
Circadian Gene Variants in Diseases
Abstract
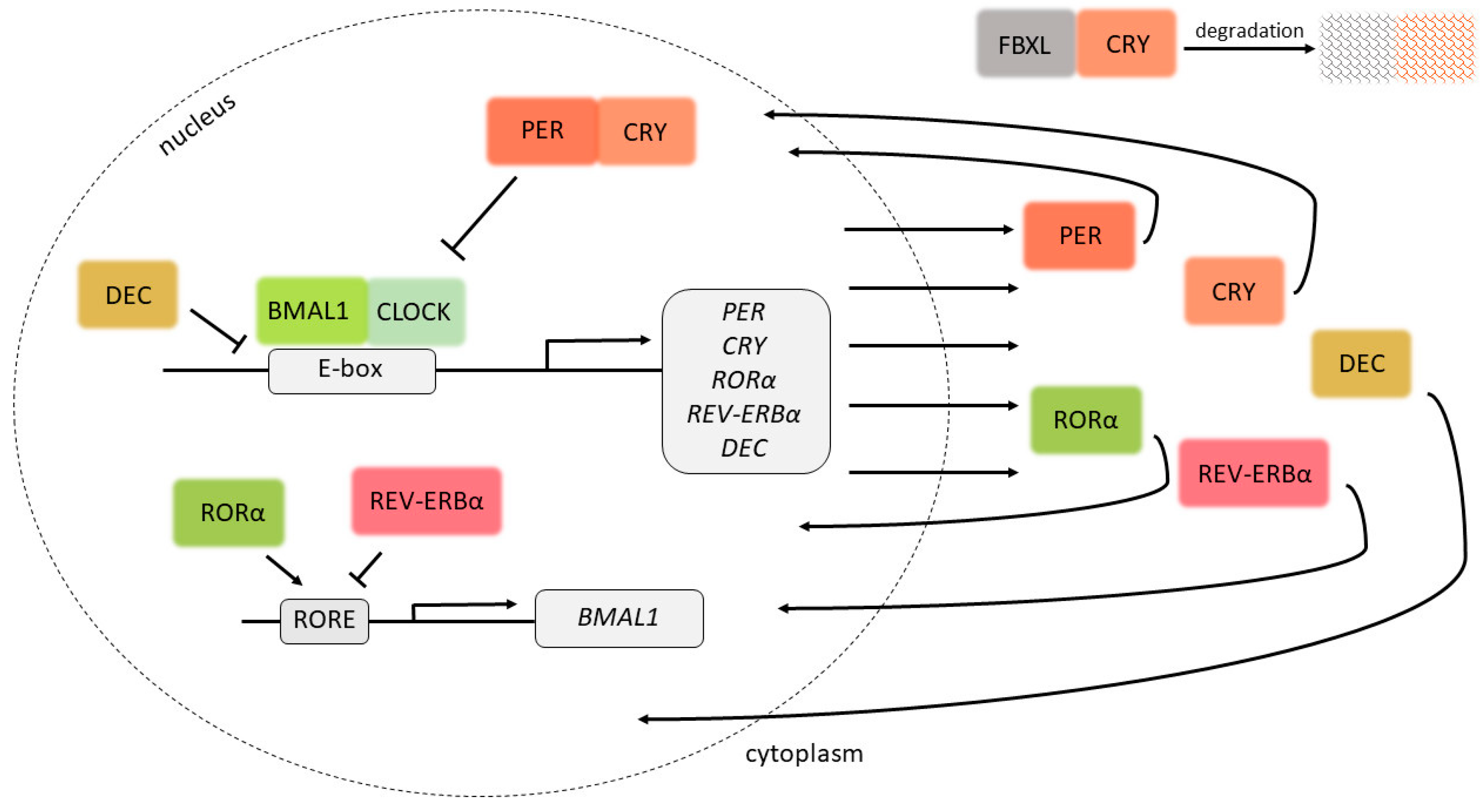
:1. Introduction
2. Circadian Gene Variants in Tumors
2.1. Associations between Circadian Gene Variants and Increased Risk for Tumor Development
2.2. Associations between Circadian Gene Variants and Increased Risk for Development of Different Tumor Subtypes
2.3. Associations between Circadian Gene Variants and Decreased Risk for Tumor Development
3. Circadian Gene Variants in Cardiovascular and Metabolic Diseases
4. Circadian Gene Variants in Sleep Disorders and Psychiatric Diseases
5. Circadian Gene Variants in Neurodegenerative Diseases
6. Circadian Gene Variants in Other Diseases
7. Circadian Genes and Immune Response
8. Future Research
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lowrey, P.L.; Takahashi, J.S. Mammalian circadian biology: Elucidating genome-wide levels of temporal organisation. Annu. Rev. Genom. Hum. Genet. 2004, 5, 407–441. [Google Scholar] [CrossRef] [PubMed]
- Harmer, S.L.; Panda, S.; Kay, S.A. Molecular bases of circadian rhythms. Annu. Rev. Cell Dev. Biol. 2001, 17, 215–253. [Google Scholar] [CrossRef]
- Yamazaki, S.; Numano, R.; Abe, M.; Hida, A.; Takahashi, R.; Ueda, M.; Block, G.D.; Sakaki, Y.; Menaker, M.; Tei, H. Resetting central and peripheral circadian oscillators in transgenic rats. Science 2000, 288, 682–685. [Google Scholar] [CrossRef]
- Ralph, M.R.; Foster, R.G.; Davis, F.C.; Menaker, M. Transplanted suprachiasmatic nucleus determines circadian period. Science 1990, 247, 975–978. [Google Scholar] [CrossRef] [PubMed]
- Sujino, M.; Masumoto, K.; Yamaguchi, S.; van der Horst, G.T.; Okamura, H.; Inouye, S.I. Suprachiasmatic nucleus grafts restore circadian behavioral rhythms of genetically arrhythmic mice. Curr. Biol. 2003, 13, 664–668. [Google Scholar] [CrossRef]
- Vitaterna, M.H.; Shimomura, K.; Jiang, P. Genetics of Circadian Rhythms. Neurol. Clin. 2019, 37, 487–504. [Google Scholar] [CrossRef] [PubMed]
- Konopka, R.J.; Benzer, S. Clock mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef]
- Bargiello, T.A.; Jackson, F.R.; Young, M.W. Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature 1984, 312, 752–754. [Google Scholar] [CrossRef]
- Zehring, W.A.; Wheeler, D.A.; Reddy, P.; Konopka, R.J.; Kyriacou, C.P.; Rosbash, M.; Hall, J.C. P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster. Cell 1984, 39, 369–376. [Google Scholar] [CrossRef]
- Albrecht, U.; Sun, Z.S.; Eichele, G.; Lee, C.C. A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell 1997, 91, 1055–1064. [Google Scholar] [CrossRef]
- Shearman, L.P.; Zylka, M.J.; Weaver, D.R.; Kolakowski, L.F., Jr.; Reppert, S.M. Two period homologs: Circadian expression and photic regulation in the suprachiasmatic nuclei. Neuron 1997, 19, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Tei, H.; Okamura, H.; Shigeyoshi, Y.; Fukuhara, C.; Ozawa, R.; Hirose, M.; Sakaki, Y. Circadian oscillation of a mammalian homologue of the Drosophila period gene. Nature 1997, 389, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Zylka, M.J.; Shearman, L.P.; Weaver, D.R.; Reppert, S.M. Three period homologs in mammals: Differential light responses in the suprachiasmatic circadian clock and oscillating transcripts outside of brain. Neuron 1998, 20, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Vitaterna, M.H.; King, D.P.; Chang, A.; Kornhauser, J.M.; Lowrey, P.L.; McDonald, J.D.; Dove, W.F.; Pinto, L.H.; Turek, F.W.; Takahashi, J.S. Mutagenesis and Mapping of a Mouse Gene, Clock, Essential for Circadian Behavior. Science 1994, 264, 719–725. [Google Scholar] [CrossRef]
- King, D.P.; Zhao, Y.; Sangoram, A.M.; Wilsbacher, L.D.; Tanaka, M.; Antoch, M.P.; Steeves, T.D.L.; Vitaterna, M.H.; Kornhauser, J.M.; Lowrey, P.L.; et al. Positional Cloning of the Mouse Circadian Clock Gene. Cell 1997, 89, 641–653. [Google Scholar] [CrossRef]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef]
- Kondratov, R.V.; Chernov, M.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Gudkov, A.V.; Antoch, M.P. BMAL1-dependent circadian oscillation of nuclear CLOCK: Posttranslational events induced by dimerization of transcriptional activators of the mammalian clock system. Genes Dev. 2003, 17, 1921–1932. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Sancar, A. Vitamin B2-based blue-light photoreceptors in the retinohypothalamic tract as the photoactive pigments for setting the circadian clock in mammals. Proc. Natl. Acad. Sci. USA 1998, 95, 6097–6102. [Google Scholar] [CrossRef]
