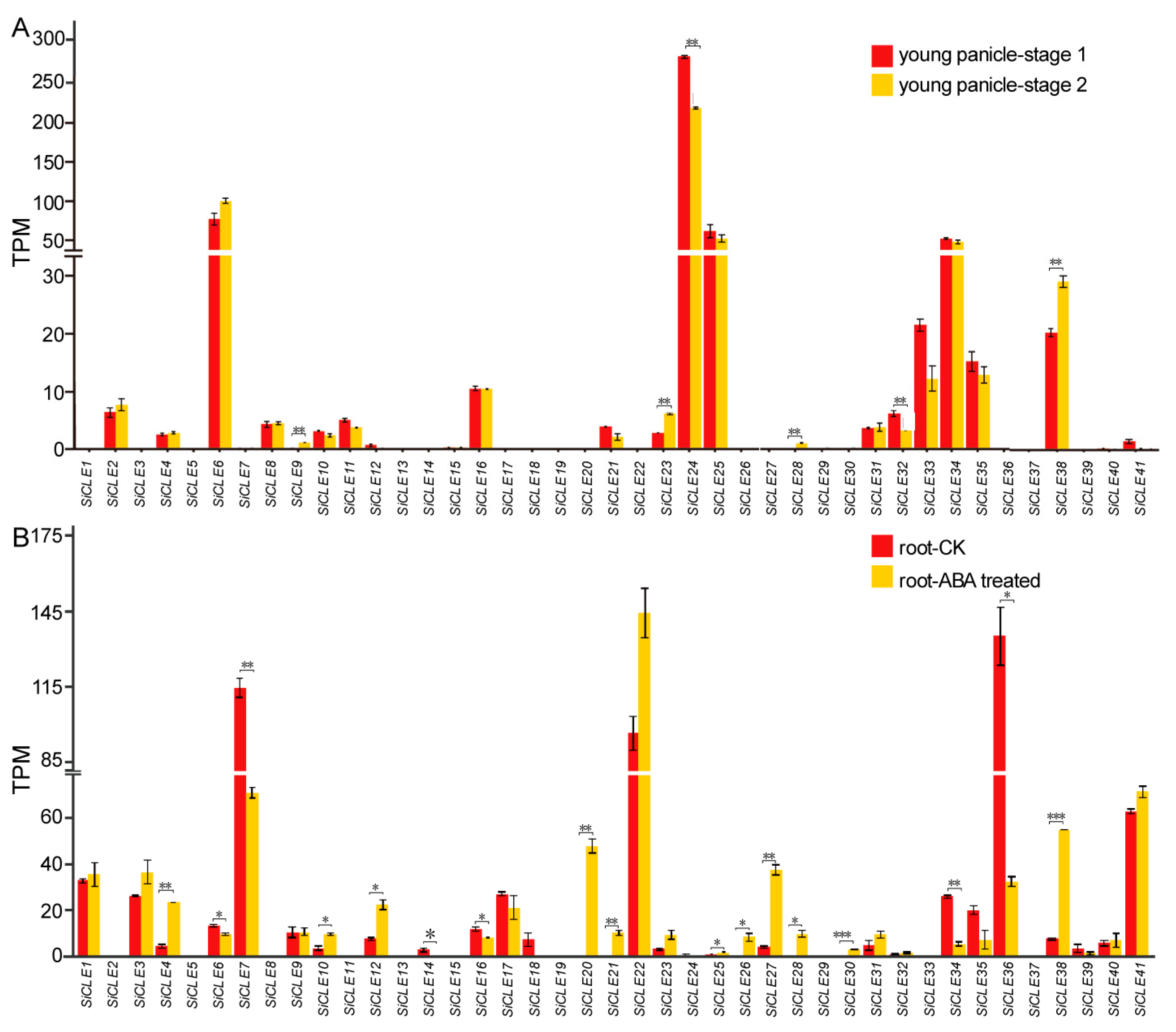
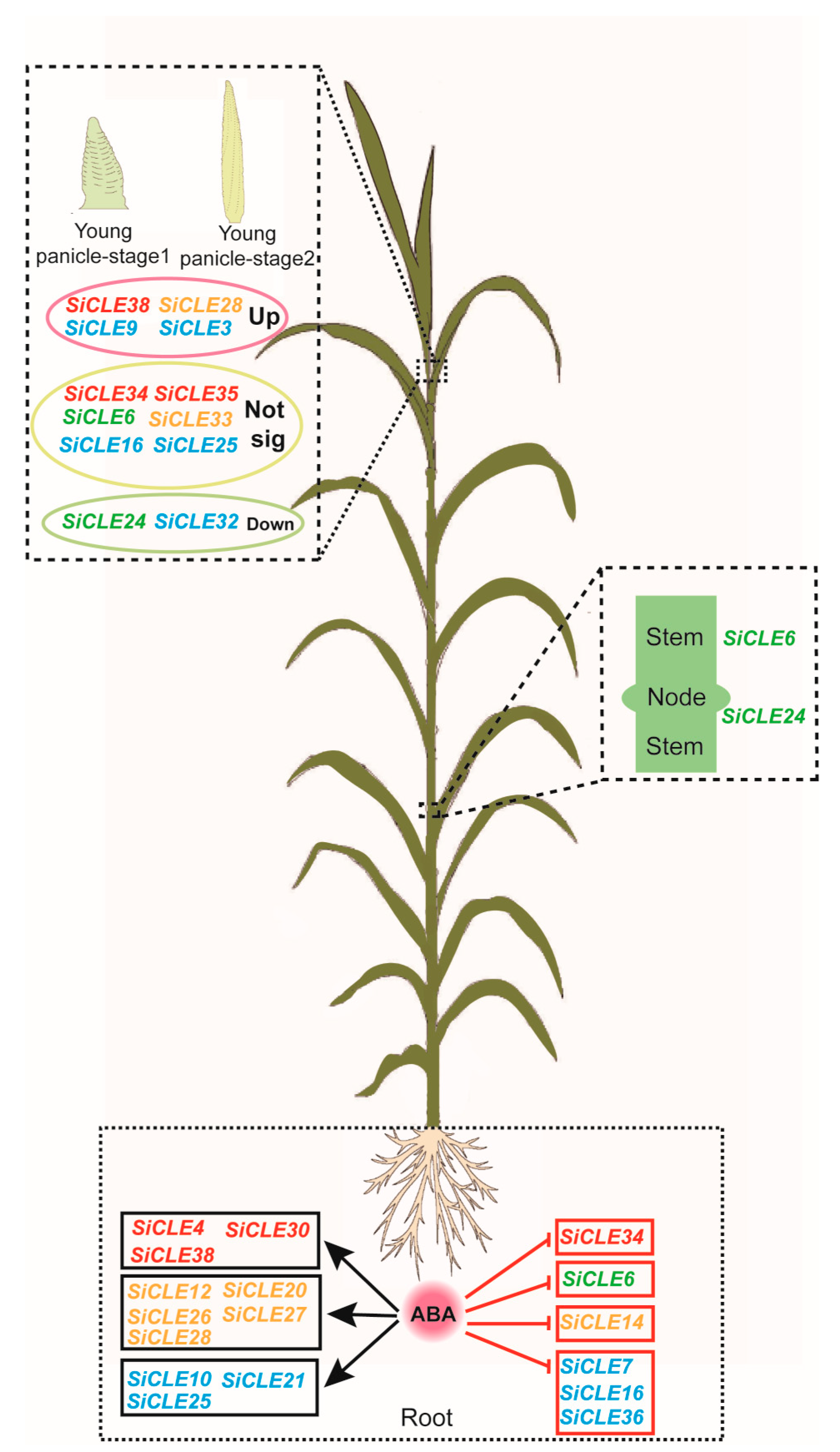
3.6. Expression Pattern Analysis of SiCLE Genes in Different Tissues
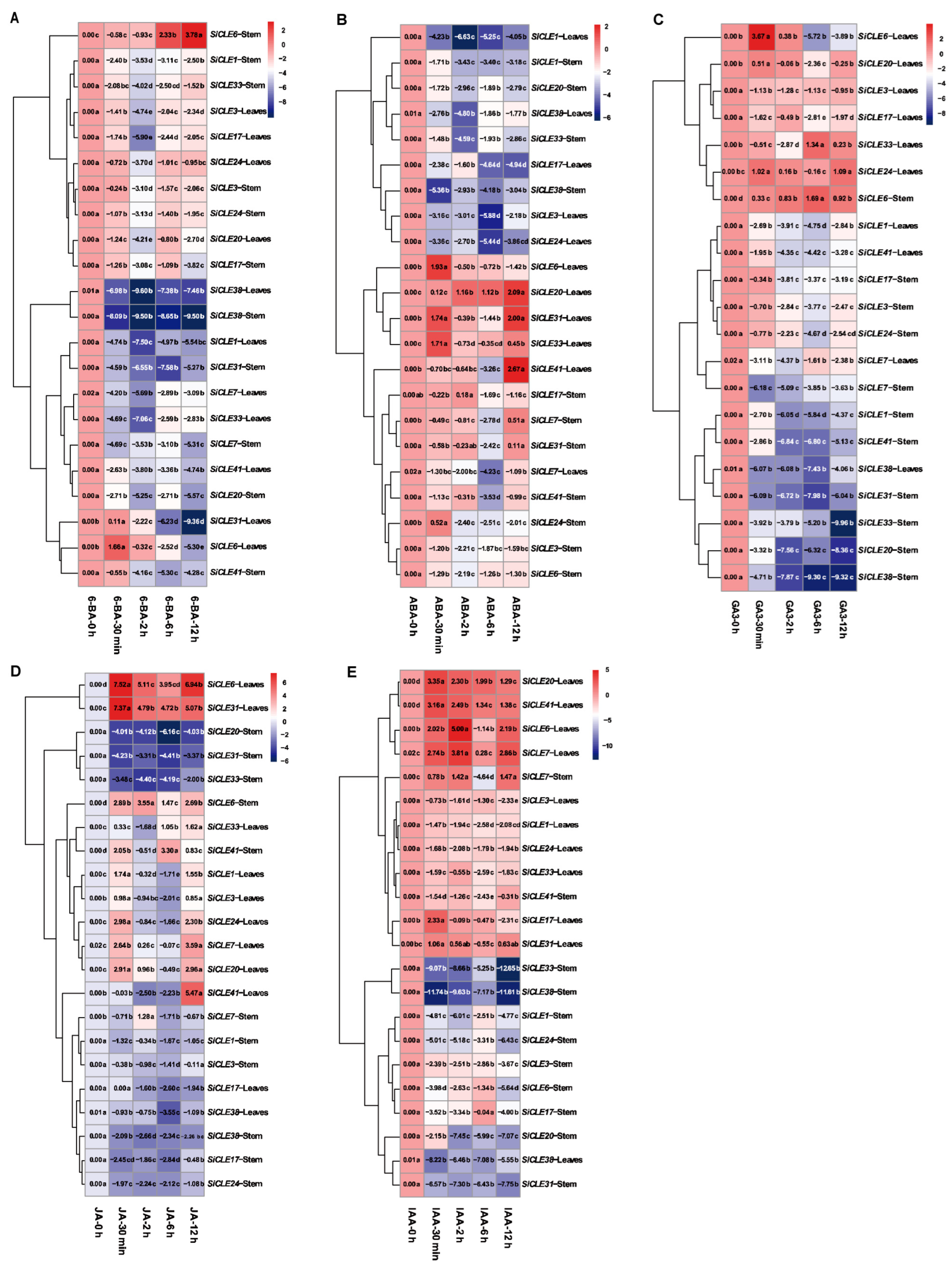
3.7. Response of SiCLEs to Plant Hormones in Foxtail Millet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fletcher, J.C. Recent Advances in Arabidopsis CLE Peptide Signaling. Trends Plant Sci. 2020, 25, 1005–1016. [Google Scholar] [PubMed]
- Tavormina, P.; De Coninck, B.; Nikonorova, N.; De Smet, I.; Cammue, B.P. The Plant Peptidome: An Expanding Repertoire of Structural Features and Biological Functions. Plant Cell 2015, 27, 2095–2118. [Google Scholar] [PubMed]
- Matsubayashi, Y. Small post-translationally modified Peptide signals in Arabidopsis. Arab. Book 2011, 9, e0150. [Google Scholar]
- Betsuyaku, S.; Sawa, S.; Yamada, M. The function of the CLE peptides in plant development and plant-microbe interactions. Arab. Book 2011, 9, e0149. [Google Scholar]
- Rodriguez-Garcia, A.; Smertenko, A. CLE peptides: Emerging players in plant development and stress responses. J. Exp. Bot. 2022, 73, 3509–3524. [Google Scholar]
- Fiume, E.; Fletcher, J.C.; Bergmann, D.C. The CLE signaling peptides regulate diverse aspects of plant development. Curr. Opin. Plant Biol. 2021, 63, 102049. [Google Scholar]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 2000, 289, 617–619. [Google Scholar]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.; Jürgens, G.; Laux, T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar]
- Chu, H.; Qian, Q.; Liang, W.; Yin, C.; Tan, H.; Yao, X.; Yuan, Z.; Yang, J.; Huang, H.; Luo, D.; et al. The FLORAL ORGAN NUMBER4 gene encoding a putative ortholog of Arabidopsis CLAVATA3 regulates apical meristem size in rice. Plant Physiol. 2006, 142, 1039–1052. [Google Scholar]
- Ohmori, Y.; Tanaka, W.; Kojima, M.; Sakakibara, H.; Hirano, H.Y. WUSCHEL-RELATED HOMEOBOX4 is involved in meristem maintenance and is negatively regulated by the CLE gene FCP1 in rice. Plant Cell 2013, 25, 229–241. [Google Scholar] [PubMed]
- Suzaki, T.; Sato, M.; Ashikari, M.; Miyoshi, M.; Nagato, Y.; Hirano, H.Y. The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis CLAVATA1. Development 2004, 131, 5649–5657. [Google Scholar] [PubMed]
- Bommert, P.; Lunde, C.; Nardmann, J.; Vollbrecht, E.; Running, M.; Jackson, D.; Hake, S.; Werr, W. Thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptor-like kinase. Development 2005, 132, 1235–1245. [Google Scholar]
