
RNA-Sequencing Characterization of lncRNA and mRNA Functions in Septic Pig Liver Injury

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissues Samples
2.2. High-Throughput Sequencing
2.3. Analysis of Differentially Expressed (DE) LncRNAs and mRNAs
2.4. Functional Enrichment Analysis
2.5. The cis- and Trans-Target Gene Prediction of DE lncRNAs
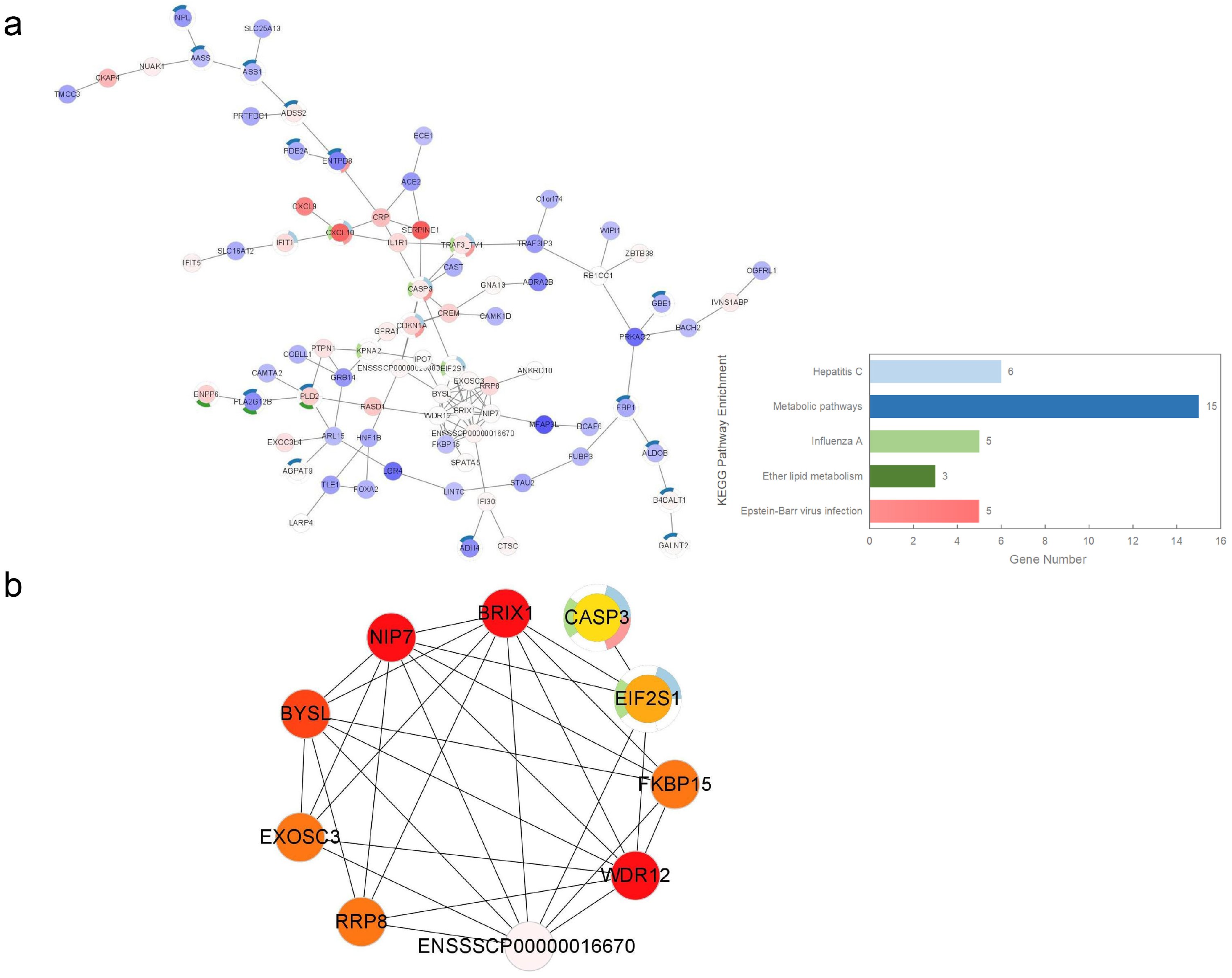
2.6. Integration of Protein-Protein Interaction Networks
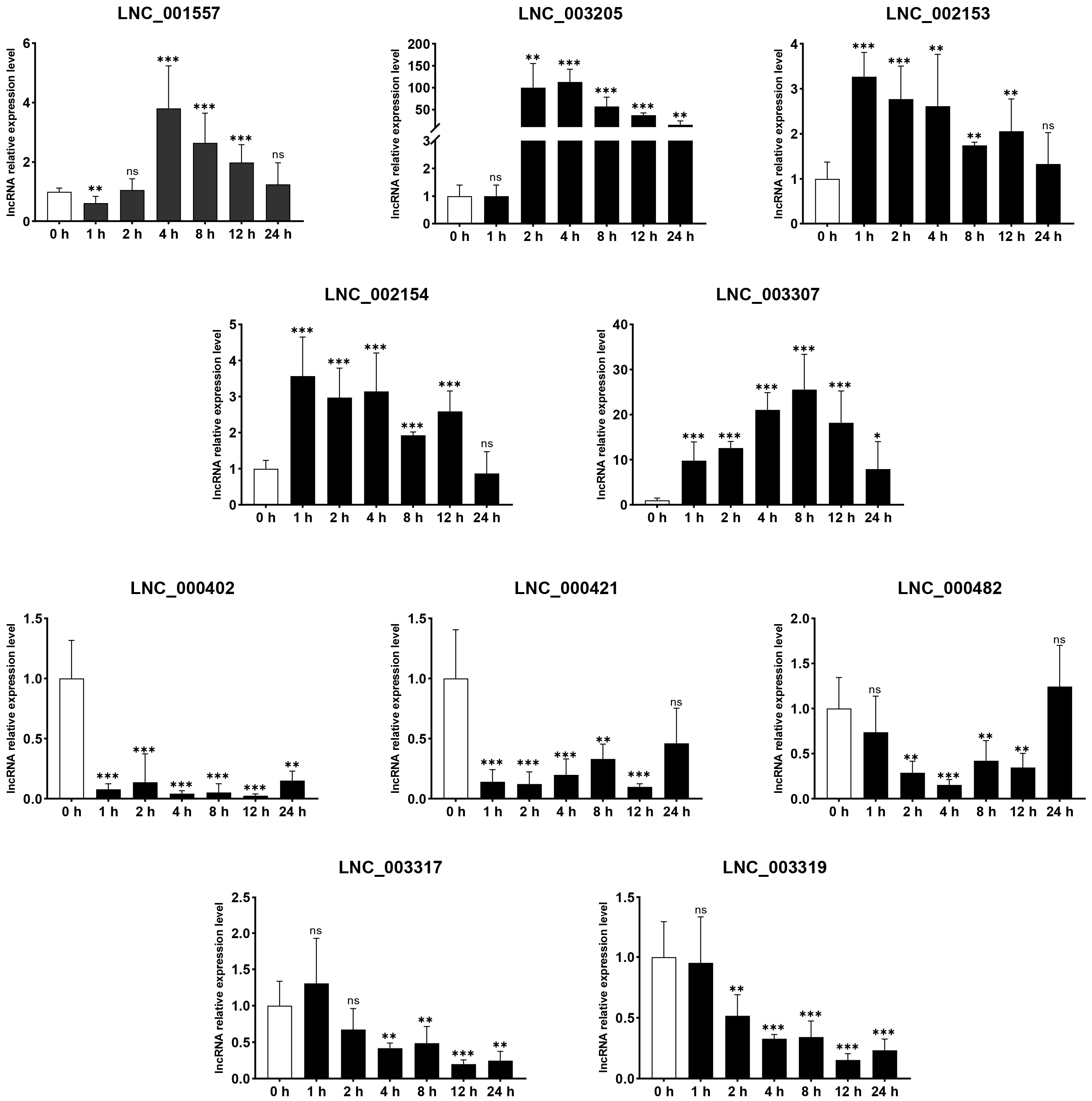
2.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.8. 5′ and 3′ Rapid Amplification of cDNA Ends (RACE)
2.9. Statistical Analysis
3. Results
3.1. Overview of RNA-Sequencing Data
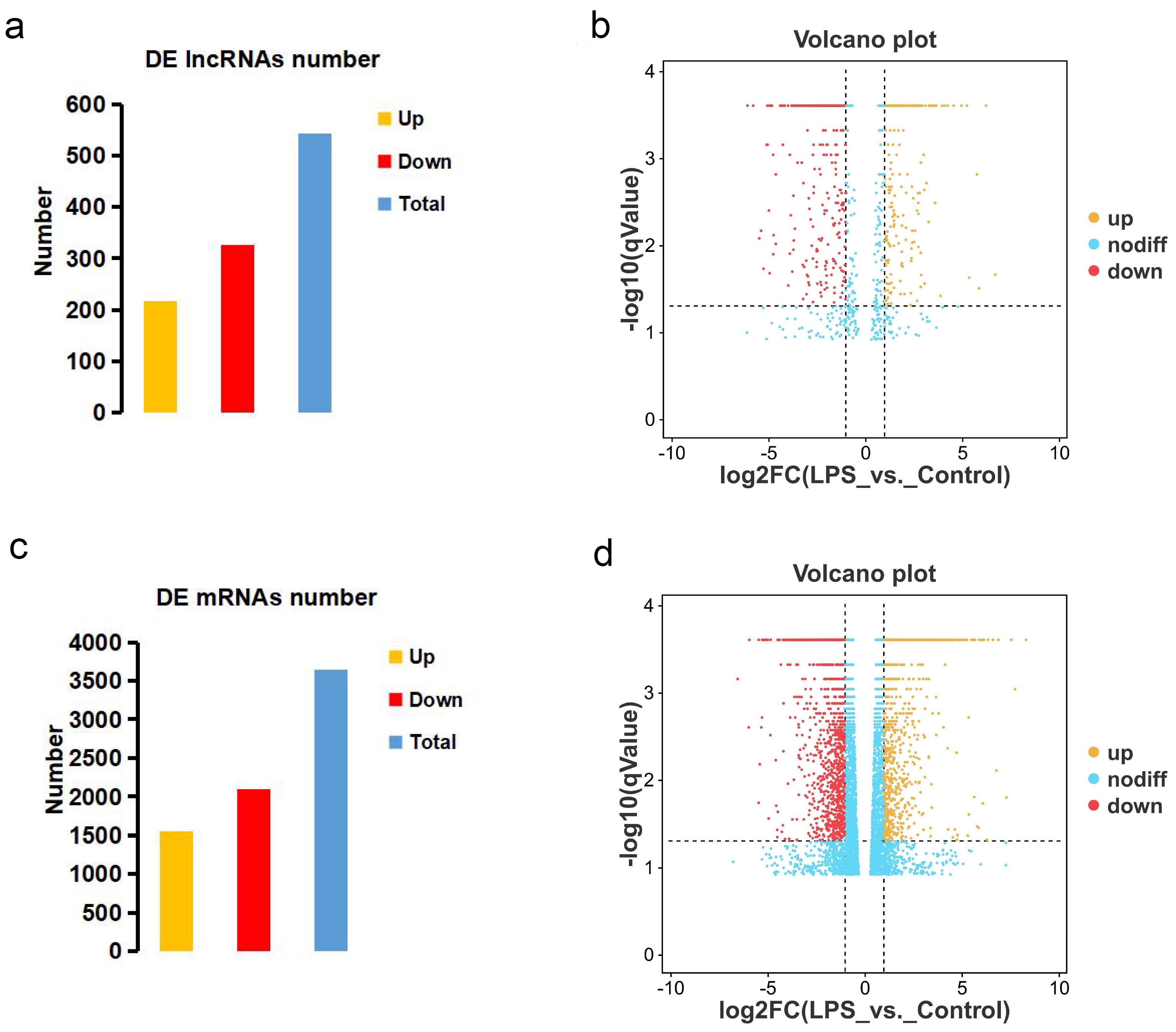
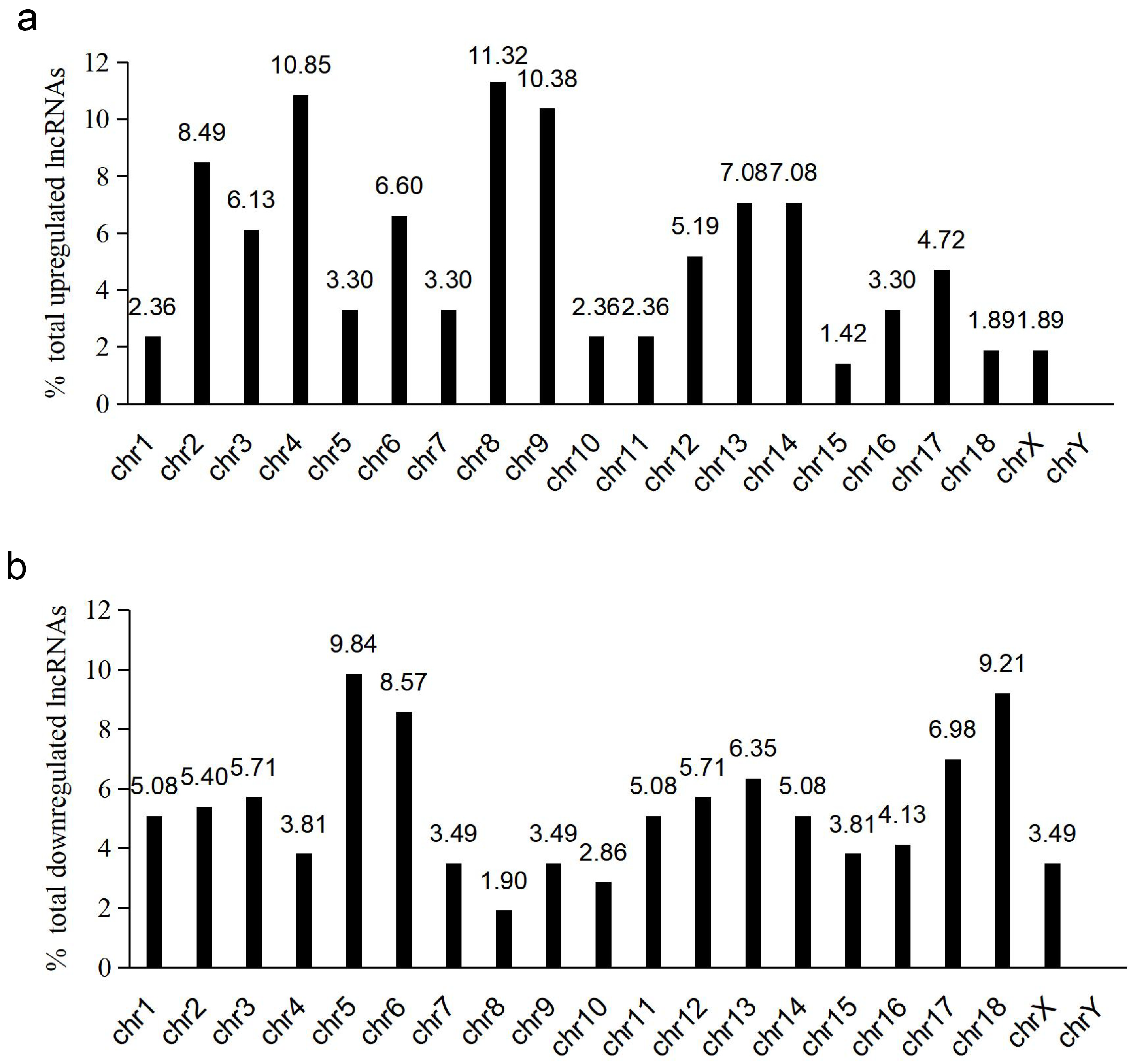
3.2. LncRNA and mRNA Expression Profiles
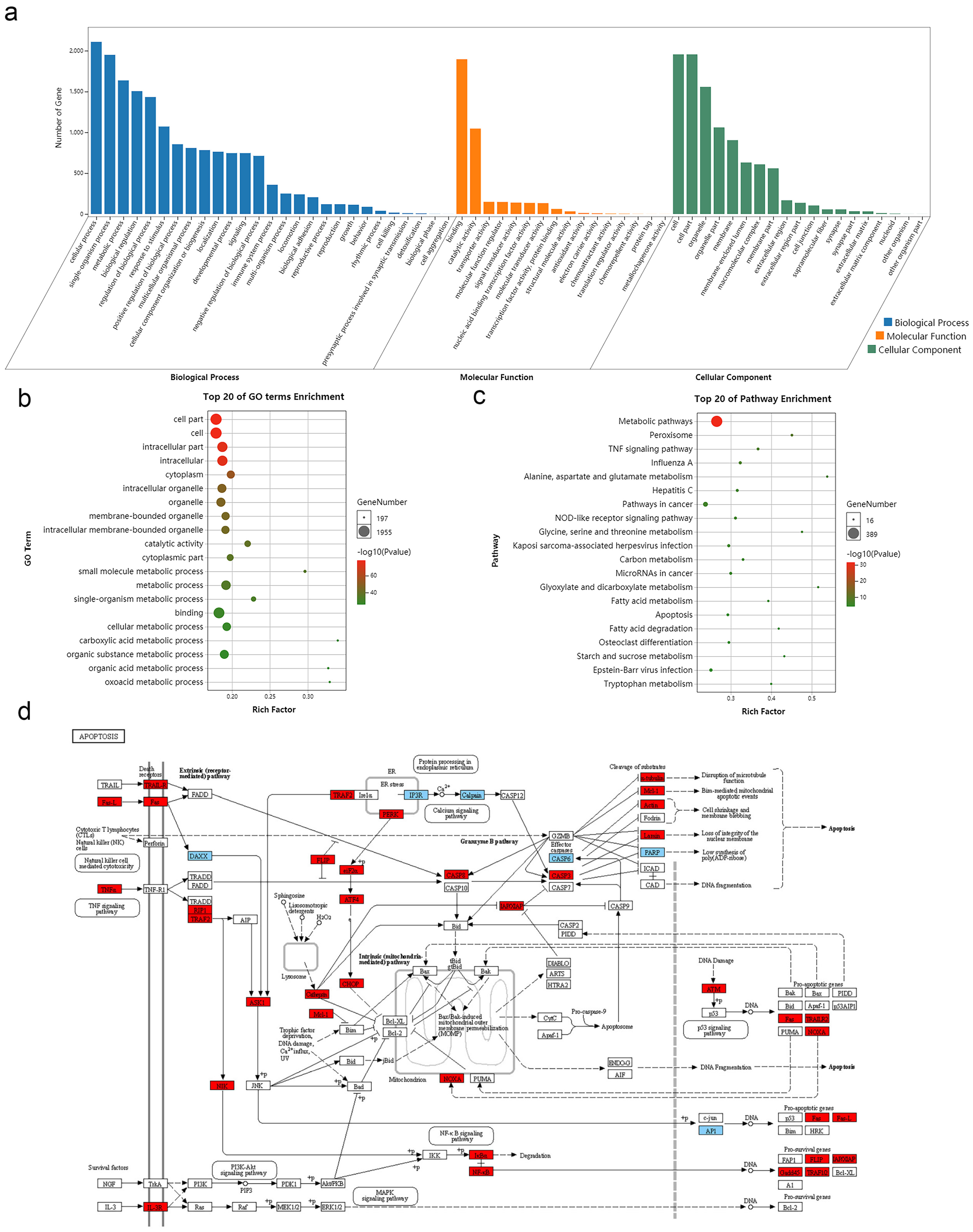
3.3. Function and Pathway Enrichment Analyses of DE mRNAs
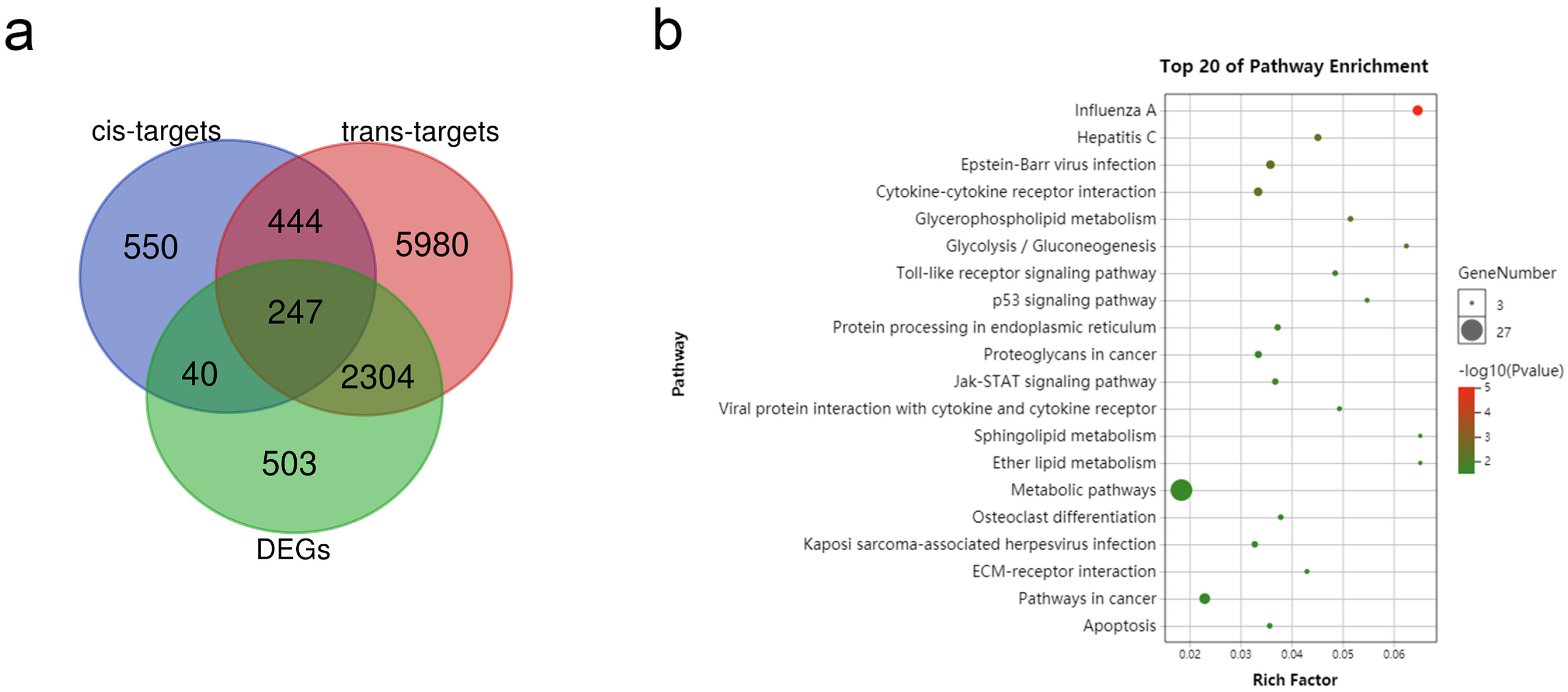
3.4. Target Gene Prediction of DE lncRNAs
3.5. PPI Network Construction and Selection of Hub Genes
3.6. qPCR Validation
3.7. Identification and Characterization of LNC_003307
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Kubes, P.; Jenne, C. Immune responses in the liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef] [PubMed]
- Matouk, A.I.; EI-Daly, M.; Habib, H.A.; Senousy, S.; Naguib Abdel Hafez, S.M.; Kasem, A.W.; Almalki, W.H.; Alzahrani, A.; Alshehri, A.; Ahmed, A.F. Protective effects of menthol against sepsis-induced hepatic injury: Role of mediators of hepatic inflammation, apoptosis, and regeneration. Front. Pharmacol. 2022, 13, 952337. [Google Scholar] [CrossRef]
- Gong, Y.; Zhu, W.; Sun, M.; Shi, L. Bioinformatics analysis of long non-coding RNA and related diseases: An overview. Front. Genet. 2021, 12, 813873. [Google Scholar] [CrossRef]
