
Effect of Bovine MEF2A Gene Expression on Proliferation and Apoptosis of Myoblast Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. MEF2A Overexpression and Construction of Interference Vector
2.3. Cell Identification and Transfection
2.4. Cell Proliferation Activity Assay
2.5. ELISA for GH and INS
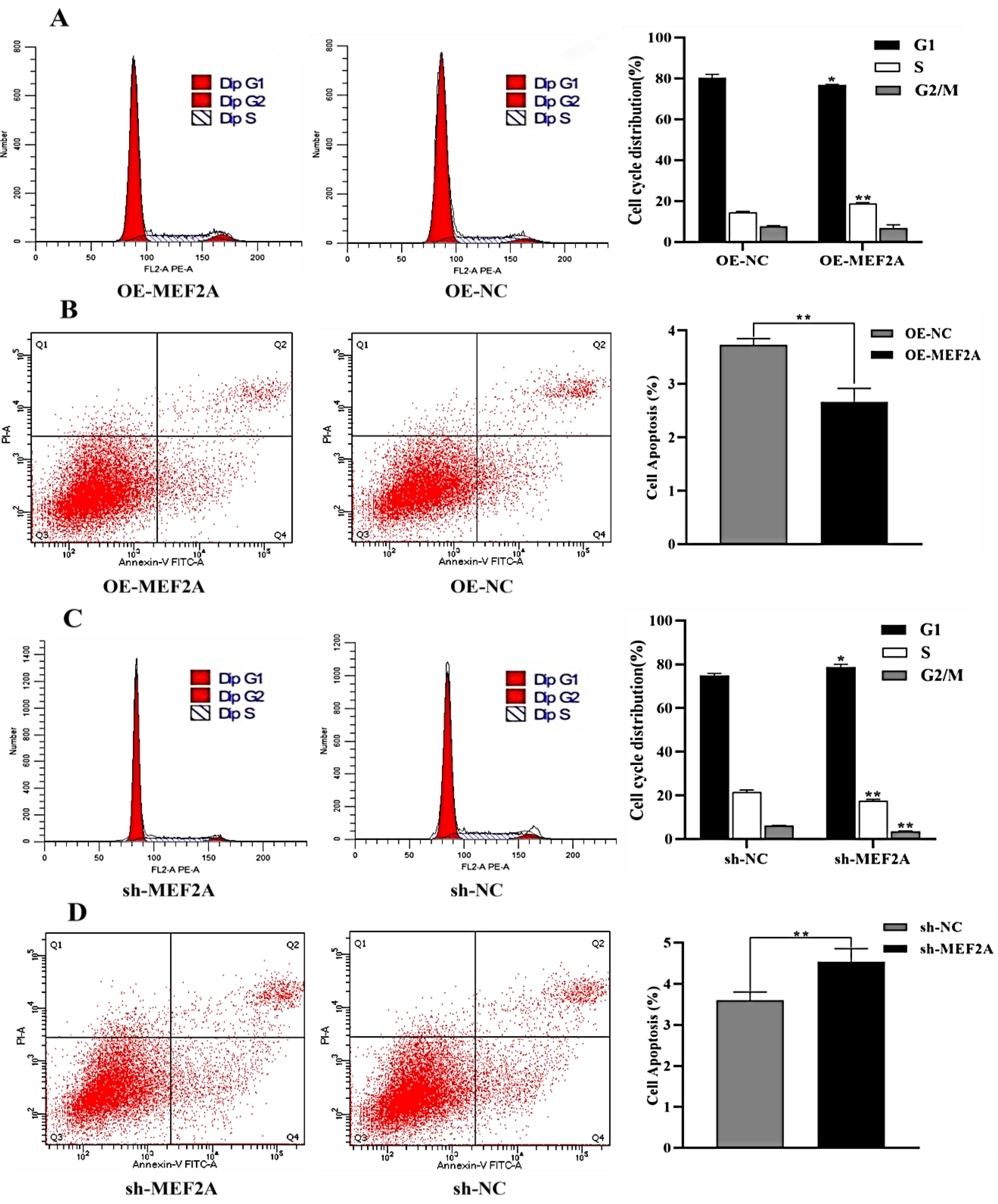
2.6. Flow Cytometry for Cycle Assay
2.7. Flow Cytometry Assay for Apoptosis Detection
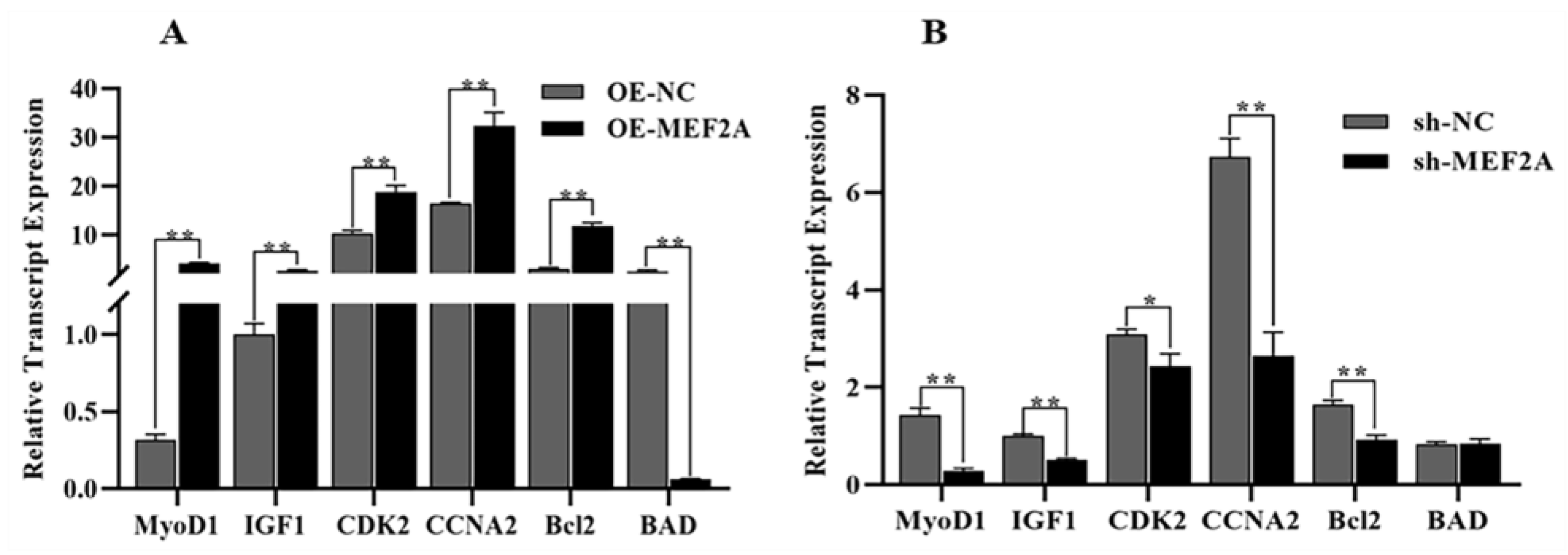
2.8. Real-Time Quantitative PCR
2.9. Statistical Analysis
3. Results
3.1. Cell Purity
3.2. Carrier Efficiency
3.3. Cell Viability Assay
3.4. GH and INS Analysis
3.5. Cell Cycle and Apoptosis
3.6. RT-qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Picard, B.; Berri, C.; Lefaucheur, L.; Molette, C.; Sayd, T.; Terlouw, C. Skeletal muscle proteomics in livestockproduction. Brief. Funct. Genom. 2010, 9, 259–278. [Google Scholar] [CrossRef]
- Motoyama, M.; Sasaki, K.; Watanabe, A. Wagyu and the factors contributing to its beef quality: A Japanese industry overview. Meat Sci. 2016, 120, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Kruk, Z.A.; Bottema, M.J.; Reyes-Veliz, L.; Forder, R.E.A.; Pitchford, W.S.; Bottema, C.D.K. Vitamin A and marbling attributes: Intramuscular fat hyperplasia effects in cattle. Meat Sci. 2018, 137, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, X.; Sun, H.; Wang, H. Long non-coding RNAs in the regulation of skeletal myogenesis andmuscle diseases. Cancer Lett. 2018, 417, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Dong, X.; Gong, X.; Kang, J.; Zhang, Y.; Quan, F. Mutation in myostatin 3′UTR promotes C2C12 myoblast proliferation and differentiation by blocking thetranslation of MSTN. Int. J. Biol. Macromol. 2020, 154, 634–643. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harbor Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [Green Version]
- Mckinsey, T.A.; Zhang, C.L.; Olson, E.N. MEF2: A calcium-dependent regulator of cell division, differentiation and death. Trends Biochem. Sci. 2002, 27, 40–47. [Google Scholar] [CrossRef]
- Brand, N.J. Myocyte enhancer factor 2(MEF2). Int. J. Biochem. Cell Biol. 1997, 29, 1467–1470. [Google Scholar] [CrossRef]
- Black, B. Transcriptional control of muscle development by myocyte enhancer factor-2 (MEF2) proteins. Annu. Rev. Cell Dev. Biol. 1998, 14, 167–196. [Google Scholar] [CrossRef]