- Xu, C.; Liberatore, K.L.; MacAlister, C.A.; Huang, Z.; Chu, Y.H.; Jiang, K.; Brooks, C.; Ogawa-Ohnishi, M.; Xiong, G.; Pauly, M.; et al. A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat. Genet. 2015, 47, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Taguchi-Shiobara, F.; Yuan, Z.; Hake, S.; Jackson, D. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Gene Dev. 2001, 15, 2755–2766. [Google Scholar] [PubMed]
- Zhang, Y.; Tan, S.; Gao, Y.; Kan, C.; Wang, H.L.; Yang, Q.; Xia, X.; Ishida, T.; Sawa, S.; Guo, H.; et al. CLE42 delays leaf senescence by antagonizing ethylene pathway in Arabidopsis. New Phytol. 2022, 235, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Cammarata, J.; Roeder, A.H.; Scanlon, M.J. Cytokinin and CLE signaling are highly intertwined developmental regulators across tissues and species. Curr. Opin. Plant Biol. 2019, 51, 96–104. [Google Scholar]
- Bidadi, H.; Matsuoka, K.; Sage-Ono, K.; Fukushima, J.; Pitaksaringkarn, W.; Asahina, M.; Yamaguchi, S.; Sawa, S.; Fukuda, H.; Matsubayashi, Y.; et al. CLE6 expression recovers gibberellin deficiency to promote shoot growth in Arabidopsis. Plant J. 2014, 78, 241–252. [Google Scholar]
- Zhang, L.; Shi, X.; Zhang, Y.; Wang, J.; Yang, J.; Ishida, T.; Jiang, W.; Han, X.; Kang, J.; Wang, X.; et al. CLE9 peptide-induced stomatal closure is mediated by abscisic acid, hydrogen peroxide, and nitric oxide in Arabidopsis thaliana. Plant Cell Environ. 2019, 42, 1033–1044. [Google Scholar]
- DiGennaro, P.; Grienenberger, E.; Dao, T.Q.; Jun, J.H.; Fletcher, J.C. Peptide signaling molecules CLE5 and CLE6 affect Arabidopsis leaf shape downstream of leaf patterning transcription factors and auxin. Plant Direct 2018, 2, e00103. [Google Scholar]
- Zhang, Y.; Yang, S.; Song, Y.; Wang, J. Genome-wide characterization, expression and functional analysis of CLV3/ESR gene family in tomato. BMC Genom. 2014, 15, 827. [Google Scholar] [CrossRef]
- Carbonnel, S.; Falquet, L.; Hazak, O. Deeper genomic insights into tomato CLE genes repertoire identify new active peptides. BMC Genom. 2022, 23, 756. [Google Scholar] [CrossRef] [PubMed]
- Wulf, K.; Sun, J.; Wang, C.; Ho-Plagaro, T.; Kwon, C.T.; Velandia, K.; Correa-Lozano, A.; Tamayo-Navarrete, M.I.; Reid, J.B.; García Garrido, J.M.; et al. The role of CLE peptides in suppression of mycorrhizal colonisation of tomato. Plant Cell Physiol. 2023, pcad124. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Gao, Y.; Cheng, X.; Yang, Y.; Wu, J.; Wang, J.; Li, S.; Xing, G. Genome-wide identification of CLE gene family and their potential roles in bolting and fruit bearing in cucumber (Cucumis sativus L.). BMC Plant Biol. 2021, 21, 143. [Google Scholar] [CrossRef]