- Guo, B.; Cheng, Y.; Yao, L.; Zhang, J.; Lu, J.; Qi, H.; Hong, C. LncRNA HOTAIR regulates the lipid accumulation in non-alcoholic fatty liver disease via miR-130b-3p/ROCK1 axis. Cell. Signal. 2022, 90, 110190. [Google Scholar] [CrossRef]
- Tian, X.; Wang, Y.; Lu, Y.; Wang, W.; Du, J.; Chen, S.; Zhou, H.; Cai, W.; Xiao, Y. Conditional depletion of macrophages ameliorates cholestatic liver injury and fibrosis via lncRNA-H19. Cell Death Dis. 2021, 12, 646. [Google Scholar] [CrossRef]
- Sun, Q.; Gong, J.; Gong, X.; Wu, J.; Hu, Z.; Zhang, Q.; Zhu, X. Long non-coding RNA MALAT1 aggravated liver ischemia-reperfusion injury via targeting miR-150-5p/AZIN1. Bioengineered 2022, 13, 13422–13436. [Google Scholar] [CrossRef]
- Zhang, C.-C.; Niu, F. LncRNA NEAT1 promotes inflammatory response in sepsis-induced liver injury via the Let-7a/TLR4 axis. Int. Immunopharmacol. 2019, 75, 105731. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Zhang, Z.; Li, S.; Jiang, W.; Li, X.; Lv, J. LncRNA MALAT1 cessation antagonizes hypoxia/reoxygenation injury in hepatocytes by inhibiting apoptosis and inflammation via the HMGB1-TLR4 axis. Mol. Immunol. 2019, 112, 22–29. [Google Scholar] [CrossRef]
- Shen, C.; Li, J. LncRNA XIST silencing protects against sepsis-induced acute liver injury via inhibition of BRD4 expression. Inflammation 2021, 44, 194–205. [Google Scholar] [CrossRef]
- Li, J.; Tan, H.; Zhou, X.; Zhang, C.; Jin, H.; Tian, Y.; Zhao, X.; Li, X.; Sun, X.; Duan, M.; et al. The protection of midazolam against immune mediated liver injury induced by lipopolysaccharide and galactosamine in mice. Front. Pharmacol. 2019, 9, 1528. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Tan, Y.; Chen, S.; Xiao, X.; Zhang, M.; Wu, Q.; Dong, M. Irisin alleviates LPS-induced liver injury and inflammation through inhibition of NLRP3 inflammasome and NF-κB signaling. J. Recept. Signal Transduct. Res. 2021, 41, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Liu, J.; Du, Y.; Zhang, S.; Cao, W.; Jia, Z.; Gong, W.; Zhang, A. Estrogen-related receptor γ agonist DY131 ameliorates lipopolysaccharide-induced acute liver injury. Front. Pharmacol. 