

Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica


Abstract
:1. Introduction
2. Two General Methods for Phagosome Isolation from E. histolytica Trophozoites
3. Phagosomal Proteins Detected in All Proteomic Studies Represent a Core Set of Constitutive Proteins Necessary for Phagosome Biogenesis
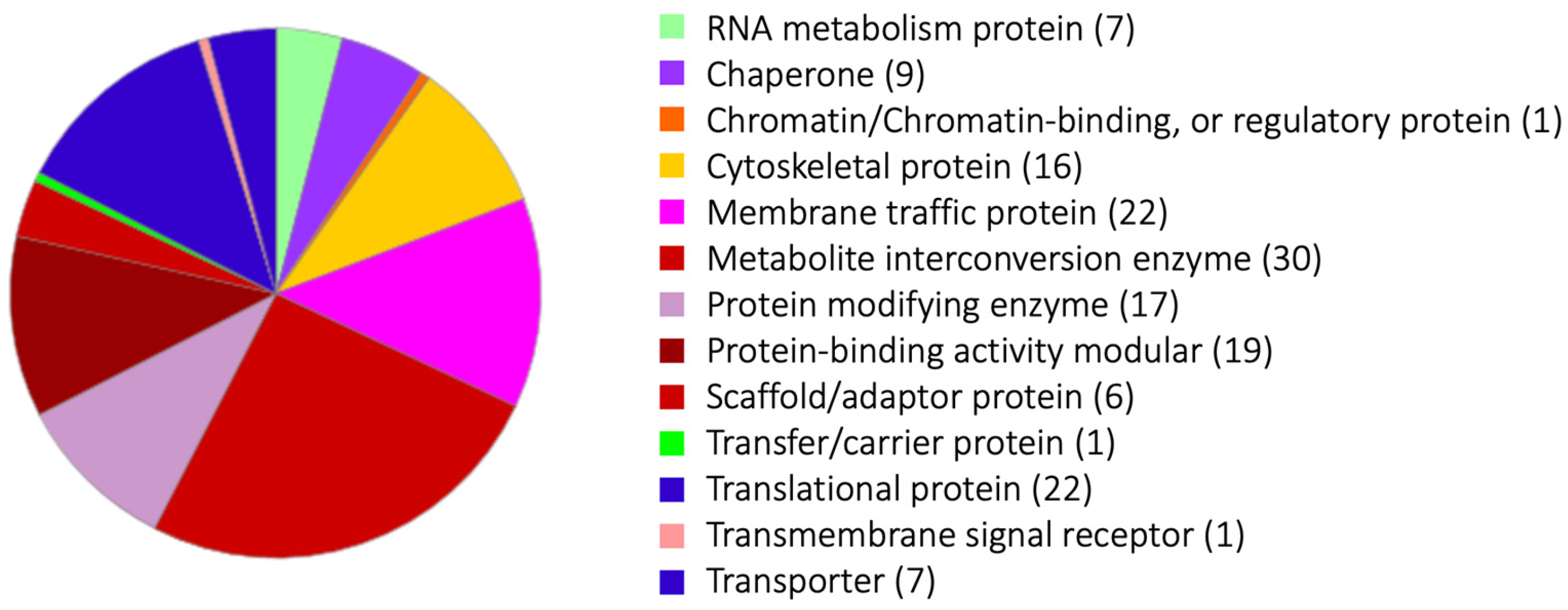
4. Phagosome Proteins Detected Exclusively by the Magnetic Bead Isolation Method, Combined with Bead Serum Coating and Chemical Cross-Linking, Represent Those Involved in Serum-Dependent Phagocytosis or Transiently Associated with Phagosomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okada, M.; Nozaki, T. New insights into molecular mechanisms of phagocytosis in Entamoeba histolytica by proteomic analysis. Arch. Med. Res. 2006, 37, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Huston, C.D.; Oue, M.; Mann, B.J.; Petri, W.A.; Kita, K.; Nozaki, T. Kinetics and strain variation of phagosome proteins of Entamoeba histolytica by proteomic analysis. Mol. Biochem. Parasitol. 2006, 145, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Marion, S.; Laurent, C.; Guillén, N. Signalization and cytoskeleton activity through myosin IB during the early steps of phagocytosis in Entamoeba histolytica: A proteomic approach. Cell. Microbiol. 2005, 7, 1504–1518. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Nakada-Tsukui, K.; Nozaki, T. Two isotypes of phosphatidylinositol 3-phosphate-binding sorting nexins play distinct roles in trogocytosis in Entamoeba histolytica. Cell Microbiol. 2020, 22, e13144. [Google Scholar] [CrossRef] [Green Version]
- Nakada-tsukui, K.; Watanabe, N.; Shibata, K.; Wahyuni, R.; Miyamoto, E.; Nozaki, T. Proteomic analysis of Atg8-dependent recruitment of phagosomal proteins in the enteric protozoan parasite Entamoeba histolytica. Front. Cell Infect. Microbiol. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Furukawa, A.; Nakada-Tsukui, K.; Nozaki, T. Cysteine protease-binding protein family 6 mediates the trafficking of amylases to phagosomes in the enteric protozoan entamoeba histolytica. Infect. Immun. 2013, 81, 1820–1829. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Huston, C.D.; Mann, B.J.; Petri, W.A.; Kita, K.; Nozaki, T. Proteomic analysis of phagocytosis in the enteric protozoan parasite Entamoeba histolytica. Eukaryot. Cell 2005, 4, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Ali, V.; Nozaki, T. A Retromerlike Complex Is a Novel Rab7 Effector That Is Involved in the Transport of the Virulence Factor Cysteine Protease in the Enteric Protozoan Parasite Entamoeba histolytica Kumiko. Mol. Biol. Cell 2005, 16, 5294–5303. [Google Scholar] [CrossRef] [Green Version]
- Saito-Nakano, Y.; Wahyuni, R.; Nakada-Tsukui, K.; Tomii, K.; Nozaki, T. Rab7D small GTPase is involved in phago-, trogocytosis and cytoskeletal reorganization in the enteric protozoan Entamoeba histolytica. Cell Microbiol. 2021, 23, e13267. [Google Scholar] [CrossRef]
- Mitra, B.N.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Sato, D.; Nozaki, T. Rab11B small GTPase regulates secretion of cysteine proteases in the enteric protozoan parasite Entamoeba histolytica. Cell Microbiol. 2007, 9, 2112–2125. [Google Scholar] [CrossRef]