- Hastwell, A.H.; Gresshoff, P.M.; Ferguson, B.J. Genome-wide annotation and characterization of CLAVATA/ESR (CLE) peptide hormones of soybean (Glycine max) and common bean (Phaseolus vulgaris), and their orthologues of Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 5271–5287. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Zhang, G.; Wu, M.; Wang, G. Identification and characterization of the Populus trichocarpa CLE family. BMC Genom. 2016, 17, 174. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Y.; Ren, F. Genome-wide identification of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) family in grape (Vitis vinifera L.). BMC Genom. 2019, 20, 553. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, D.; Xia, Y.; Li, Z.; Niu, N.; Ma, S.; Wang, J.; Song, Y.; Zhang, G. Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Wheat. Int. J. Mol. Sci. 2019, 20, 4319. [Google Scholar] [CrossRef]
- Han, S.; Khan, M.H.U.; Yang, Y.; Zhu, K.; Li, H.; Zhu, M.; Amoo, O.; Khan, S.U.; Fan, C.; Zhou, Y. Identification and comprehensive analysis of the CLV3/ESR-related (CLE) gene family in Brassica napus L. Plant Biol. 2020, 22, 709–721. [Google Scholar] [CrossRef]
- Wan, K.; Lu, K.; Gao, M.; Zhao, T.; He, Y.; Yang, D.L.; Tao, X.; Xiong, G.; Guan, X. Functional analysis of the cotton CLE polypeptide signaling gene family in plant growth and development. Sci. Rep. 2021, 11, 5060. [Google Scholar] [CrossRef]
- Gancheva, M.S.; Lutova, L.A. Nitrogen-Activated CLV3/ESR-Related 4 (CLE4) Regulates Shoot, Root, and Stolon Growth in Potato. Plants 2023, 12, 3468. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C4 model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hou, S.; Feng, M.; Xia, R.; Li, J.; Tang, S.; Han, Y.; Gao, J.; Wang, X. MDSi: Multi-omics Database for Setaria italica. BMC Plant Biol. 2023, 23, 223. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Oelkers, K.; Goffard, N.; Weiller, G.F.; Gresshoff, P.M.; Mathesius, U.; Frickey, T. Bioinformatic analysis of the CLE signaling peptide family. BMC Plant Biol. 2008, 8, 1. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Rozewicki, J.; Li, S.L.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated protein sequence and structural alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Hereditas 2007, 29, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Cheng, L.; Wang, J.; Liu, J.; Cheng, J.; Yang, Z.; Cao, R.; Han, Y.; Li, H.; Zhang, B. Carotenoid Cleavage Dioxygenase 1 Catalyzes Lutein Degradation To Influence Carotenoid Accumulation and Color Development in Foxtail Millet Grains. J. Agric. Food Chem. 2022, 70, 9283–9294. [Google Scholar] [CrossRef] [PubMed]
