
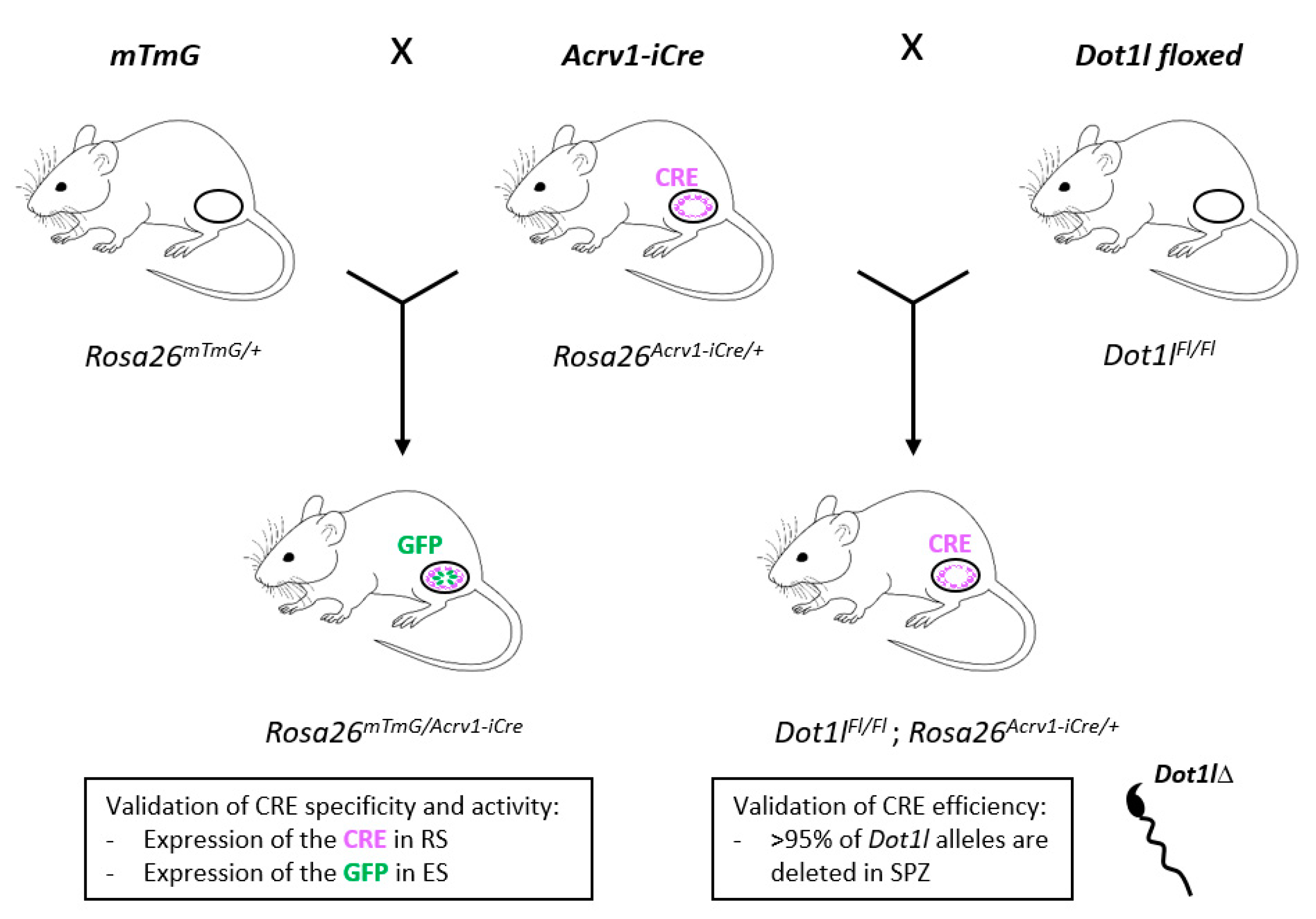
Generation and Characterization of a Transgenic Mouse That Specifically Expresses the Cre Recombinase in Spermatids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
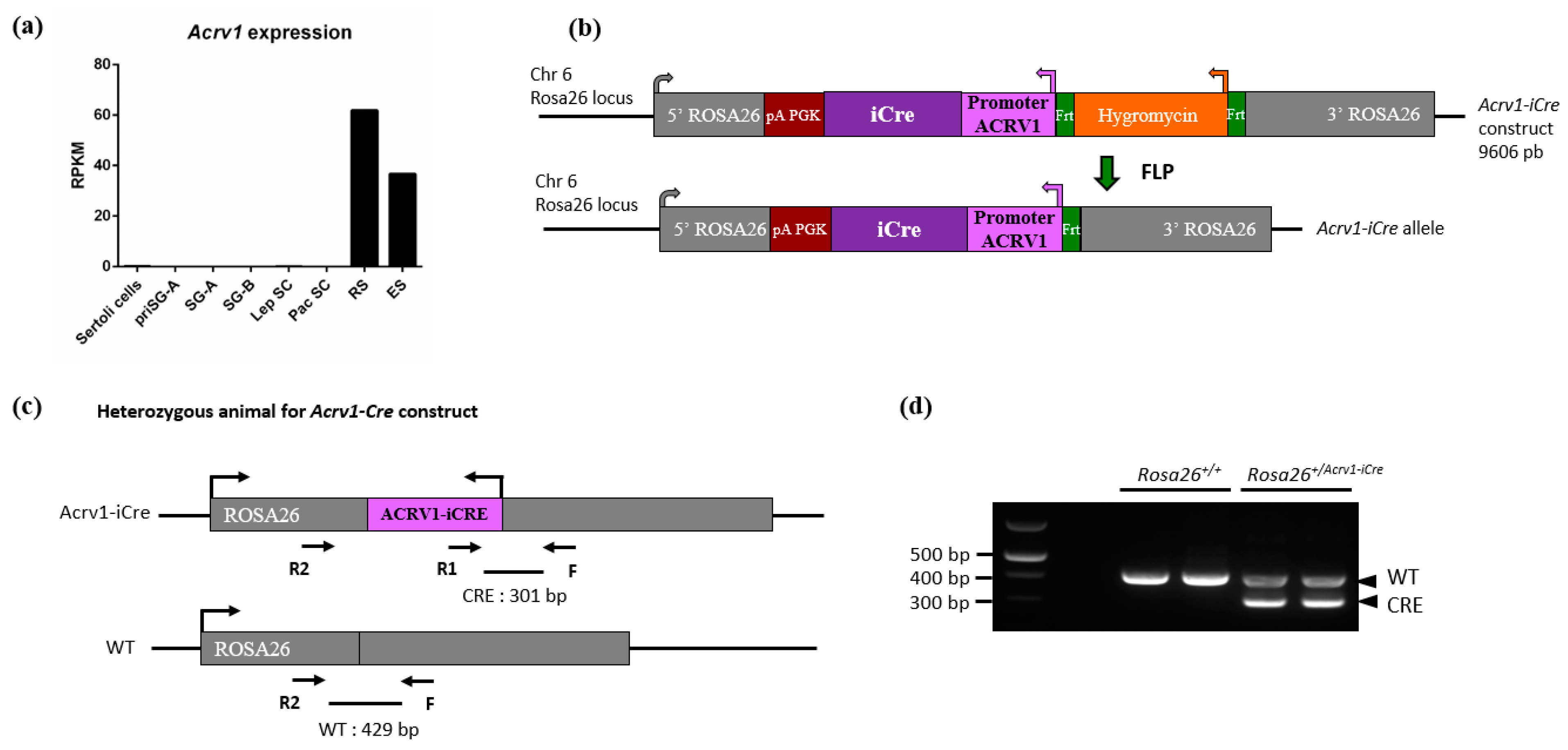
2.1. Generation of Transgenic Acrv1-iCre Mice
2.2. Other Mouse Strains and Genotyping
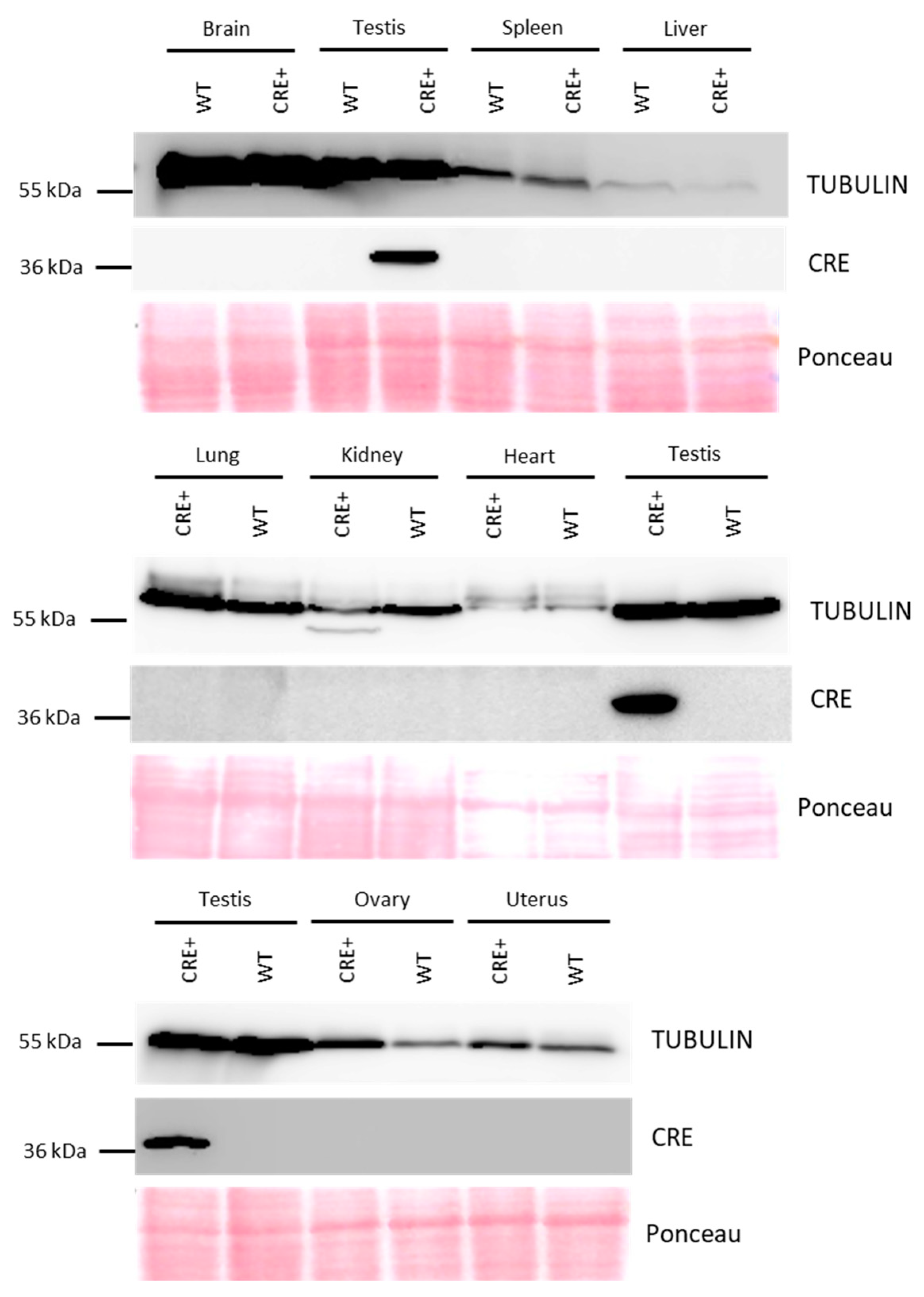
2.3. Western Blot Analysis of Cre Expression
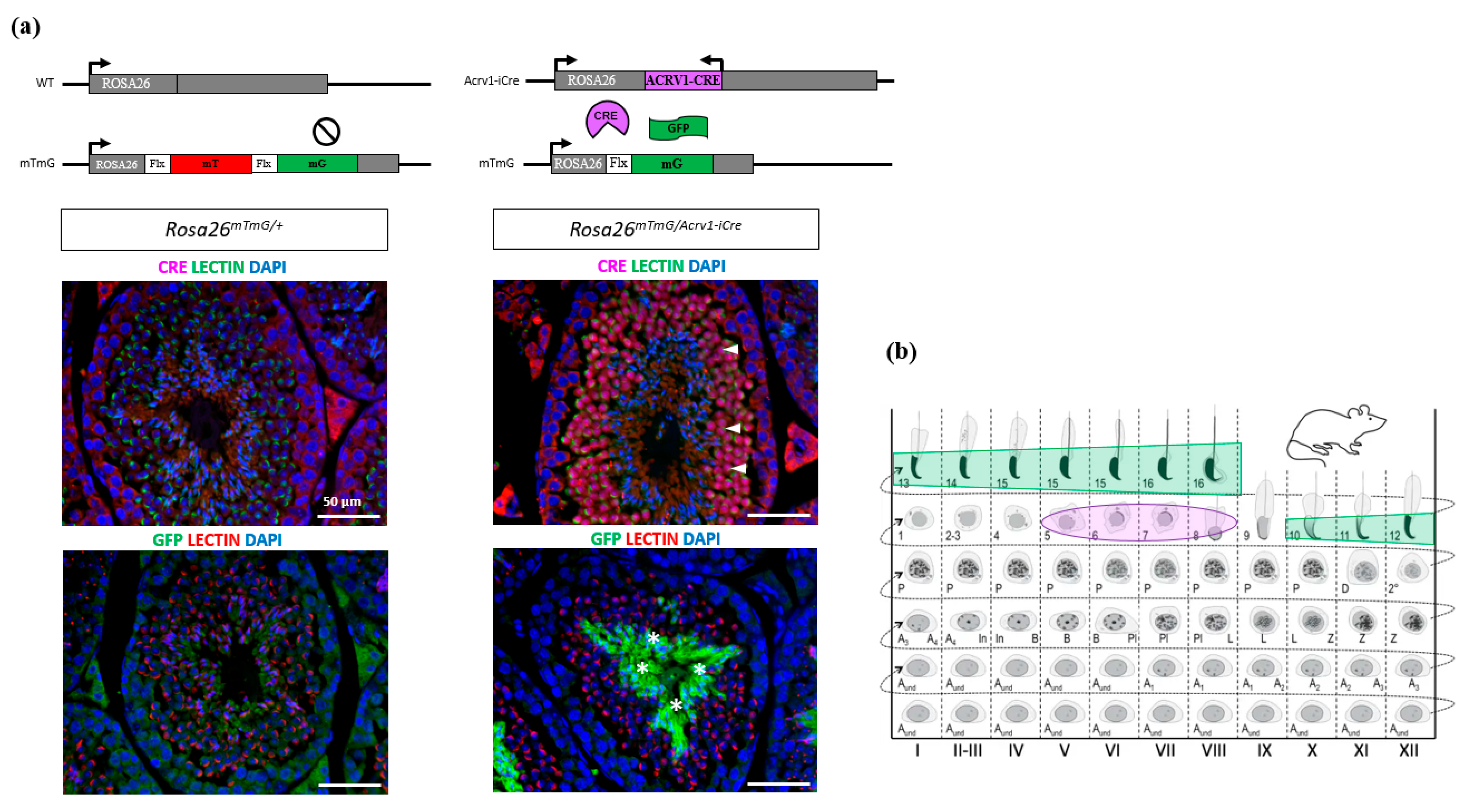
2.4. Immunofluorescence on Testis Section
2.5. Sperm Collection and Genomic DNA Extraction
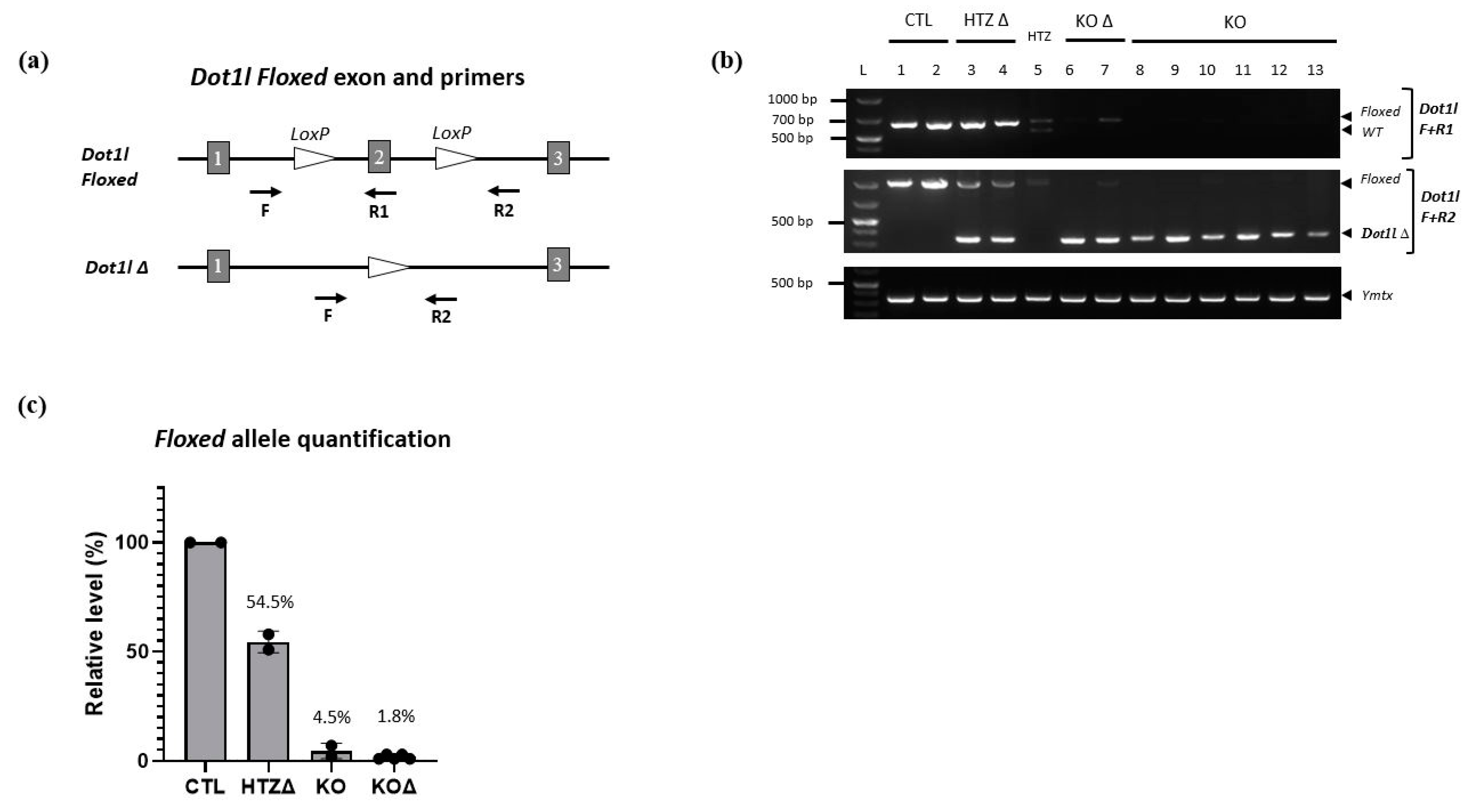
2.6. Assessment of Acrv1-iCre Recombination Efficiency
3. Results
3.1. Acrv1-iCre Expression Is Testis-Specific and Restricted to Round Spermatids
3.2. Acrv1-iCre Line Is Efficient to Knock out Genes during Spermiogenesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Houston, B.J.; Riera-Escamilla, A.; Wyrwoll, M.J.; Salas-Huetos, A.; Xavier, M.J.; Nagirnaja, L.; Friedrich, C.; Conrad, D.F.; Aston, K.I.; Krausz, C.; et al. A systematic review of the validated monogenic causes of human male infertility: 2020 update and a discussion of emerging gene-disease relationships. Hum. Reprod. Update 2022, 28, 15–29. [Google Scholar] [CrossRef]
- Ahmed, E.A.; de Rooij, D.G. Staging of mouse seminiferous tubule cross-sections. Methods Mol. Biol. 2009, 558, 263–277. [Google Scholar] [CrossRef]