- Akhtar, M.W.; Kim, M.S.; Adachi, M.; Morris, M.J.; Qi, X.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N.; Kavalali, E.T.; Monteggia, L.M. In vivo analysis of MEF2 transcription factors in synapse regulation and neuronal survival. PLoS ONE 2012, 7, e34863. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.F.; Miano, J.M.; Hustad, C.M.; Copeland, N.G.; Jenkins, N.A.; Olson, E.N. A Mef2 gene that generates a muscle-specific isoform via alternative mRNA splicing. Mol. Cell. Biol. 1994, 14, 1647–1656. [Google Scholar]
- Molkentin, J.D.; Olson, E.N. Combinatorial control of muscle development by basic helix-loop-helixand MADS-box transcription factors. Proc. Natl. Acad. Sci. USA 1996, 93, 9366–9373. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Carmichael, R.E.; Valois, L.S.; Wilkinson, K.A.; Henley, J.M. The transcription factor MEF2A plays a key role in the differentiation/maturation of rat neural stem cellsinto neurons. Biochem. Biophys. Res. Commun. 2018, 500, 645–649. [Google Scholar] [CrossRef]
- Edmondson, D.G.; Lyons, G.E.; Martin, J.F.; Olson, E.N. Mef2 gene expression marks the cardiac and skeletal muscle lineages during mouse embryogenesis. Development 1994, 120, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Dey, R.; Han, A.; Jayathilaka, N.; Philips, M.; Ye, J.; Chen, L. Structure of the MADS-box/MEF2 domainof MEF2A bound to DNA and its implication for myocardin recruitment. J. Mol. Biol. 2010, 397, 520–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Zhang, S.; Wu, W. Analysis of MEF2A mutations in a Chinese population with premature coronary artery disease. Genet. Test. Mol. Biomark. 2013, 17, 352–355. [Google Scholar] [CrossRef]
- Blixt, N.; Norton, A.; Zhang, A.; Aparicio, C.; Prasad, H.; Gopalakrishnan, R.; Jensen, E.D.; Mansky, K.C. Loss of myocyte enhancer factor 2 expression in osteoclasts leads to opposing skeletal phenotypes. Bone 2020, 138, 115466. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.L.; Naya, F.J. MicroRNAs in the Myocyte Enhancer Factor 2(MEF2)regulated MEG3-DIO3 Noncoding RNA Locus Promote Cardiomyocyte Proliferation by Targeting the Transcriptional Coactivator Cited2. J. Biol. Chem. 2015, 290, 23162–23172. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Nelson, B.R.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Requirement of MEF2A, C, and D for skeletal muscle regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 4109–4114. [Google Scholar] [CrossRef]