- Kume, K.; Zylka, M.J.; Sriram, S.; Shearman, L.P.; Weaver, D.R.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M. CRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 1999, 98, 193–205. [Google Scholar] [CrossRef]
- Sato, T.K.; Panda, S.; Miraglia, L.J.; Reyes, T.M.; Rudic, R.D.; McNamara, P.; Naik, K.A.; FitzGerald, G.A.; Kay, S.A.; Hogenesch, J.B. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron 2004, 43, 527–537. [Google Scholar] [CrossRef]
- Takeda, Y.; Jothi, R.; Birault, V.; Jetten, A.M. RORγ directly regulates the circadian expression of clock genes and downstream targets in vivo. Nucleic Acids Res. 2012, 40, 8519–8535. [Google Scholar] [CrossRef] [PubMed]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef] [PubMed]
- DeBruyne, J.P.; Weaver, D.R.; Reppert, S.M. CLOCK and NPAS2 have overlapping roles in the suprachiasmatic circadian clock. Nat. Neurosci. 2007, 10, 543–545. [Google Scholar] [CrossRef]
- Siepka, S.M.; Yoo, S.; Park, J.; Song, W.; Kumar, V.; Hu, Y.; Lee, C.; Takahashi, J.S. Circadian Mutant Overtime Reveals F-box Protein FBXL3 Regulation of Cryptochrome and Period Gene Expression. Cell 2007, 129, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Hirano, A.; Yumimoto, K.; Tsunematsu, R.; Matsumoto, M.; Oyama, M.; Kozuka-Hata, H.; Nakagawa, T.; Lanjakornsiripan, D.; Nakayama, K.I.; Fukada, Y. FBXL21 regulates oscillation of the circadian clock through ubiquitination and stabilization of cryptochromes. Cell 2013, 152, 1106–1118. [Google Scholar] [CrossRef]
- Honma, S.; Kawamoto, T.; Takagi, Y.; Fujimoto, K.; Sato, F.; Noshiro, M.; Kato, Y.; Honma, K. Dec1 and Dec2 are regulators of the mammalian molecular clock. Nature 2002, 419, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Saez, L.; Delahaye-Brown, A.M.; Myers, M.P.; Sehgal, A.; Young, M.W.; Weitz, C.J. Isolation of timeless by PER protein interaction: Defective interaction between timeless protein and long-period mutant PERL. Science 1995, 270, 811–815. [Google Scholar] [CrossRef]
- Andreani, T.S.; Itoh, T.Q.; Yildirim, E.; Hwangbo, D.; Allada, R. Genetics of Circadian Rhythms. Sleep Med. Clin. 2015, 10, 413–421. [Google Scholar] [CrossRef]
- Li, H.X. The role of circadian clock genes in tumors. Onco Targets Ther. 2019, 12, 3645–3660. [Google Scholar] [CrossRef]
- Kettner, N.M.; Katchy, C.A.; Fu, L. Circadian gene variants in cancer. Ann. Med. 2014, 46, 208–220. [Google Scholar] [CrossRef]
- Morales-Santana, S.; Morell, S.; Leon, J.; Carazo-Gallego, A.; Jimenez-Lopez, J.C.; Morell, M. An Overview of the Polymorphisms of Circadian Genes Associated with Endocrine Cancer. Front. Endocrinol. 2019, 10, 104. [Google Scholar] [CrossRef]
- Bass, J.; Takahashi, J.S. Circadian Integration of Metabolism and Energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Young, M.E. The Cardiomyocyte Circadian Clock: Emerging Roles in Health and Disease. Circ. Res. 2010, 106, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jain, M.K. Circadian regulation of cardiac metabolism. J. Clin. Investig. 2021, 131, e148276. [Google Scholar] [CrossRef]
- Liu, C.; Tang, X.; Gong, Z.; Zeng, W.; Hou, Q.; Lu, R. Circadian Rhythm Sleep Disorders: Genetics, Mechanisms, and Adverse Effects on Health. Front. Genet. 2022, 13, 875342. [Google Scholar] [CrossRef] [PubMed]
- Lamont, E.W.; Legault-Coutu, D.; Cermakian, N.; Boivin, D.B. The role of circadian clock genes in mental disorders. Dialogues Clin. Neurosci. 2007, 9, 333–342. [Google Scholar] [CrossRef]
- Canever, J.B.; Queiroz, L.Y.; Soares, E.S.; de Avelar, N.C.P.; Cimarosti, H.I. Circadian rhythm alterations affecting the pathology of neurodegenerative diseases. J. Neurochem. 2023. [Google Scholar] [CrossRef]
- SNPinfo Web Server. Available online: https://snpinfo.niehs.nih.gov/ (accessed on 13 August 2023).
- dbSNP—NCBI—National Institutes of Health (NIH). Available online: https://www.ncbi.nlm.nih.gov/snp/ (accessed on 13 August 2023).
- de Bakker, P.I.; Yelensky, R.; Pe’er, I.; Gabriel, S.B.; Daly, M.J.; Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 2005, 37, 1217–1223. [Google Scholar] [CrossRef]