- Muciaccia, B.; Boitani, C.; Berloco, B.P.; Nudo, F.; Spadetta, G.; Stefanini, M.; de Rooij, D.G.; Vicini, E. Novel stage classification of human spermatogenesis based on acrosome development. Biol. Reprod. 2013, 89, 60. [Google Scholar] [CrossRef]
- da Cruz, I.; Rodriguez-Casuriaga, R.; Santinaque, F.F.; Farias, J.; Curti, G.; Capoano, C.A.; Folle, G.A.; Benavente, R.; Sotelo-Silveira, J.R.; Geisinger, A. Transcriptome analysis of highly purified mouse spermatogenic cell populations: Gene expression signatures switch from meiotic-to postmeiotic-related processes at pachytene stage. BMC Genom. 2016, 17, 294. [Google Scholar] [CrossRef]
- Maezawa, S.; Sakashita, A.; Yukawa, M.; Chen, X.; Takahashi, K.; Alavattam, K.G.; Nakata, I.; Weirauch, M.T.; Barski, A.; Namekawa, S.H. Super-enhancer switching drives a burst in gene expression at the mitosis-to-meiosis transition. Nat. Struct. Mol. Biol. 2020, 27, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Archambeault, D.R.; Matzuk, M.M. Disrupting the male germ line to find infertility and contraception targets. Ann. Endocrinol. 2014, 75, 101–108. [Google Scholar] [CrossRef]