- Shalabi, L.A.; Shaaban, Z.; Kasasbeh, B. Data mining: A preprocessing engine. J. Comput. Sci 2006, 2, 735–739. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Khan, S.U.; Khan, M.H.U.; Ahmar, S.; Fan, C. Comprehensive study and multipurpose role of the CLV3/ESR-related (CLE) genes family in plant growth and development. J. Cell. Physiol. 2021, 236, 2298–2317. [Google Scholar] [CrossRef]
- Chu, H.; Liang, W.; Li, J.; Hong, F.; Wu, Y.; Wang, L.; Wang, J.; Wu, P.; Liu, C.; Zhang, Q.; et al. A CLE-WOX signalling module regulates root meristem maintenance and vascular tissue development in rice. J. Exp. Bot. 2013, 64, 5359–5369. [Google Scholar] [CrossRef]
- Je, B.I.; Xu, F.; Wu, Q.; Liu, L.; Meeley, R.; Gallagher, J.P.; Corcilius, L.; Payne, R.J.; Bartlett, M.E.; Jackson, D. The CLAVATA receptor FASCIATED EAR2 responds to distinct CLE peptides by signaling through two downstream effectors. eLife 2018, 7, e35673. [Google Scholar] [CrossRef]
- Jun, J.; Fiume, E.; Roeder, A.H.; Meng, L.; Sharma, V.K.; Osmont, K.S.; Baker, C.; Ha, C.M.; Meyerowitz, E.M.; Feldman, L.J.; et al. Comprehensive analysis of CLE polypeptide signaling gene expression and overexpression activity in Arabidopsis. Plant Physiol. 2010, 154, 1721–1736. [Google Scholar] [CrossRef]
- Kinoshita, A.; Nakamura, Y.; Sasaki, E.; Kyozuka, J.; Fukuda, H.; Sawa, S. Gain-of-function phenotypes of chemically synthetic CLAVATA3/ESR-related (CLE) peptides in Arabidopsis thaliana and Oryza sativa. Plant Cell Physiol 2007, 48, 1821–1825. [Google Scholar] [CrossRef] [PubMed]
- Qiang, Y.; Wu, J.; Han, H.; Wang, G. CLE peptides in vascular development. J. Integr. Plant Biol. 2013, 55, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Richardson, A.O. The evolution of spliceosomal introns. Curr. Opin. Genet. Dev. 2002, 12, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Patel, N.; Corcilius, L.; Payne, R.J.; Djordjevic, M.A. CLE peptide tri-arabinosylation and peptide domain sequence composition are essential for SUNN-dependent autoregulation of nodulation in Medicago truncatula. New Phytol. 2018, 218, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 140, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Khong, G.; Richaud, F.; Coudert, Y.; Pati, P.K.; Santi, C.; Périn, C.; Breitler, J.C.; Meynard, D.; Vinh, D.N.; Guiderdoni, E.; et al. Modulating rice stress tolerance by transcription factors. Biotechnol. Genet. Eng. Rev. 2008, 25, 381–404. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.S.; Wilhelm, K.S.; Thomashow, M.F. The 5′-region of Arabidopsis thaliana cor15a has cis-acting elements that confer cold-, drought-and ABA-regulated gene expression. Plant Mol. Biol. 1994, 24, 701–713. [Google Scholar] [CrossRef]