- Barret, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Horita, N.; Kaneko, T. Genetic model selection for a case–control study and a meta-analysis. Meta Gene 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Lesicka, M.; Jabłońska, E.; Wieczorek, E.; Pepłońska, B.; Gromadzińska, J.; Seroczyńska, B.; Kalinowski, L.; Skokowski, J.; Reszka, E. Circadian Gene Polymorphisms Associated with Breast Cancer Susceptibility. Int. J. Mol. Sci. 2019, 20, 5704. [Google Scholar] [CrossRef]
- Zhu, Y.; Brown, H.N.; Zhang, Y.; Stevens, R.G.; Zheng, T. Period3 structural variation: A circadian biomarker associated with breast cancer in young women. Cancer Epidemiol. Biomark. Prev. 2005, 14, 268–270. [Google Scholar] [CrossRef]
- Dai, H.; Zhang, L.; Cao, M.; Song, F.; Zheng, H.; Zhu, X.; Wei, Q.; Zhang, W.; Chen, K. The role of polymorphisms in circadian pathway genes in breast tumorigenesis. Breast Cancer Res. Treat. 2011, 127, 531–540. [Google Scholar] [CrossRef]
- Pham, T.T.; Lee, E.S.; Kong, S.Y.; Kim, J.; Kim, S.Y.; Joo, J.; Yoon, K.A.; Park, B. Night-shift work, circadian and melatonin pathway related genes and their interaction on breast cancer risk: Evidence from a case-control study in Korean women. Sci. Rep. 2019, 9, 10982. [Google Scholar] [CrossRef] [PubMed]
- Truong, T.; Liquet, B.; Menegaux, F.; Plancoulaine, S.; Laurent-Puig, P.; Mulot, C.; Cordina-Duverger, E.; Sanchez, M.; Arveux, P.; Kerbrat, P.; et al. Breast cancer risk, nightwork, and circadian clock gene polymorphisms. Endocr. Relat. Cancer 2014, 21, 629–638. [Google Scholar] [CrossRef]
- Zhu, Y.; Stevens, R.G.; Leaderer, D.; Hoffman, A.; Holford, T.; Zhang, Y.; Brown, H.N.; Zheng, T. Non-synonymous polymorphisms in the circadian gene NPAS2 and breast cancer risk. Breast Cancer Res. Treat. 2008, 107, 421–425. [Google Scholar] [CrossRef]
- Zhu, Y.; Stevens, R.G.; Hoffman, A.E.; FitzGerald, L.M.; Kwon, E.M.; Ostrander, E.A.; Davis, S.; Zheng, T.; Stanford, J.L. Testing the circadian gene hypothesis in prostate cancer: A population-based case-control study. Cancer Res. 2009, 69, 9315–9322. [Google Scholar] [CrossRef]
- Chu, L.; Zhu, Y.; Yu, K.; Zheng, T.; Yu, H.; Zhang, Y.; Sesterhenn, I.; Chokkalingam, A.P.; Danforth, K.N.; Shen, M.C.; et al. Variants in circadian genes and prostate cancer risk: A population-based study in China. Prostate Cancer Prostatic Dis. 2008, 11, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Couto, P.; Miranda, D.; Vieira, R.; Vilhena, A.; De Marco, L.; Bastos-Rodrigues, L. Association between CLOCK, PER3 and CCRN4L with non-small cell lung cancer in Brazilian patients. Mol. Med. Rep. 2014, 10, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, X.; Liu, P.; Quan, X.; Zheng, C.; Zhou, B. Association between three polymorphisms in BMAL1 genes and risk of lung cancer in a northeast Chinese population. DNA Cell Biol. 2019, 38, 1437–1443. [Google Scholar] [CrossRef]
- Rajendran, S.; Benna, C.; Marchet, A.; Nitti, D.; Mocellin, S. Germline polymorphisms of circadian genes and gastric cancer predisposition. Cancer Commun. 2020, 40, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.E.; Zheng, T.; Stevens, R.G.; Ba, Y.; Zhang, Y.; Leaderer, D.; Yi, C.; Holford, T.R.; Zhu, Y. Clock-Cancer Connection in Non–Hodgkin’s Lymphoma: A Genetic Association Study and Pathway Analysis of the Circadian Gene Cryptochrome 2. Cancer Res. 2009, 69, 3605–3613. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, A.E.; Zheng, T.; Yi, C.H.; Stevens, R.G.; Ba, Y.; Zhang, Y.; Leaderer, D.; Holford, T.; Hansen, J.; Zhu, Y. The core circadian gene cryptochrome 2 influences breast cancer risk, possibly by mediating hormone signalling. Cancer Prev. Res. 2010, 3, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Wirth, M.D.; Burch, J.B.; Hébert, J.R.; Kowtal, P.; Kapoor, A.; Steck, S.E.; Hurley, T.G.; Gupta, P.C.; Pednekar, M.S.; Youngstedt, S.D.; et al. Case-control study of breast cancer in India: Role of PERIOD3 clock gene length polymorphism and chronotype. Cancer Invest. 2014, 32, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Fores-Martos, J.; Cervera-Vidal, R.; Sierra-Roca, J.; Lozano-Asencio, C.; Fedele, V.; Cornelissen, S.; Edvarsen, H.; Tadeo-Cervera, I.; Eroles, P.; Lluch, A.; et al. Circadian PERformance in breast cancer: A germline and somatic genetic study of PER3VNTR polymorphisms and gene co-expression. NPJ Breast Cancer 2021, 7, 118. [Google Scholar] [CrossRef]
- Monsees, G.M.; Kraft, P.; Hankinson, S.E.; Hunter, D.J.; Schernhammer, E.S. Circadian genes and breast cancer susceptibility in rotating shift workers. Int. J. Cancer 2012, 131, 2547–2552. [Google Scholar] [CrossRef]