- Gu, H.; Marth, J.D.; Orban, P.C.; Mossmann, H.; Rajewsky, K. Deletion of a DNA polymerase beta gene segment in T cells using cell type-specific gene targeting. Science 1994, 265, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Smith, L. Good planning and serendipity: Exploiting the Cre/Lox system in the testis. Reproduction 2011, 141, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Kido, T.; Lau, Y.F. A Cre gene directed by a human TSPY promoter is specific for germ cells and neurons. Genesis 2005, 42, 263–275. [Google Scholar] [CrossRef]
- Schmidt, E.E.; Taylor, D.S.; Prigge, J.R.; Barnett, S.; Capecchi, M.R. Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse spermatids. Proc. Natl. Acad. Sci. USA 2000, 97, 13702–13707. [Google Scholar] [CrossRef]
- Shimshek, D.R.; Kim, J.; Hubner, M.R.; Spergel, D.J.; Buchholz, F.; Casanova, E.; Stewart, A.F.; Seeburg, P.H.; Sprengel, R. Codon-improved Cre recombinase (iCre) expression in the mouse. Genesis 2002, 32, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Reddi, P.P.; Shore, A.N.; Shapiro, J.A.; Anderson, A.; Stoler, M.H.; Acharya, K.K. Spermatid-specific promoter of the SP-10 gene functions as an insulator in somatic cells. Dev. Biol. 2003, 262, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Reddi, P.P.; Flickinger, C.J.; Herr, J.C. Round spermatid-specific transcription of the mouse SP-10 gene is mediated by a 294-base pair proximal promoter. Biol. Reprod. 