- Casneuf, T.; De Bodt, S.; Raes, J.; Maere, S.; Van de Peer, Y. Nonrandom divergence of gene expression following gene and genome duplications in the flowering plant Arabidopsis thaliana. Genome Biol. 2006, 7, R13. [Google Scholar] [CrossRef]
- Song, X.F.; Hou, X.L.; Liu, C.M. CLE peptides: Critical regulators for stem cell maintenance in plants. Planta 2021, 255, 5. [Google Scholar] [CrossRef]
- De Smet, I.; Signora, L.; Beeckman, T.; Inzé, D.; Foyer, C.H.; Zhang, H. An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. Plant J. 2003, 33, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Fernie, A.R.; Shinozaki, K.; Takahashi, F. Long-distance stress and developmental signals associated with abscisic acid signaling in environmental responses. Plant J. 2021, 105, 477–488. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.; Cai, X.; Han, Z.; Si, J.; Chen, D. Jasmonate Signaling Pathway Modulates Plant Defense, Growth, and Their Trade-Offs. Int. J. Mol. Sci. 2022, 23, 3945. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiao, Y. Cell signaling in the shoot apical meristem. Plant Physiol. 2023, 193, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Awale, P.; McSteen, P. Hormonal regulation of inflorescence and intercalary meristems in grasses. Curr. Opin. Plant Biol. 2023, 102451. [Google Scholar] [CrossRef] [PubMed]
- Selby, R.; Jones, D.S. Complex peptide hormone signaling in plant stem cells. Curr. Opin. Plant Biol. 2023, 75, 102442. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [PubMed]
- John, A.; Smith, E.S.; Jones, D.S.; Soyars, C.L.; Nimchuk, Z.L. A network of CLAVATA receptors buffers auxin-dependent meristem maintenance. Nat. Plants 2023, 9, 1306–1317. [Google Scholar] [CrossRef]
- Goad, D.M.; Zhu, C.; Kellogg, E.A. Comprehensive identification and clustering of CLV3/ESR-related (CLE) genes in plants finds groups with potentially shared function. New Phytol. 2016, 216, 605–616. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Uchida, N.; Yamaguchi, Y.L.; Tabata, R.; Ishida, S.; Ishizaki, K.; Nishihama, R.; Kohchi, T.; Sawa, S.; Bowman, J.L. Control of proliferation in the haploid meristem by CLE peptide signaling in Marchantia polymorpha. PLoS Genet. 2019, 15, e1007997. [Google Scholar] [CrossRef]