- Yu, C.C.; Chen, L.C.; Chiou, C.Y.; Chang, Y.J.; Lin, V.C.; Huang, C.Y.; Lin, I.L.; Chang, T.Y.; Lu, T.L.; Lee, C.H.; et al. Genetic variants in the circadian rhythm pathway as indicators of prostate cancer progression. Cancer Cell Int. 2019, 19, 87. [Google Scholar] [CrossRef]
- Wendeu-Foyet, M.G.; Koudou, Y.; Cénée, S.; Trétarre, B.; Rébillard, X.; Cancel-Tassin, G.; Cussenot, O.; Boland, A.; Bacq, D.; Deleuze, J.F.; et al. Circadian genes and risk of prostate cancer: Findings from the EPICAP study. Int. J. Cancer 2019, 145, 1745–1753. [Google Scholar] [CrossRef]
- Markt, S.C.; Valdimarsdottir, U.A.; Shui, I.M.; Sigurdardottir, L.G.; Rider, J.R.; Tamimi, R.M.; Batista, J.L.; Haneuse, S.; Flynn-Evans, E.; Lockley, S.W.; et al. Circadian clock genes and risk of fatal prostate cancer. Cancer Causes Control 2015, 26, 25–33. [Google Scholar] [CrossRef]
- Rabstein, S.; Harth, V.; Justenhoven, C.; Pesch, B.; Plöttner, S.; Heinze, E.; Lotz, A.; Baisch, C.; Schiffermann, M.; Brauch, H.; et al. Polymorphisms in circadian genes, night work and breast cancer: Results from the GENICA study. Chronobiol. Int. 2014, 31, 1115–1122. [Google Scholar] [CrossRef]
- Fu, A.; Leaderer, D.; Zheng, T.; Hoffman, A.E.; Stevens, R.G.; Zhu, Y. Genetic and Epigenetic Associations of Circadian Gene TIMELESS and Breast Cancer Risk. Mol. Carcinog. 2012, 51, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Chang-Claude, J.; Critchley, A.M.; Kyriacou, C.; Lavers, S.; Rattay, T.; Seibold, P.; Webb, A.; West, C.; Symonds, R.P.; et al. Genetic Variants Predict Optimal Timing of Radiotherapy to Reduce Side-effects in Breast Cancer Patients. Clin. Oncol. 2019, 31, 9–16. [Google Scholar] [CrossRef]
- Webb, A.J.; Harper, E.; Rattay, T.; Aguado-Barrera, M.E.; Azria, D.; Bourgier, C.; Brengues, M.; Briers, E.; Bultijnck, R.; Chang-Claude, J.; et al. Treatment time and circadian genotype interact to influence radiotherapy side-effects. A prospective European validation study using the REQUITE cohort. EBioMedicine 2022, 84, 104269. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.W.; Zhu, Y.; Yu, K.; Zheng, T.; Chokkalingam, A.P.; Stanczyk, F.Z.; Gao, Y.T.; Hsing, A.W. Correlation between Circadian Gene Variants and Serum Levels of Sex Steroids and Insulin-like Growth Factor-I. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Leu, H.B.; Chung, C.M.; Lin, S.J.; Chiang, K.M.; Yang, H.C.; Ho, H.Y.; Ting, C.T.; Lin, T.H.; Sheu, S.H.; Tsai, W.C.; et al. Association of circadian genes with diurnal blood pressure changes and non-dipper essential hypertension: A genetic association with young-onset hypertension. Hypertens. Res. 2015, 38, 155–162. [Google Scholar] [CrossRef]
- Garaulet, M.; Lee, Y.C.; Shen, J.; Parnell, L.D.; Arnett, D.K.; Tsai, M.Y.; Lai, C.Q.; Ordovas, J.M. CLOCK genetic variation and metabolic syndrome risk: Modulation by monounsaturated fatty acids. Am. J. Clin. Nutr. 2009, 90, 1466–1475. [Google Scholar] [CrossRef]
- Garaulet, M.; Lee, Y.C.; Shen, J.; Parnell, L.D.; Arnett, D.K.; Tsai, M.Y.; Lai, C.Q.; Ordovas, J.M. Genetic variants in human CLOCK associate with total energy intake and cytokine sleep factors in overweight subjects (GOLDN population). Eur. J. Hum. Genet. 2010, 18, 364–369. [Google Scholar] [CrossRef]
- Corella, D.; Asensio, E.M.; Coltell, O.; Sorlí, J.V.; Estruch, R.; Martínez-González, M.A.; Salas-Salvadó, J.; Castañer, O.; Arós, F.; Lapetra, J.; et al. CLOCK gene variation is associated with incidence of type-2 diabetes and cardiovascular diseases in type-2 diabetic subjects: Dietary modulation in the PREDIMED randomized trial. Cardiovasc. Diabetol. 2016, 15, 4. [Google Scholar] [CrossRef]
- Uemura, H.; Katsuura-Kamano, S.; Yamaguchi, M.; Arisawa, K.; Hamajima, N.; Hishida, A.; Kawai, S.; Oze, I.; Shinchi, K.; Takashima, N.; et al. Variant of the clock circadian regulator (CLOCK) gene and related haplotypes are associated with the prevalence of type 2 diabetes in the Japanese population. J. Diabetes 2016, 8, 667–676. [Google Scholar] [CrossRef]
- Škrlec, I.; Talapko, J.; Džijan, S.; Lepeduš, H. The Association of Cardiovascular Disease with the T3111C Polymorphism in the CLOCK Gene. In Proceedings of the The 1st International Electronic Conference on Medicine, online, 20–30 June 2021. Med. Sci. Forum 2021, 6, 1. [Google Scholar] [CrossRef]