1999, 61, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Kanki, H.; Suzuki, H.; Itohara, S. High-efficiency CAG-FLPe deleter mice in C57BL/6J background. Exp. Anim. 2006, 55, 137–141. [Google Scholar] [CrossRef]
- Blanco, M.; El Khattabi, L.; Gobé, C.; Crespo, M.; Coulée, M.; De la Iglesia, A.; Ialy-Radio, C.; Lapoujade, C.; Givelet, M.; Delessard, M.; et al. The histone methyltransferase DOT1L regulates chromatin reorganization and gene expression during the postmeiotic differentiation of male germ cells. Embo Reports bioRxiv, 2023; in press. [Google Scholar] [CrossRef]
- Muzumdar, M.D.; Tasic, B.; Miyamichi, K.; Li, L.; Luo, L. A global double-fluorescent Cre reporter mouse. Genesis 2007, 45, 593–605. [Google Scholar] [CrossRef]
- Jo, S.Y.; Granowicz, E.M.; Maillard, I.; Thomas, D.; Hess, J.L. Requirement for Dot1l in murine postnatal hematopoiesis and leukemogenesis by MLL translocation. Blood 2011, 117, 4759–4768. [Google Scholar] [CrossRef]
- Comptour, A.; Moretti, C.; Serrentino, M.E.; Auer, J.; Ialy-Radio, C.; Ward, M.A.; Toure, A.; Vaiman, D.; Cocquet, J. SSTY proteins co-localize with the post-meiotic sex chromatin and interact with regulators of its expression. FEBS J. 2014, 281, 1571–1584. [Google Scholar] [CrossRef]