- Zhou, W.P.; Zhang, H.; Zhao, Y.X.; Liu, G.Q.; Zhang, J.Y. RNA interference of myocyte enhancer factor 2A accelerates atherosclerosis in apolipoprotein E-deficient mice. PLoS One. 2015, 10, e0121823. [Google Scholar] [CrossRef]
- Liu, C.; Yao, M.D.; Li, C.P.; Shan, K.; Yang, H.; Wang, J.J.; Liu, B.; Li, X.M.; Yao, J.; Jiang, Q.; et al. Silencing of circular RNA-ZNF609 ameliorates vascular endothelial dysfunction. Theranostics 2017, 7, 2863–2877. [Google Scholar] [CrossRef] [PubMed]
- Potthoff, M.J.; Olson, E.N. MEF2: A central regulator of diverse developmental programs. Development 2007, 134, 4131–4140. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Telese, F. Genome wide epigenetic analysis of MEF2A and MEF2C transcription factors in mouse cortical neurons. Commun. Integr. Biol. 2015, 8, e1087624. [Google Scholar] [CrossRef] [Green Version]
- Shahjahan, M. Skeletal muscle development in vertebrate animals. Asian J. Med. Biol. Res. 2015, 1, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Medrano, J.L.; Naya, F.J. The transcription factor MEF2A fine-tunes gene expression in the atrial and ventricular chambers of the adult heart. J. Biol. Chem. 2017, 292, 20975–20988. [Google Scholar] [CrossRef]
- Wang, Y.; Mei, C.; Su, X.; Wang, H.; Yang, W.; Zan, L. MEF2A regulates the MEG3-DIO3 miRNA megacluster-targeted PP2A signaling in bovine skeletal myoblast differentiation. Int. J. Mol. Sci. 2019, 20, 27–48. [Google Scholar]
- Chen, L.; Cheng, B.; Li, L.; Zhan, S.; Wang, L.; Zhong, T.; Chen, Y.; Zhang, H. The molecular characterization and temporal spatial expression of myocyte enhancer factor 2 genes in the goat and their association with myofiber traits. Gene 2015, 555, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Harashima, H.; Dissmeyer, N.; Schnittger, A. Cell cycle control across the eukaryotic kingdom. Trends Cell Biol. 2013, 23, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Yang, W.C.; Li, P.W.; Wang, H.B.; Zhang, Y.Y.; Zan, L.S. Myocyte enhancer factor 2A promotes proliferation and its inhibition attenuates myogenic differentiation via myozenin 2 in bovine skeletal muscle myoblast. PLoS ONE 2018, 13, e0196255. [Google Scholar] [CrossRef] [Green Version]