2021, 12, 626166. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, X.; Zhu, H.; Wang, Y.; Hou, Y.; Liu, Y. Dietary fish oil supplementation alters liver gene expressions to protect against LPS-induced liver injury in weanling piglets. Innate Immun. 2019, 25, 60–72. [Google Scholar] [CrossRef]
- Xu, Q.; Guo, J.; Li, X.; Wang, Y.; Wang, D.; Xiao, K.; Zhu, H.; Wang, X.; Hu, C.A.; Zhang, G.; et al. Necroptosis underlies hepatic damage in a piglet model of lipopolysaccharide-induced sepsis. Front. Immunol. 2021, 12, 633830. [Google Scholar] [CrossRef]
- Yang, Y.; Lv, Q.; Huang, X.; Fan, J.; Li, P.; Zhu, H.; Kang, P.; Liu, Y. Identification and characterization of MicroRNAs in pig liver after the LPS challenge using RNA-seq. Food Agric. Immunol. 2022, 33, 652–663. [Google Scholar] [CrossRef]
- Wang, S.; Su, W.; Zhong, C.; Yang, T.; Chen, W.; Chen, G.; Liu, Z.; Wu, K.; Zhong, W.; Li, B.; et al. An eight-circRNA assessment model for predicting biochemical recurrence in prostate cancer. Front. Cell Dev. Biol. 2020, 8, 599494. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Hu, S.; Peng, Q.; Yang, F.; Zhao, L.; Lin, Y.; Tang, Q.; Jin, L.; Ma, J.; et al. Transcriptome analysis revealed the roles of long non-coding RNA and mRNA in the bursa of Fabricius during pigeon (Columba livia) development. Front. Immunol. 2022, 13, 916086. [Google Scholar] [CrossRef]
- Cook, H.V.; Doncheva, N.T.; Szklarczyk, D.; von Mering, C.; Jensen, L.J. Viruses.STRING: A virus-host protein-protein interaction database. Viruses 2018, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.A.; Park, K.K.; Lee, S.J. LncRNAs act as a link between chronic liver disease and hepatocellular carcinoma. Int. J. Mol. Sci. 2020, 21, 2883. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Li, X.; Ren, X.; Ding, N.; Tao, L.; Dong, X.; Chen, Z. GBP5 promotes liver injury and inflammation by inducing hepatocyte apoptosis. FASEB J. 2022, 36, e22119. [Google Scholar] [CrossRef]