- Sookoian, S.; Gemma, C.; Gianotti, T.F.; Burgueño, A.; Castaño, G.; Pirola, C.J. Genetic variants of Clock transcription factor are associated with individual susceptibility to obesity. Am. J. Clin. Nutr. 2008, 87, 1606–1615. [Google Scholar] [CrossRef]
- Schroor, M.M.; Plat, J.; Mensink, R.P. Relation between single nucleotide polymorphisms in circadian clock relevant genes and cholesterol metabolism. Mol. Genet. Metab. 2023, 138, 107561. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milić, J.; Steiner, R. The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction. J. Clin. Med. 2020, 9, 484. [Google Scholar] [CrossRef]
- Škrlec, I.; Talapko, J.; Džijan, S.; Cesar, V.; Lazić, N.; Lepeduš, H. The Association between Circadian Clock Gene Polymorphisms and Metabolic Syndrome: A Systematic Review and Meta-Analysis. Biology 2022, 11, 20. [Google Scholar] [CrossRef]
- Dashti, H.S.; Smith, C.E.; Lee, Y.C.; Parnell, L.D.; Lai, C.Q.; Arnett, D.K.; Ordovás, J.M.; Garaulet, M. CRY1 circadian gene variant interacts with carbohydrate intake for insulin resistance in two independent populations: Mediterranean and North American. Chronobiol. Int. 2014, 31, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Smith, C.E.; Gomez-Abellán, P.; Ordovás-Montañés, M.; Lee, Y.C.; Parnell, L.D.; Arnett, D.K.; Ordovás, J.M. REV-ERB-ALPHA circadian gene variant associates with obesity in two independent populations: Mediterranean and North American. Mol. Nutr. Food Res. 2014, 58, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Tortorella, A.; Docimo, L.; Maldonato, M.N.; Canestrelli, B.; De Luca, L.; Maj, M. Investigation of 3111T/C polymorphism of the CLOCK gene in obese individuals with or without binge eating disorder: Association with higher body mass index. Neurosci. Lett. 2008, 435, 30–33. [Google Scholar] [CrossRef]
- Garaulet, M.; Corbalán, M.D.; Madrid, J.A.; Morales, E.; Baraza, J.C.; Lee, Y.C.; Ordovas, J.M. CLOCK gene is implicated in weight reduction in obese patients participating in a dietary programme based on the Mediterranean diet. Int. J. Obes. 2010, 34, 516–523. [Google Scholar] [CrossRef]
- Li, G.Y.; Wang, H.; Chen, H. Association of insulin resistance with polymorphic variants of Clock and Bmal1 genes: A case-control study. Clin. Exp. Hypertens. 2020, 42, 371–375. [Google Scholar] [CrossRef]
- Torrego-Ellacuría, M.; Barabash, A.; Matía-Martín, P.; Sánchez-Pernaute, A.; Torres, A.J.; Calle-Pascual, A.L.; Rubio-Herrera, M.A. Influence of CLOCK Gene Variants on Weight Response after Bariatric Surgery. Nutrients 2022, 14, 3472. [Google Scholar] [CrossRef]
- Riestra, P.; Gebreab, S.Y.; Xu, R.; Khan, R.J.; Gaye, A.; Correa, A.; Min, N.; Sims, M.; Davis, S.K. Circadian CLOCK gene polymorphisms in relation to sleep patterns and obesity in African Americans: Findings from the Jackson heart study. BMC Genet. 2017, 18, 58. [Google Scholar] [CrossRef]
- Molina-Montes, E.; Rodríguez-Barranco, M.; Ching-Lopez, A.; Artacho, R.; Huerta, J.M.; Amiano, P.; Lasheras, C.; Moreno-Iribas, C.; Jimenez-Zabala, A.; Chirlaque, M.D.; et al. Circadian clock gene variants and their link with chronotype, chrononutrition, sleeping patterns and obesity in the European prospective investigation into cancer and nutrition (EPIC) study. Clin. Nutr. 2022, 41, 1977–1990. [Google Scholar] [CrossRef] [PubMed]
- Woon, P.Y.; Kaisaki, P.J.; Bragança, J.; Bihoreau, M.T.; Levy, J.C.; Farrall, M.; Gauguier, D. Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef]
- Mirzaei, K.; Xu, M.; Qi, Q.; de Jonge, L.; Bray, G.A.; Sacks, F.; Qi, L. Variants in glucose- and circadian rhythm–related genes affect the response of energy expenditure to weight-loss diets: The POUNDS LOST Trial. Am. J. Clin. Nutr. 2014, 99, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Kovanen, L.; Donner, K.; Kaunisto, M.; Partonen, T. CRY1, CRY2 and PRKCDBP genetic variants in metabolic syndrome. Hypertens. Res. 2015, 38, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Mariman, E.C.M.; Bouwman, F.G.; Aller, E.E.J.G.; van Baak, M.A.; Wang, P. Extreme obesity is associated with variation in genes related to the circadian rhythm of food intake and hypothalamic signalling. Physiol. Genom. 2015, 47, 225–231. [Google Scholar] [CrossRef]