- Somssich, M.; Je, B.I.; Simon, R.; Jackson, D. CLAVATA–WUSCHEL signalling in the shoot meristem. Development 2016, 143, 3238–3248. [Google Scholar] [CrossRef] [PubMed]
- Cock, J.M.; McCormick, S. A large family of genes that share homology with CLAVATA3. Plant Physiol. 2001, 126, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Ruth, K.C.; Fletcher, J.C.; Feldman, L. The roles of different CLE domains in Arabidopsis CLE polypeptide activity and functional specificity. Mol. Plant 2010, 3, 760–772. [Google Scholar] [CrossRef]
- Miyawaki, K.; Tabata, R.; Sawa, S. Evolutionarily conserved CLE peptide signaling in plant development, symbiosis, and parasitism. Curr. Opin. Plant Biol. 2013, 16, 598–606. [Google Scholar] [CrossRef]
- Ni, J.; Clark, S.E. Evidence for functional conservation, sufficiency, and proteolytic processing of the CLAVATA3 CLE domain. Plant Physiol. 2006, 140, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Sawa, S.; Kinoshita, A.; Mizuno, S.; Kakimoto, T.; Fukuda, H.; Sakagami, Y. A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science 2006, 313, 845–848. [Google Scholar] [CrossRef]
- Ohyama, K.; Shinohara, H.; Ogawa-Ohnishi, M.; Matsubayashi, Y. A glycopeptide regulating stem cell fate in Arabidopsis thaliana. Nat. Chem. Biol. 2009, 5, 578–580. [Google Scholar] [CrossRef]
- Whitford, R.; Fernandez, A.; De Groodt, R.; Ortega, E.; Hilson, P. Plant CLE peptides from two distinct functional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA 2008, 105, 18625–18630. [Google Scholar] [CrossRef]
- Yamaguchi, Y.L.; Ishida, T.; Sawa, S. CLE peptides and their signaling pathways in plant development. J. Exp. Bot. 2016, 67, 4813–4826. [Google Scholar] [CrossRef]
- Li, S.; Pan, Y.; Wen, C.; Li, Y.; Liu, X.; Zhang, X.; Behera, T.K.; Xing, G.; Weng, Y. Integrated analysis in bi-parental and natural populations reveals CsCLAVATA3 (CsCLV3) underlying carpel number variations in cucumber. Theor. Appl. Genet. 2016, 129, 1007–1022. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, L.; Crowell, O.; Zhao, H.; Brutnell, T.P.; Jackson, D.; Kellogg, E.A. The CLV3 Homolog in Setaria viridis Selectively Controls Inflorescence Meristem Size. Front. Plant Sci. 2021, 12, 636749. [Google Scholar] [CrossRef] [PubMed]