- Verma, K.; Saito-Nakano, Y.; Nozaki, T.; Datta, S. Insights into endosomal maturation of human holo-transferrin in the enteric parasite Entamoeba histolytica: Essential roles of Rab7A and Rab5 in biogenesis of giant early endocytic vacuoles. Cell Microbiol. 2015, 17, 1779–1796. [Google Scholar] [CrossRef] [Green Version]
- Saito-nNkano, Y.; Mitra, B.N.; Nakada-tsukui, K.; Sato, D.; Nozaki, T. Two Rab7 isotypes, Eh Rab7A and Eh Rab7B, play distinct roles in biogenesis of lysosomes and phagosomes in the enteric protozoan parasite Entamoeba histolytica. Cell Microbiol. 2007, 9, 1796–1808. [Google Scholar] [CrossRef]
- Saito-Nakano, Y.; Yasuda, T.; Nakada-Tsukui, K.; Leippe, M.; Nozaki, T. Rab5-associated vacuoles play a unique role in phagocytosis of the enteric protozoan parasite Entamoeba histolytica. J. Biol. Chem. 2004, 279, 49497–49507. [Google Scholar] [CrossRef] [Green Version]
- Herman, E.; Siegesmund, M.A.; Bottery, M.J.; Van Aerle, R.; Shather, M.M.; Caler, E.; Dacks, J.B.; Van Der Giezen, M. Membrane Trafficking Modulation during Entamoeba Encystation. Sci. Rep. 2017, 7, 12854. [Google Scholar] [CrossRef] [Green Version]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Husain, A.; Nozaki, T. Conservation and function of Rab small GTPases in Entamoeba: Annotation of E. invadens Rab and its use for the understanding of Entamoeba biology. Exp. Parasitol. 2010, 126, 337–347. [Google Scholar] [CrossRef]
- Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Endoplasmic reticulum-resident Rab8A GTPase is involved in phagocytosis in the protozoan parasite Entamoeba histolytica. Cell Microbiol. 2016, 18, 1358–1373. [Google Scholar] [CrossRef] [Green Version]
- Hanadate, Y.; Saito-Nakano, Y.; Nakada-Tsukui, K.; Nozaki, T. Identification and characterization of the entamoeba histolytica rab8a binding protein: A Cdc50 homolog. Int. J. Mol. Sci. 2018, 19, 3831. [Google Scholar] [CrossRef] [Green Version]
- Bansal, D.; Ave, P.; Kerneis, S.; Frileux, P.; Boché, O.; Baglin, A.C.; Dubost, G.; Leguern, A.S.; Prevost, M.C.; Bracha, R.; et al. An ex-vivo human intestinal model to study Entamoeba histolytica pathogenesis. PLoS Negl. Trop. Dis. 2009, 3, 551. [Google Scholar] [CrossRef]
- Que, X.; Reed, S.L. Cysteine proteinases and the pathogenesis of amebiasis. Clin. Microbiol. Rev. 2000, 13, 196–206. [Google Scholar] [CrossRef]
- Temesvari, L.A.; Harris, E.N.; Stanley, S.L.; Cardelli, J.A. Early and late endosomal compartments of Entamoeba histolytica are enriched in cysteine proteases, acid phosphatase and several Ras-related Rab GTPases. Mol. Biochem. Parasitol. 1999, 103, 225–241. [Google Scholar] [CrossRef]
- Freitas, M.A.R.; Fernandes, H.C.; Calixto, V.C.; Martins, A.S.; Silva, E.F.; Pesquero, J.L.; Gomes, M.A. Entamoeba histolytica: Cysteine proteinase activity and virulence. Focus on cysteine proteinase 5 expression levels. Exp. Parasitol. 2009, 122, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine proteases in protozoan parasites. PLoS Negl. Trop. Dis. 2018, 12, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthiesen, J.; Bär, A.K.; Bartels, A.K.; Marien, D.; Ofori, S.; Biller, L.; Tannich, E.; Lotter, H.; Bruchhaus, I. Overexpression of specific cysteine peptidases confers pathogenicity to a nonpathogenic Entamoeba histolytica clone. MBio 2013, 4, 00072-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, B.N.; Yasuda, T.; Kobayashi, S.; Saito-Nakano, Y.; Nozaki, T. Differences in morphology of phagosomes and kinetics of acidification and degradation in phagosomes between the pathogenic Entamoeba histolytica and the non-pathogenic Entamoeba dispar. Cell Motil. Cytoskelet. 2005, 62, 84–99. [Google Scholar] [CrossRef]