- Garaulet, M.; Corbalán-Tutau, M.D.; Madrid, J.A.; Baraza, J.C.; Parnell, L.D.; Lee, Y.C.; Ordovas, J.M. PERIOD2 Variants Are Associated with Abdominal Obesity, Psycho-Behavioral Factors, and Attrition in the Dietary Treatment of Obesity. J. Am. Diet. Assoc. 2010, 110, 917–921. [Google Scholar] [CrossRef]
- Katzenberg, D.; Young, Y.; Finn, L.; Lin, L.; King, D.P.; Takahashi, J.S.; Mignot, E. A CLOCK polymorphism associated with human diurnal preference. Sleep 1998, 21, 569–576. [Google Scholar] [CrossRef]
- Johansson, C.; Willeit, M.; Smedh, C.; Ekholm, J.; Paunio, T.; Kieseppä, T.; Lichtermann, D.; Praschak-Rieder, N.; Neumeister, A.; Nilsson, L.G.; et al. Circadian clock-related polymorphisms in seasonal affective disorder and their relevance to diurnal preference. Neuropsychopharmacology 2003, 28, 734–739. [Google Scholar] [CrossRef]
- Archer, S.N.; Robilliard, D.L.; Skene, D.J.; Smits, M.; Williams, A.; Arendt, J.; von Schantz, M. A length polymorphism in the circadian clock gene Per3 is linked to delayed sleep phase syndrome and extreme diurnal preference. Sleep 2003, 26, 413–415. [Google Scholar] [CrossRef]
- Pereira, D.S.; Tufik, S.; Louzada, F.M.; Benedito-Silva, A.A.; Lopez, A.R.; Lemos, N.A.; Korczak, A.L.; D’Almeida, V.; Pedrazzoli, M. Association of the length polymorphism in the human Per3 gene with the delayed sleep-phase syndrome: Does latitude have an influence upon it? Sleep 2005, 28, 29–32. [Google Scholar] [CrossRef]
- Peng, X.; Li, J.; Han, B.; Zhu, Y.; Cheng, D.; Li, Q.; Du, J. Association of occupational stress, period circadian regulator 3 (PER3) gene polymorphism and their interaction with poor sleep quality. J. Sleep Res. 2022, 31, e13390. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, T.; Hida, A.; Shimada, M.; Uehara, C.; Nishino, Y.; Kadotani, H.; Uchiyama, M.; Ebisawa, T.; Inoue, Y.; Kamei, Y.; et al. A missense variant in PER2 is associated with delayed sleep-wake phase disorder in a Japanese population. J. Hum. Genet. 2019, 64, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Mishima, K.; Tozawa, T.; Satoh, K.; Saitoh, H.; Mishima, Y. The 3111T/C polymorphism of hClock is associated with evening preference and delayed sleep timing in a Japanese population sample. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 133B, 101–104. [Google Scholar] [CrossRef]
- Patke, A.; Murphy, P.J.; Onat, O.E.; Krieger, A.C.; Özçelik, T.; Campbell, S.S.; Young, M.W. Mutation of the Human Circadian Clock Gene CRY1 in Familial Delayed Sleep Phase Disorder. Cell 2017, 169, 203–215.e13. [Google Scholar] [CrossRef]
- Takano, A.; Uchiyama, M.; Kajimura, N.; Mishima, K.; Inoue, Y.; Kamei, Y.; Kitajima, T.; Shibui, K.; Katoh, M.; Watanabe, T.; et al. A missense variation in human casein kinase I epsilon gene that induces functional alteration and shows an inverse association with circadian rhythm sleep disorders. Neuropsychopharmacology 2004, 29, 1901–1909. [Google Scholar] [CrossRef]
- Toh, K.L.; Jones, C.R.; He, Y.; Eide, E.J.; Hinz, W.A.; Virshup, D.M.; Ptácek, L.J.; Fu, Y.H. An hPer2 phosphorylation site mutation in familial advanced sleep phase syndrome. Science 2001, 291, 1040–1043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hirano, A.; Hsu, P.K.; Jones, C.R.; Sakai, N.; Okuro, M.; McMahon, T.; Yamazaki, M.; Xu, Y.; Saigoh, N.; et al. A PERIOD3 variant causes a circadian phenotype and is associated with a seasonal mood trait. Proc. Natl. Acad. Sci. USA 2016, 113, E1536–E1544. [Google Scholar] [CrossRef]
- Hirano, A.; Shi, G.; Jones, C.R.; Lipzen, A.; Pennacchio, L.A.; Xu, Y.; Hallows, W.C.; McMahon, T.; Yamazaki, M.; Ptáček, L.J.; et al. A Cryptochrome 2 mutation yields advanced sleep phase in humans. eLife 2016, 5, e16695. [Google Scholar] [CrossRef]
- Kurien, P.; Hsu, P.K.; Leon, J.; Wu, D.; McMahon, T.; Shi, G.; Xu, Y.; Lipzen, A.; Pennacchio, L.A.; Jones, C.R.; et al. TIMELESS mutation alters phase responsiveness and causes advanced sleep phase. Proc. Natl. Acad. Sci. USA 2019, 116, 12045–12053. [Google Scholar] [CrossRef]
- Xu, Y.; Padiath, Q.S.; Shapiro, R.E.; Jones, C.R.; Wu, S.C.; Saigoh, N.; Saigoh, K.; Ptácek, L.J.; Fu, Y.H. Functional consequences of a CKIδ mutation causing familial advanced sleep phase syndrome. Nature 2005, 434, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Hida, A.; Kitamura, S.; Katayose, Y.; Kato, M.; Ono, H.; Kadotani, H.; Uchiyama, M.; Ebisawa, T.; Inoue, Y.; Kamei, Y.; et al. Screening of Clock Gene Polymorphisms Demonstrates Association of a PER3 Polymorphism with Morningness–Eveningness Preference and Circadian Rhythm Sleep Disorder. Sci. Rep. 2014, 4, 6309. [Google Scholar] [CrossRef] [PubMed]