- Kocer, A.; Henry-Berger, J.; Noblanc, A.; Champroux, A.; Pogorelcnik, R.; Guiton, R.; Janny, L.; Pons-Rejraji, H.; Saez, F.; Johnson, G.D.; et al. Oxidative DNA damage in mouse sperm chromosomes: Size matters. Free. Radic. Biol. Med. 2015, 89, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Wen, L.; Liao, S.; Lin, X.; Ma, T.; Liu, J.; Song, C.X.; Wang, M.; He, C.; Han, C.; et al. Dynamics of 5-hydroxymethylcytosine during mouse spermatogenesis. Nat. Commun. 2013, 4, 1995. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.; Serrentino, M.E.; Ialy-Radio, C.; Delessard, M.; Soboleva, T.A.; Tores, F.; Leduc, M.; Nitschke, P.; Drevet, J.R.; Tremethick, D.J.; et al. SLY regulates genes involved in chromatin remodeling and interacts with TBL1XR1 during sperm differentiation. Cell Death Differ. 2017, 24, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Makela, J.A.; Cisneros-Montalvo, S.; Lehtiniemi, T.; Olotu, O.; La, H.M.; Toppari, J.; Hobbs, R.M.; Parvinen, M.; Kotaja, N. Transillumination-Assisted Dissection of Specific Stages of the Mouse Seminiferous Epithelial Cycle for Downstream Immunostaining Analyses. J. Vis. Exp. 2020, e61800. [Google Scholar] [CrossRef]