- Mitra, B.N.; Kobayashi, S.; Saito-Nakano, Y.; Nozaki, T. Entamoeba histolytica: Differences in phagosome acidification and degradation between attenuated and virulent strains. Exp. Parasitol. 2006, 114, 57–61. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Fleming, D.; Chahin, L.; Rumbaugh, K. Glycoside hydrolases degrade polymicrobial bacterial biofilms in wounds. Antimicrob. Agents Chemother. 2017, 61, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Koo, I.C.; Ohol, Y.M.; Wu, P.; Morisaki, J.H.; Cox, J.S.; Brown, E.J. Role for lysosomal enzyme β-hexosaminidase in the control of mycobacteria infection. Proc. Natl. Acad. Sci. USA 2008, 105, 710–715. [Google Scholar] [CrossRef] [Green Version]
- Marumo, K.; Nakada-Tsukui, K.; Tomii, K.; Nozaki, T. Ligand heterogeneity of the cysteine protease binding protein family in the parasitic protist Entamoeba histolytica. Int. J. Parasitol. 2014, 44, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, A.; Nakada-Tsukui, K.; Nozaki, T. Novel transmembrane receptor involved in phagosome transport of lysozymes and β-hexosaminidase in the enteric protozoan Entamoeba histolytica. PLoS Pathog. 2012, 8, e1002539. [Google Scholar] [CrossRef]
- Isnard, A.; Shio, M.T.; Olivier, M. Impact of Leishmania metalloprotease GP63 on macrophage signaling. Front. Cell Infect. Microbiol. 2012, 2, 72. [Google Scholar] [CrossRef] [Green Version]
- da Silva Lira Filho, A.; Fajardo, E.F.; Chang, K.P.; Clément, P.; Olivier, M. Leishmania Exosomes/Extracellular Vesicles Containing GP63 Are Essential for Enhance Cutaneous Leishmaniasis Development Upon Co-Inoculation of Leishmania amazonensis and Its Exosomes. Front. Cell Infect. Microbiol. 2022, 11, 1–14. [Google Scholar] [CrossRef]
- Castro Neto, A.L.; Brito, A.N.A.L.M.; Rezende, A.M.; Magalhães, F.B.; De Melo Neto, O.P. In silico characterization of multiple genes encoding the GP63 virulence protein from Leishmania braziliensis: Identification of sources of variation and putative roles in immune evasion. BMC Genomics 2019, 20, 118. [Google Scholar] [CrossRef] [Green Version]
- Lieke, T.; Nylén, S.; Eidsmo, L.; McMaster, W.R.; Mohammadi, A.M.; Khamesipour, A.; Berg, L.; Akuffo, H. Leishmania surface protein gp63 binds directly to human natural killer cells and inhibits proliferation. Clin. Exp. Immunol. 2008, 153, 221–230. [Google Scholar] [CrossRef]
- Hassani, K.; Shio, M.T.; Martel, C.; Faubert, D.; Olivier, M. Absence of metalloprotease GP63 alters the protein content of leishmania exosomes. PLoS One 2014, 9, 95007. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Morgado, P.; Zhang, H.; Ehrenkaufer, G.; Manna, D.; Singh, U. Characterization of Extracellular Vesicles from Entamoeba histolytica Identifies Roles in Intracellular Communication That Regulates Parasite Growth and Development. Infect. Immun. 2020, 88, e00349-20. [Google Scholar] [CrossRef]
- Saharan, O.; Mehendale, N.; Kamat, S.S. Phagocytosis: A (Sphingo) Lipid Story. Curr. Res. Chem. Biol. 2022, 2, 100030. [Google Scholar] [CrossRef]