- Frazzette, N.; Cruz, A.C.; Wu, X.; Hammer, J.A.; Lippincott-Schwartz, J.; Siegel, R.M.; Sengupta, P. Super-resolution imaging of Fas/CD95 reorganization induced by membrane-bound Fas ligand reveals nanoscale clustering upstream of FADD recruitment. Cells. 2022, 11, 1908. [Google Scholar] [CrossRef]
- Brenner, C.; Galluzzi, L.; Kepp, O.; Kroemer, G. Decoding cell death signals in liver inflammation. J. Hepatol. 2013, 59, 583–594. [Google Scholar] [CrossRef]
- Wang, X.; Xin, H.; Xing, M.; Gu, X.; Hao, Y. Acute endoplasmic reticulum stress induces inflammation reaction, complement system activation, and lipid metabolism disorder of piglet livers: A proteomic approach. Front. Physiol. 2022, 13, 857853. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, C.; Wang, S.; Huang, Y.; Wang, H.; Zhao, J.; Yang, N. Anti-dsDNA antibodies induce inflammation via endoplasmic reticulum stress in human mesangial cells. J. Transl. Med. 2015, 13, 178. [Google Scholar] [CrossRef]
- Åberg, F.; Byrne, C.D.; Pirola, C.J.; Männistö, V.; Sookoian, S. Alcohol consumption and metabolic syndrome: Clinical and epidemiological impact on liver disease. J. Hepatol. 2023, 78, 191–206. [Google Scholar] [CrossRef]
- Antonucci, A.; Marucci, A.; Trischitta, V.; Di Paola, R. Role of GALNT2 on insulin sensitivity, lipid metabolism and fat homeostasis. Int. J. Mol. Sci. 2022, 23, 929. [Google Scholar] [CrossRef]
- Goth, C.K.; Halim, A.; Khetarpal, S.A.; Rader, D.J.; Clausen, H.; Schjoldager, K.T. A systematic study of modulation of ADAM-mediated ectodomain shedding by site-specific O-glycosylation. Proc. Natl. Acad. Sci. USA 2015, 112, 14623–14628. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Peng, X.; Zhou, X.; Li, P.; Ye, Z.; Liang, W. FXR/ASS1 axis attenuates the TAA-induced liver injury through arginine metabolism. Biochem. Biophys. Res. Commun. 2022, 611, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, X.; Yang, W.; Li, M.; Wang, G.; Luo, Q. Ferrostatin-1 ameliorates liver dysfunction via reducing iron in thioacetamide-induced acute liver injury in mice. Front. Pharmacol. 2022, 13, 869794. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, R.M.; Ahmed, H.I.; Abd El-Lateef, A.E.S.; Ali, A.A. Apoptosis perturbations and expression of regulatory inflammatory factors in cisplatin-depleted rat livers under l-arginine protection. Can J. Physiol. Pharmacol. 2019, 97, 359–369. [Google Scholar] [CrossRef]
- Huang, L.; He, X.; Peng, W.; He, X.; Xu, B.; Xu, H.; Wang, Y.; Xu, W.; Chen, W.; Wang, S.; et al. Hyperuricemia induces liver injury by upregulating HIF-1α and inhibiting arginine biosynthesis pathway in mouse liver and human L02 hepatocytes. Biochem. Biophys. Res. Commun. 2022, 617, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, Y.; Che, Z.; Zhu, H.; Meng, G.; Hou, Y.; Ding, B.; Yin, Y.; Chen, F. Dietary L-arginine supplementation alleviates liver injury caused by Escherichia coli LPS in weaned pigs. Innate immun. 2012, 18, 804–814. [Google Scholar] [CrossRef]
- Li, F.; Huangyang, P.W.; Burrows, M.; Guo, K.; Riscal, R.; Godfrey, J.; Lee, K.E.; Lin, N.; Lee, P.; Blair, I.A.; et al. FBP1 loss disrupts liver metabolism and promotes tumorigenesis through a hepatic stellate cell senescence secretome. Nat. Cell Biol. 2020, 22, 728–739. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, J.; Zhao, Z.; Zhu, Y.; Xing, J.; An, J.; Guo, X. Low expression of phosphodiesterase 2 (PDE2A) promotes the progression by regulating mmitochondrial morphology and ATP content and predicts poor prognosis in hepatocellular carcinoma. Cells. 2022, 12, 68. [Google Scholar] [CrossRef]