- Sadate-Ngatchou, P.I.; Payne, C.J.; Dearth, A.T.; Braun, R.E. Cre recombinase activity specific to postnatal, premeiotic male germ cells in transgenic mice. Genesis 2008, 46, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, T.; Shirley, L.; John, G.B.; Castrillon, D.H. Generation of a germ cell-specific mouse transgenic Cre line, Vasa-Cre. Genesis 2007, 45, 413–417. [Google Scholar] [CrossRef]
- Schonhoff, S.E.; Giel-Moloney, M.; Leiter, A.B. Neurogenin 3-expressing progenitor cells in the gastrointestinal tract differentiate into both endocrine and non-endocrine cell types. Dev. Biol. 2004, 270, 443–454. [Google Scholar] [CrossRef]
- Bao, J.; Ma, H.Y.; Schuster, A.; Lin, Y.M.; Yan, W. Incomplete cre-mediated excision leads to phenotypic differences between Stra8-iCre; Mov10l1(lox/lox) and Stra8-iCre; Mov10l1(lox/Delta) mice. Genesis 2013, 51, 481–490. [Google Scholar] [CrossRef]
- Hirota, T.; Blakeley, P.; Sangrithi, M.N.; Mahadevaiah, S.K.; Encheva, V.; Snijders, A.P.; ElInati, E.; Ojarikre, O.A.; de Rooij, D.G.; Niakan, K.K.; et al. SETDB1 Links the Meiotic DNA Damage Response to Sex Chromosome Silencing in Mice. Dev. Cell 2018, 47, 645–659.e646. [Google Scholar] [CrossRef]
- Hobbs, R.M.; La, H.M.; Makela, J.A.; Kobayashi, T.; Noda, T.; Pandolfi, P.P. Distinct germline progenitor subsets defined through Tsc2-mTORC1 signaling. EMBO Rep. 2015, 16, 467–480. [Google Scholar] [CrossRef]
- Song, J.; Xu, Y.; Hu, X.; Choi, B.; Tong, Q. Brain expression of Cre recombinase driven by pancreas-specific promoters. Genesis 2010, 48, 628–634. [Google Scholar] [CrossRef]