- Robilliard, D.L.; Archer, S.N.; Arendt, J.; Lockley, S.W.; Hack, L.M.; English, J.; Leger, D.; Smits, M.G.; Williams, A.; Skene, D.J.; et al. The 3111 Clock gene polymorphism is not associated with sleep and circadian rhythmicity in phenotypically characterized human subjects. J. Sleep Res. 2002, 11, 305–312. [Google Scholar] [CrossRef]
- Iwase, T.; Kajïmura, N.; Uchiyama, M.; Ebisawa, T.; Yoshimura, K.; Kamei, Y.; Shibui, K.; Kim, K.; Kudo, Y.; Katoh, M.; et al. Mutation screening of the human Clock gene in circadian rhythm sleep disorders. Psychiatry Res. 2002, 109, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Serretti, A.; Benedetti, F.; Mandelli, L.; Lorenzi, C.; Pirovano, A.; Colombo, C.; Smeraldi, E. Genetic dissection of psychopathological symptoms: Insomnia in mood disorders and CLOCK gene polymorphism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2003, 121B, 39–43. [Google Scholar] [CrossRef]
- Benedetti, F.; Serretti, A.; Colombo, C.; Barbini, B.; Lorenzi, C.; Campori, E.; Smeraldi, E. Influence of CLOCK gene polymorphism on circadian mood fluctuation and illness recurrence in bipolar depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2003, 123B, 23–26. [Google Scholar] [CrossRef]
- Benedetti, F.; Dallaspezia, S.; Fulgosi, M.C.; Lorenzi, C.; Serretti, A.; Barbini, B.; Colombo, C.; Smeraldi, E. Actimetric evidence that CLOCK 3111 T/C SNP influences sleep and activity patterns in patients affected by bipolar depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2007, 144B, 631–635. [Google Scholar] [CrossRef]
- Geoffroy, P.E.; Lajnef, M.; Bellivier, F.; Jamain, S.; Gard, S.; Kahn, J.-P.; Henry, C.; Leboyer, M.; Etain, B. Genetic association study of circadian genes with seasonal pattern in bipolar disorders. Sci. Rep. 2015, 5, 10232. [Google Scholar] [CrossRef]
- Shi, S.-Q.; White, M.J.; Borsetti, H.M.; Pendergast, J.S.; Hida, A.; Ciarleglio, C.M.; de Verteuil, P.A.; Cadar, A.G.; Cala, C.; McMahon, D.G.; et al. Molecular analyses of circadian gene variants reveal sex-dependent links between depression and clocks. Transl. Psychiatry 2016, 6, e748. [Google Scholar] [CrossRef]
- Schuch, J.B.; Genro, J.P.; Bastos, C.R.; Ghisleni, G.; Tovo-Rodrigues, L. The role of CLOCK gene in psychiatric disorders: Evidence from human and animal research. Am. J. Med. Genet. Part B 2018, 177B, 181–198. [Google Scholar] [CrossRef]
- Xu, Y.; Ma, H.; Zhao, t.; Wen, D.; Wen, Y.; Qiao, D.; Liu, Z. Association Between Period 3 Gene Polymorphisms and Adverse Effects of Antidepressants for Major Depressive Disorder. Genet. Test. Mol. Biomark. 2019, 23, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Serretti, A.; Cusin, C.; Benedetti, F.; Mandelli, L.; Pirovano, A.; Zanardi, R.; Colombo, C.; Smeraldi, E. Insomnia improvement during antidepressant treatment and CLOCK gene polymorphism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 137B, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Gyorik, D.; Eszlari, N.; Gal, Z.; Torok, D.; Baksa, D.; Kristof, Z.; Sutori, S.; Petschner, P.; Juhasz, G.; Bagdy, G.; et al. Every Night and Every Morn: Effect of Variation in CLOCK Gene on Depression Depends on Exposure to Early and Recent Stress. Front. Psychiatry 2021, 12, 687487. [Google Scholar] [CrossRef] [PubMed]
- Bacalini, M.G.; Palombo, F.; Garagnani, P.; Giuliani, C.; Fiorini, C.; Caporali, L.; Stanzani Maserati, M.; Capellari, S.; Romagnoli, M.; De Fanti, S.; et al. Association of rs3027178 polymorphism in the circadian clock gene PER1 with susceptibility to Alzheimer’s disease and longevity in an Italian population. GeroScience 2022, 44, 881–896. [Google Scholar] [CrossRef]
- Bessi, V.; Balestrini, J.; Bagnoli, S.; Mazzeo, S.; Giacomucci, G.; Padiglioni, S.; Piaceri, I.; Carraro, M.; Ferrari, C.; Bracco, L.; et al. Influence of ApoE Genotype and Clock T3111C Interaction with Cardiovascular Risk Factors on the Progression to Alzheimer’s Disease in Subjective Cognitive Decline and Mild Cognitive Impairment Patients. J. Pers. Med. 2020, 10, 45. [Google Scholar] [CrossRef]
- Lavtar, P.; Rudolf, G.; Maver, A.; Hodžić, A.; Starčević Čizmarević, N.; Živković, M.; Šega Jazbec, S.; Klemenc Ketiš, Z.; Kapović, M.; Dinčić, E.; et al. Association of circadian rhythm genes ARNTL/BMAL1 and CLOCK with multiple sclerosis. PLoS ONE 2018, 13, e0190601. [Google Scholar] [CrossRef]