- Lobo, M.J.; Reverte-Salisa, L.; Chao, Y.C.; Koschinski, A.; Gesellchen, F.; Subramaniam, G.; Jiang, H.; Pace, S.; Larcom, N.; Paolocci, E.; et al. Phosphodiesterase 2A2 regulates mitochondria clearance through Parkin-dependent mitophagy. Commun. Biol. 2020, 3, 596. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Zhao, J.; Zhang, Z.; Jin, M.; Zhou, F.; Jin, C.; Zhang, J.; Xing, J.; Wang, N.; et al. PDE2 inhibits PKA-mediated phosphorylation of TFAM to promote mitochondrial Ca2+-induced colorectal cancer growth. Front. Oncol. 2021, 11, 663778. [Google Scholar] [CrossRef]
- Barbagallo, F.; Rotilio, V.; Assenza, M.R.; Aguanno, S.; Orsini, T.; Putti, S.; Isidori, A.M.; Lenzi, A.; Naro, F.; De Angelis, L.; et al. PDE2A is indispensable for mouse liver development and hematopoiesis. Int. J. Mol. Sci. 2020, 21, 2902. [Google Scholar] [CrossRef] [PubMed]
- Rentsendorj, O.; D’Alessio, F.R.; Pearse, D.B. Phosphodiesterase 2A is a major negative regulator of iNOS expression in lipopolysaccharide-treated mouse alveolar macrophages. J. Leukoc. Biol. 2014, 96, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Morello, L.G.; Coltri, P.P.; Quaresma, A.J.; Simabuco, F.M.; Silva, T.C.; Singh, G.; Nickerson, J.A.; Oliveira, C.C.; Moore, M.J.; Zanchin, N.I. The human nucleolar protein FTSJ3 associates with NIP7 and functions in pre-rRNA processing. PLoS ONE 2011, 6, e29174. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Huang, X.; Wang, P.; Lu, C. Expression of biogenesis of ribosomes BRX1 is associated with malignant progression and prognosis in colorectal cancer. Transl. Cancer Res. 2020, 9, 5595–5602. [Google Scholar] [CrossRef]
- Mi, L.; Qi, Q.; Ran, H.; Chen, L.; Li, D.; Xiao, D.; Wu, J.; Cai, Y.; Zhang, S.; Li, Y.; et al. Suppression of ribosome biogenesis by targeting WD repeat domain 12 (WDR12) inhibits glioma stem-like cell growth. Front. Oncol. 2021, 11, 751792. [Google Scholar] [CrossRef]