- Williams, E.T.; Chen, X.; Moore, D.J. VPS35, the retromer complex and Parkinson’s disease. J. Parkinsons. Dis. 2017, 7, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Welter, B.H.; Temesvari, L.A. A unique Rab GTPase, EhRabA, of Entamoeba histolytica, localizes to the leading edge of motile cells. Mol. Biochem. Parasitol. 2004, 135, 185–195. [Google Scholar] [CrossRef]
- Welter, B.H.; Temesvari, L.A. Overexpression of a mutant form of EhRabA, a unique rab GTPase of entamoeba histolytica, alters endoplasmic reticulum morphology and localization of the Gal/GalNAc adherence lectin. Eukaryot. Cell 2009, 8, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.C.; Griffiths, G.; Wessling-Resnick, M.; Fransen, J.A.M.; Bennett, H.; Jones, A.T. A role for the small GTPase Rab21 in the early endocytic pathway. J. Cell Sci. 2004, 117, 6297–6311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin-Konigsberg, R.; Grinstein, S. Phagosome-endoplasmic reticulum contacts: Kissing and not running. Traffic 2020, 21, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.K.; Yuk, J.M.; Shin, D.M.; Sasakawa, C. Roles of autophagy in elimination of intracellular bacterial pathogens. Front. Immunol. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zindrou, S.; Orozco, E.; Linder, E.; Téllez, A.; Björkman, A. Specific detection of Entamoeba histolytica DNA by hemolysin gene targeted PCR. Acta Trop. 2001, 78, 117–125. [Google Scholar] [CrossRef]
- Rath, P.P.; Gourinath, S. The actin cytoskeleton orchestra in Entamoeba histolytica. Proteins Struct. Funct. Bioinforma. 2020, 88, 1361–1375. [Google Scholar] [CrossRef]
- Bharadwaj, R.; Sharma, S.; Arya, R.; Bhattacharya, S.; Bhattacharya, A. EhRho1 regulates phagocytosis by modulating actin dynamics through EhFormin1 and EhProfilin1 in Entamoeba histolytica. Cell Microbiol. 2018, 20, e12851. [Google Scholar] [CrossRef]
- Dos Remedios, C.G.; Chhabra, D.; Kekic, M.; Dedova, I.V.; Tsubakihara, M.; Berry, D.A.; Nosworthy, N.J. Actin binding proteins: Regulation of cytoskeletal microfilaments. Physiol. Rev. 2003, 83, 433–473. [Google Scholar] [CrossRef]


{kind=link}
| Number | Journal | Year | Method | Detected Proteins | Reference |
|---|---|---|---|---|---|
| 1 | Mol. Biochem. Parasitol. | 2006 | Density-gradient | 155 | [2] |
| 2 | Arch. Med. Res. | 2006 | Density-gradient | 151 | [1] |
| 3 | Infect. Immun. | 2013 | Density-gradient | 375 | [6] |
| 4 | Cell. Microbiol. | 2019 | Magnetic beads (serum coated; cross linking) | 884 | [4] |
| 5 | Front. Cell. Infect. Microbiol. | 2022 | Magnetic beads (serum coated; cross-linking) | 738 | [5] |
| AmoebaDB Annotation | EHI Number | Common Name | Phagosome Isolation Method | Proteome Number | General Function |
|---|---|---|---|---|---|
| Rab family GTPase | EHI_108610 | Rab1A | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Small GTPase Rab7A | EHI_192810 | Rab7A | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Early phagosome maturation |
| EhRab7B protein | EHI_081330 | Rab7B | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| EhRab7D protein | EHI_082070 | Rab7D | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Phagosome maturation |
| Small GTPase Rab11B | EHI_107250 | Rab11B | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Phagosome maturation (in Eh)/CP secretion (in Eh)/endosome recycling |