- O’Gorman, S.; Dagenais, N.A.; Qian, M.; Marchuk, Y. Protamine-Cre recombinase transgenes efficiently recombine target sequences in the male germ line of mice, but not in embryonic stem cells. Proc. Natl. Acad. Sci. USA 1997, 94, 14602–14607. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gobé, C.; Ialy-Radio, C.; Pierre, R.; Cocquet, J. Generation and Characterization of a Transgenic Mouse That Specifically Expresses the Cre Recombinase in Spermatids. Genes 2023, 14, 983. https://doi.org/10.3390/genes14050983
Gobé C, Ialy-Radio C, Pierre R, Cocquet J. Generation and Characterization of a Transgenic Mouse That Specifically Expresses the Cre Recombinase in Spermatids. Genes. 2023; 14(5):983. https://doi.org/10.3390/genes14050983
Chicago/Turabian StyleGobé, Clara, Côme Ialy-Radio, Rémi Pierre, and Julie Cocquet. 2023. "Generation and Characterization of a Transgenic Mouse That Specifically Expresses the Cre Recombinase in Spermatids" Genes 14, no. 5: 983. https://doi.org/10.3390/genes14050983
APA StyleGobé, C., Ialy-Radio, C., Pierre, R., & Cocquet, J. (2023). Generation and Characterization of a Transgenic Mouse That Specifically Expresses the Cre Recombinase in Spermatids. Genes, 14(5), 983. https://doi.org/10.3390/genes14050983