- Yuste-Lisbona, F.J.; Fernández-Lozano, A.; Pineda, B.; Bretones, S.; Ortíz-Atienza, A.; García-Sogo, B.; Müller, N.A.; Angosto, T.; Capel, J.; Moreno, V.; et al. ENO regulates tomato fruit size through the floral meristem development network. Proc. Natl. Acad. Sci. USA 2020, 117, 8187–8195. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wu, Y.; Yang, Q.; Yang, Y.; Meng, Q.; Zhang, K.; Li, J.; Wang, J.; Zhou, Y. A novel single-nucleotide mutation in a CLAVATA3 gene homolog controls a multilocular silique trait in Brassica rapa L. Mol. Plant 2014, 7, 1788–1792. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, K.; Li, H.; Han, S.; Meng, Q.; Khan, S.U.; Fan, C.; Xie, K.; Zhou, Y. Precise editing of CLAVATA genes in Brassica napus L. regulates multilocular silique development. Plant Biotechnol. J. 2018, 16, 1322–1335. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Chromosome Location | Signal Peptide Prediction | Number of Amino Acids | Molecular Weight (kD) | Isoelectric Poin | GRAVY |
|---|---|---|---|---|---|---|---|
| Si1g02740 | SiCLE1 | Chr1:1701179–1702028 | + | 83 | 8.67 | 5.45 | 0.071 |
| Si1g09260 | SiCLE2 | Chr1:8075455–8076779 | + | 110 | 10.98 | 10.87 | −0.06 |
| Si1g15040 | SiCLE3 | Chr1:16870079–16870441 | + | 120 | 13.42 | 11.20 | −0.497 |
| Si1g21450 | SiCLE4 | Chr1:28806486–28807552 | + | 91 | 9.66 | 10.39 | −0.204 |
| Si1g25600 | SiCLE5 | Chr1:32729889–32731715 | + | 85 | 8.68 | 9.49 | 0.232 |
| Si1g36220 | SiCLE6 | Chr1:41284255–41285566 | − | 98 | 10.46 | 11.83 | −0.427 |
| Si1g37440 | SiCLE7 | Chr1:42036658–42038549 | + | 85 | 9.70 | 9.99 | −0.359 |
| Si2g07840 | SiCLE8 | Chr2:6541423–6542683 | − | 104 | 10.92 | 11.49 | −0.34 |
| Si2g37520 | SiCLE9 | Chr2:45004975–45005241 | + | 88 | 9.12 | 6.03 | −0.016 |
| Si3g03240 | SiCLE10 | Chr3:1462866–1464336 | + | 92 | 9.50 | 11.01 | −0.049 |
| Si3g10460 | SiCLE11 | Chr3:6732083–6733514 | + | 86 | 9.26 | 11.83 | −0.2 |
| Si3g14760 | SiCLE12 | Chr3:10562885–10563172 | + | 95 | 9.64 | 8.25 | 0.082 |
| Si3g17810 | SiCLE13 | Chr3:13430959–13431192 | + | 77 | 8.02 | 9.18 | 0.112 |
| Si3g17830 | SiCLE14 | Chr3:13472475–13472753 | + | 92 | 9.27 | 5.68 | 0 |
| Si3g18030 | SiCLE15 | Chr3:13627896–13628249 | + | 117 | 12.66 | 11.25 | −0.368 |
| Si3g18460 | SiCLE16 | Chr3:13936956–13938953 | − | 99 | 10.24 | 11.34 | −0.269 |
| Si3g22990 | SiCLE17 | Chr3:19091857–19092216 | + | 119 | 13.17 | 11.50 | −0.403 |
| Si4g07460 | SiCLE18 | Chr4:5221231–5221955 | + | 83 | 8.54 | 7.78 | 0.094 |
| Si4g16140 | SiCLE19 | Chr4:25203756–25204438 | + | 119 | 12.22 | 9.98 | −0.317 |
| Si4g16170 | SiCLE20 | Chr4:25243962–25244464 | + | 82 | 8.56 | 5.98 | 0.126 |
| Si4g23620 | SiCLE21 | Chr4:34831659–34833580 | + | 127 | 12.86 | 11.03 | −0.366 |
| Si5g09450 | SiCLE22 | Chr5:7782941–7783884 | + | 93 | 9.72 | 11.86 | 0.103 |
| Si5g12400 | SiCLE23 | Chr5:10508454–10508813 | + | 119 | 12.50 | 11.89 | −0.3 |