- Song, Q.; Chen, Y.; Wang, J.; Hao, L.; Huang, C.; Griffiths, A.; Sun, Z.; Zhou, Z.; Song, Z. ER stress-induced upregulation of NNMT contributes to alcohol-related fatty liver development. J. Hepatol. 2020, 73, 783–793. [Google Scholar] [CrossRef]
- Anzaghe, M.; Resch, T.; Schaser, E.; Kronhart, S.; Diez, C.; Niles, M.A.; Korotkova, E.; Schülke, S.; Wolfheimer, S.; Kreuz, D.; et al. Organ-specific expression of IL-1 receptor results in severe liver injury in type I interferon receptor deficient mice. Front. Immunol. 2019, 10, 1009. [Google Scholar] [CrossRef]
- Yun, Y.; Chen, J.; Wang, X.; Li, Y.; Hu, Z.; Yang, P.; Qin, L. Tofacitinib ameliorates lipopolysaccharide-induced acute kidney injury by blocking the JAK-STAT1/STAT3 signaling pathway. Biomed. Res. Int. 2021, 2021, 8877056. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wise, J.T.; Shi, X.; Chen, Z. Verteporfin inhibits lipopolysaccharide-induced inflammation by multiple functions in RAW 264.7 cells. Toxicol. Appl. Pharmacol. 2019, 387, 114852. [Google Scholar] [CrossRef]
- Ho, D.V.; Chan, J.Y. Induction of Herpud1 expression by ER stress is regulated by Nrf1. FEBS Lett. 2015, 589, 615–620. [Google Scholar] [CrossRef]
- Li, S.X.; Li, J.; Dong, L.W.; Guo, Z.Y. Cytoskeleton-associated protein 4, a promising biomarker for tumor diagnosis and therapy. Front. Mol. Biosci. 2021, 7, 552056. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A cell’s response to stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Oyang, L.; Liang, J.; Yi, P.; Han, Y.; Luo, X.; Xia, L.; Lin, J.; Tan, S.; Hu, J.; et al. Biological function of HYOU1 in tumors and other diseases. Onco Targets Ther. 2020, 14, 1727–1735. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Mu, X.; Wu, H.; Chen, L.; Liu, J.; Zhao, Y. Calreticulin (CALR)-induced activation of NF-ĸB signaling pathway boosts lung cancer cell proliferation. Bioengineered 2022, 13, 6856–6865. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Q30 (%) | Clean Reads | Clean Ratio (%) | Mapped Reads | Mapped Ratio (%) |