- Hamed, M.; Khilji, S.; Chen, J.; Li, Q. Stepwise acetyltransferase association and histone acetylation at the Myod1 locus during myogenic differentiation. Sci. Rep. 2013, 3, 2390. [Google Scholar] [CrossRef] [Green Version]
- Lai, Z.; Wu, F.; Li, M.; Bai, F.; Gao, Y.; Yu, J.; Li, H.; Lei, C.; Dang, R. Tissue expression profile, polymorphism of IGF1 gene and its effect on body size traits of Dezhou donkey. Gene 2021, 766, 145118. [Google Scholar] [CrossRef] [PubMed]
- Berryman, D.E.; Glad CA, M.; List, E.O.; Johannsson, G. The GH/IGF-1 axis in obesity: Pathophysiology and therapeutic considerations. Nat. Rev. Endocrinol. 2013, 9, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Lu, Y.W.; Martino, N.; Gerlach, B.D.; Lamar, J.M.; Vincent, P.A.; Adam, A.P.; Schwarz, J.J. MEF2 (Myocyte Enhancer Factor 2) is essential for endothelial homeostasis and the atheroprotective gene expression program. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 1105–1123. [Google Scholar] [CrossRef]
- Liu, B.R.; Pang, L.H.; Ji, Y.; Fang, L.; Tian, C.W.; Chen, J.; Chen, C.N.; Zhong, Y.; Ou, W.C.; Xiong, Y.J.; et al. MEF2A Is the Trigger of Resveratrol Exerting Protection on Vascular Endothelial Cell. Front. Cardiovasc. Med. 2022, 8, 775392. [Google Scholar] [CrossRef]
- Tobin, S.W.; Yang, D.; Girgis, J.; Farahzad, A.; Blais, A.; McDermott, J.C. Regulation of Hspb7 by MEF2 and AP-1: Implications for Hspb7 in muscle atrophy. J. Cell Sci. 2016, 129, 4076–4090. [Google Scholar]
- Snyder, C.M.; Rice, A.L.; Estrella, N.L.; Held, A.; Kandarian, S.C.; Naya, F.J. MEF2A regulates theMEG3-DIO3 microRNA mega-cluster to modulate WNT signaling in skeletal muscle regeneration. Development 2012, 140, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Estrella, N.L.; Desjardins, C.A.; Nocco, S.E.; Clark, A.L.; Maksimenko, Y.; Naya, F.J. MEF2 transcription factors reguLate distinct gene programs in mammalian skeletal muscledifferentiation. J. Biol. Chem. 2015, 290, 1256–1268. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Fan, C.; Topol, S.E.; Topol, E.J.; Wang, Q. Mutation of MEF2A in an inherited disorder with features of coronary artery disease. Science 2003, 302, 1578–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naya, F.J.; Black, B.L.; Wu, H.; Bassel-Duby, R.; Richardson, J.A.; Hill, J.A.; Olson, E.N. Mitochondrial deficiency and cardiac sudden death in mice lacking the MEF2A transcription factor. Nat. Med. 2002, 8, 1303–1309. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, M.; Fan, Z.; Qin, X.; Zhu, X.; Ji, H.; Qin, Y.; Wang, Q.; Wu, L. Vertebrate paralogous MEF2 genes: Origin, conservation, and evolution. PLoS ONE 2011, 6, e17334. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| sh-NC | TTCTCCGAACGTCTCACGT |
| sh-RNA1 | GCAGAACCAACTCGGATATTG |
| sh-RNA2 | GCCTCCACTGAATACCCAAAG |
| sh-RNA3 | GCAGCACCATTTAGGACAAGC |
| sh-RNA4 | GCAGTTATCTCAGGGTTCAAA |
| Gene | Accession Numbers | Primer Sequence(5′-3′) | Product Size (bp) |
|---|---|---|---|
| MEF2A | NM_001083638.2 | F: AATGAACCTCACGAAAGCAGAAC R: TTAGCACATAGGAAGTATCAGGGTC | 106 |
| CDK2 | NM_001014934.1 | F: CCTGGATGAAGATGGACG R: CTTGGAAGAAAGGGTGAG | 101 |
| CCNA2 | NM_001075123.1 | F: GCAGCCTTTCATTTAGCACTCT R: ATTGACTGTTGTGCGTGCTG | 155 |
| Bcl2 | NM_001166486.1 | F: ATGTGTGTGGAGAGCGTCAA R: ATACAGCTCCACAAAGGCGT | 138 |
| BAD | NM_001035459.2 | F: TCCCAGAGTTTGAGCAGAGTG R: TTAGCCAGTGCTTGCTGAGAC | 108 |
| MyoD1 | NM_001040478.2 | F: AACCCCAACCCGATTTACC R: CACAACAGTTCCTTCGCCTCT | 162 |
| IGF1 | NM_001077828.1 | F: TGCGGAGACAGGGGCTTTTATTTC R: AAGCAGCACTCATCCACGATTCC | 95 |
| GAPDH | NM_001034034.2 | F: TTGTGATGGGCGTGAACC R: GTCTTCTGGGTGGCAGTGAT | 169 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Ruan, Y.; Xu, J.; Shi, P.; Xu, H. Effect of Bovine MEF2A Gene Expression on Proliferation and Apoptosis of Myoblast Cells. Genes 2023, 14, 1498. https://doi.org/10.3390/genes14071498
Sun J, Ruan Y, Xu J, Shi P, Xu H. Effect of Bovine MEF2A Gene Expression on Proliferation and Apoptosis of Myoblast Cells. Genes. 2023; 14(7):1498. https://doi.org/10.3390/genes14071498
Chicago/Turabian StyleSun, Jinkui, Yong Ruan, Jiali Xu, Pengfei Shi, and Houqiang Xu. 2023. "Effect of Bovine MEF2A Gene Expression on Proliferation and Apoptosis of Myoblast Cells" Genes 14, no. 7: 1498. https://doi.org/10.3390/genes14071498
APA StyleSun, J., Ruan, Y., Xu, J., Shi, P., & Xu, H. (2023). Effect of Bovine MEF2A Gene Expression on Proliferation and Apoptosis of Myoblast Cells. Genes, 14(7), 1498. https://doi.org/10.3390/genes14071498