| Si5g16690 | SiCLE24 | Chr5:15497255–15497578 | + | 107 | 10.95 | 11.91 | 0.068 |
| Si5g18600 | SiCLE25 | Chr5:21570492–21570803 | + | 103 | 11.00 | 12.16 | −0.194 |
| Si5g32640 | SiCLE26 | Chr5:37962048–37962314 | + | 88 | 8.93 | 11.65 | −0.086 |
| Si5g32660 | SiCLE27 | Chr5:37979626–37979868 | + | 80 | 7.93 | 6.57 | 0.173 |
| Si5g32670 | SiCLE28 | Chr5:37981960–37982229 | + | 89 | 9.14 | 9.10 | −0.128 |
| Si5g33310 | SiCLE29 | Chr5:38510416–38510766 | + | 116 | 12.87 | 9.75 | −0.422 |
| Si6g16350 | SiCLE30 | Chr6:27510936–27511193 | + | 85 | 9.61 | 11.42 | −0.351 |
| Si7g13900 | SiCLE31 | Chr7:21729033–21729898 | + | 113 | 11.40 | 11.66 | −0.113 |
| Si7g19100 | SiCLE32 | Chr7:25632473–25632760 | + | 95 | 10.07 | 10.74 | −0.065 |
| Si8g17400 | SiCLE33 | Chr8:31420418–31420996 | + | 100 | 10.33 | 8.97 | −0.068 |
| Si9g05550 | SiCLE34 | Chr9:3007939–3009361 | − | 99 | 10.28 | 11.34 | −0.362 |
| Si9g13150 | SiCLE35 | Chr9:8512203–8514840 | + | 103 | 10.46 | 10.24 | −0.087 |
| Si9g19660 | SiCLE36 | Chr9:14748515–14748763 | + | 82 | 9.10 | 6.56 | −0.402 |
| Si9g19670 | SiCLE37 | Chr9:14751454–14751981 | + | 101 | 11.09 | 10.22 | 0.02 |
| Si9g38030 | SiCLE38 | Chr9:45088642–45088950 | + | 102 | 10.44 | 11.11 | 0.047 |
| Si9g38660 | SiCLE39 | Chr9:45679290–45679866 | + | 94 | 10.38 | 9.81 | −0.068 |
| Si9g44430 | SiCLE40 | Chr9:50178960–50179208 | − | 82 | 8.53 | 11.61 | 0.263 |
| Si9g55410 | SiCLE41 | Chr9:58385979–58386266 | + | 95 | 10.84 | 10.00 | −0.287 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Chen, J.; Chen, S.; Zhang, H.; Li, L. Genome-Wide Identification and Characterization of CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Foxtail Millet (Setaria italica L.). Genes 2023, 14, 2046. https://doi.org/10.3390/genes14112046
Ren X, Chen J, Chen S, Zhang H, Li L. Genome-Wide Identification and Characterization of CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Foxtail Millet (Setaria italica L.). Genes. 2023; 14(11):2046. https://doi.org/10.3390/genes14112046
Chicago/Turabian StyleRen, Xuemei, Jinjie Chen, Shuwan Chen, Hui Zhang, and Li Li. 2023. "Genome-Wide Identification and Characterization of CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Foxtail Millet (Setaria italica L.)" Genes 14, no. 11: 2046. https://doi.org/10.3390/genes14112046
APA StyleRen, X., Chen, J., Chen, S., Zhang, H., & Li, L. (2023). Genome-Wide Identification and Characterization of CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Foxtail Millet (Setaria italica L.). Genes, 14(11), 2046. https://doi.org/10.3390/genes14112046