|---|---|---|---|---|---|---|
| N1 | 103,431,076 | 95.15 | 101,547,492 | 98.18 | 84,293,797 | 83.01 |
| N2 | 153,426,742 | 95.37 | 150,724,416 | 98.24 | 124,265,319 | 82.45 |
| N3 | 112,256,790 | 95.17 | 110,219,924 | 98.19 | 90,385,080 | 82.00 |
| L1 | 130,247,426 | 95.28 | 127,916,070 | 98.21 | 103,566,992 | 80.96 |
| L2 | 164,291,314 | 95.24 | 161,346,466 | 98.21 | 129,899,294 | 80.51 |
| L3 | 139,897,100 | 94.90 | 137,305,488 | 98.15 | 113,383,824 | 82.58 |
| DE lncRNAs | Liver_LPS FPKM | Liver_Control FPKM | log2(FC) | q-Value |
|---|---|---|---|---|
| LNC_003307 | 272.26 | 61.8278 | 2.13866 | 0.000249496 |
| LNC_002814 | 15.5746 | 37.2039 | −1.25626 | 0.000249496 |
| LNC_002495 | 13.6656 | 32.3223 | −1.24198 | 0.000480016 |
| LNC_001034 | 6.56286 | 25.9666 | −1.98426 | 0.0159756 |
| LNC_000416 | 4.60465 | 21.7599 | −2.24051 | 0.000249496 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Xue, Z.; Zhao, Q.; Zhang, K.; Zhou, A.; Shi, L.; Liu, Y. RNA-Sequencing Characterization of lncRNA and mRNA Functions in Septic Pig Liver Injury. Genes 2023, 14, 945. https://doi.org/10.3390/genes14040945
Zhang J, Xue Z, Zhao Q, Zhang K, Zhou A, Shi L, Liu Y. RNA-Sequencing Characterization of lncRNA and mRNA Functions in Septic Pig Liver Injury. Genes. 2023; 14(4):945. https://doi.org/10.3390/genes14040945
Chicago/Turabian StyleZhang, Jing, Zhihui Xue, Qingbo Zhao, Keke Zhang, Ao Zhou, Liangyu Shi, and Yulan Liu. 2023. "RNA-Sequencing Characterization of lncRNA and mRNA Functions in Septic Pig Liver Injury" Genes 14, no. 4: 945. https://doi.org/10.3390/genes14040945
APA StyleZhang, J., Xue, Z., Zhao, Q., Zhang, K., Zhou, A., Shi, L., & Liu, Y. (2023). RNA-Sequencing Characterization of lncRNA and mRNA Functions in Septic Pig Liver Injury. Genes, 14(4), 945. https://doi.org/10.3390/genes14040945