| Rab family GTPase | EHI_161030 | Rab11C | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Endosome recycling |
| Rab family GTPase | EHI_153690 | RabC1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Rab family GTPase | EHI_143650 | RabC3 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Membrane traffic |
| Rho family GTPase | EHI_070730 | Rho1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Cell motility |
| Rho family GTPase | EHI_129750 | Rho7 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Cell motility |
| Ras family GTPase | EHI_058090 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Signal transduction | |
| Cell surface protease gp63, putative | EHI_200230 | Gp63 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Lysozyme, putative | EHI_199110 | Lysozyme I | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Lysozyme, putative | EHI_096570 | Lysozyme II | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase 1 | EHI_074180 | CP-A1 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Histolysain | EHI_033710 | CP-A2 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase, putative | EHI_050570 | CP-A4 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Cysteine proteinase | EHI_168240 | CP-A5 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme |
| Dipeptidyl-peptidase | EHI_136440 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| α-amylase family protein | EHI_023360 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| β-hexosaminidase | EHI_007330 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| Serine carboxypeptidase (S28) family protein | EHI_054530 | Density-gradient/ Magnetic beads | 1,2,3,4,5 | Degrading enzyme | |
| Rab family GTPase | EHI_168600 | RabA | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_079890 | RabC7 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_122730 | RabC5 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_053420 | RabI2 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_024680 | RabK1 | Magnetic beads | 4,5 | Membrane traffic |
| Rab family GTPase | EHI_117890 | RabP2 | Magnetic beads | 4,5 | Membrane traffic |
| Ras-related protein | EHI_129330 | Rab21 | Magnetic beads | 4,5 | Membrane traffic |
| Rho family GTPase | EHI_190440 | Rho10 | Magnetic beads | 4,5 | Cell motility |
| Rho family GTPase | EHI_135450 | Rho13 | Magnetic beads | 4,5 | Cell motility |
| Ras family GTPase | EHI_198330 | Not assigned | Magnetic beads | 4,5 | Signal transduction |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, N.; Nakada-Tsukui, K.; Nozaki, T. Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes 2023, 14, 379. https://doi.org/10.3390/genes14020379
Watanabe N, Nakada-Tsukui K, Nozaki T. Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes. 2023; 14(2):379. https://doi.org/10.3390/genes14020379
Chicago/Turabian StyleWatanabe, Natsuki, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki. 2023. "Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica" Genes 14, no. 2: 379. https://doi.org/10.3390/genes14020379
APA StyleWatanabe, N., Nakada-Tsukui, K., & Nozaki, T. (2023). Molecular Dissection of Phagocytosis by Proteomic Analysis in Entamoeba histolytica. Genes, 14(2), 379. https://doi.org/10.3390/genes14020379