- de Rojas, I.; Martin-Montero, C.; Fedetz, M.; González-Jiménez, A.; Matesanz, F.; Urcelay, E.; Espino-Paisán, L. Polymorphisms in ARNTL/BMAL1 and CLOCK Are Not Associated with Multiple Sclerosis in Spanish Population. Biology 2022, 11, 1417. [Google Scholar] [CrossRef]
- Gu, Z.; Wang, B.; Zhang, Y.B.; Ding, H.; Zhang, Y.; Yu, J.; Gu, M.; Chan, P.; Cai, Y. Association of ARNTL and PER1 genes with Parkinson’s disease: A case-control study of han chinese. Sci. Rep. 2015, 5, 15891. [Google Scholar] [CrossRef]
- Chen, H.; Ding, X.; Ding, E.; Chen, M.; Wang, H.; Yang, G.; Zhu, B. A missense variant rs2585405 in clock gene PER1 is associated with the increased risk of noise-induced hearing loss in a Chinese occupational population. BMC Med. Genom. 2021, 14, 221. [Google Scholar] [CrossRef]
- Curtis, A.M.; Bellet, M.M.; Sassone-Corsi, P.; O’Neill, L.A.J. Circadian clock proteins and immunity. Immunity 2014, 40, 178–186. [Google Scholar] [CrossRef]
- Poole, J.; Kitchen, G. Circadian regulation of innate immunity in animals and humans and implications for human disease. Semin. Immunopathol. 2022, 44, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Downton, P.; Early, J.O.; Gibbs, J.E. Circadian rhythms in adaptive immunity. Immunology 2020, 161, 268–277. [Google Scholar] [CrossRef]
- Zhang, Z.; Zeng, P.; Gao, W.; Zhou, Q.; Feng, T.; Tian, X. Circadian clock: A regulator of the immunity in cancer. Cell Commun. Signal. 2021, 19, 37. [Google Scholar] [CrossRef] [PubMed]
- Awuah, W.A.; Huang, H.; Kalmanovich, J.; Mehta, A.; Mikhailova, T.; Ng, J.C.; Abdul-Rahman, T.; Adebusoye, F.T.; Tan, J.K.; Kamanousa, K.; et al. Circadian rhythm in systemic autoimmune conditions: Potential of chrono-immunology in clinical practice: A narrative review. Medicine 2023, 102, e34614. [Google Scholar] [CrossRef]
- Xiang, H.; Xu, Z.; Hu, Y.-Q.; He, Y.-S.; Wu, G.-C.; Li, T.-Y.; Wang, X.-R.; Ding, L.-H.; Zhang, Q.; Tao, S.-S.; et al. Circadian clock genes as promising therapeutic targets for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102866. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tumor Type | Gene | SNP ID | Genotype | Population | Reference |
|---|---|---|---|---|---|
| Breast cancer | PER1 | rs2735611 | AA RM | Caucasian (Polish) | [43] |
| PER2 | rs934945 | CT + TT DM | Caucasian (Polish) | [43] | |
| PER3 | rs57875989 | 4/5 + 5/5 VNTR DM | Caucasian | [44] | |
| CLOCK | rs3805151 | CT + TT DM | Chinese | [45] | |
| rs11133373 | CG + GG DM | Korean | [46] | ||
| CRY2 | rs10838524 | AG + GG DM | Caucasian (Polish) | [43] | |
| RORA | rs1482057 | French from two administrative areas | [47] | ||
| rs12914272 | French from two administrative areas | [47] | |||
| NPAS2 | rs2305160 | AG | predominantly Caucasian | [48] | |
| Prostate cancer | BMAL1 | rs7950226 | GA + AA DM | Caucasian | [49] |
| CRY2 | rs1401417 | GC + CC DM | Chinese | [50] | |
| NPAS2 | rs1369481 | GA + AA DM | Caucasian | [49] | |
| rs895521 | GA + AA DM | Caucasian | [49] | ||
| rs17024926 | TC + CC DM | Caucasian | [49] | ||
| CSNK1E | rs1534891 | TT | Caucasian | [49] | |
| Lung cancer | PER3 | rs228729 | GT + GG DM | Brazilian | [51] |
| BMAL1 | rs3816360 | CC | northeast Chinese | [52] | |
| rs2290035 | AA | northeast Chinese | [52] | ||
| Gastric cancer | NPAS2 | rs895520 | AA RM | Caucasian | [53] |
| Non-Hodgkin lymphoma/B-cell lymphoma | CRY2 | rs11038689 | GG RM | predominantly Caucasian | [54] |
| rs7123390 | AA RM | predominantly Caucasian | [54] | ||
| rs1401417 | CC RM | predominantly Caucasian | [54] |
| Gene | Variant | Genotype | Phenotype | Population | Reference |
|---|---|---|---|---|---|
| PER1 | rs2585405 | GC + CC | Lower 3α-diol, higher SHBG | Chinese | [66] |
| PER2 | rs6431590 | AG + AA | Lack of overnight blood pressure decrease | Chinese | [67] |
| PER3 | rs57875989 | 4/5 + 5/5 VNTR | Increased serum IGF-I levels andIGF-I:IGFBP3 ratio | Chinese | [66] |
| CLOCK | rs4580704 | CG + GG DM | Lower blood pressure, higher serum concentrations of MCP1 and adiponectin, lower type 2 diabetes risk | Caucasian or predominantly Caucasian | [68,69,70] |
| rs1801260 (3111T/C) | GG RM | Higher fasting insulin, higher HOMA-IR | Caucasian | [68] | |
| CT + CC | Higher oddsratio for the prevalence of diabetes | Japanese | [71] | ||
| Higher risk for cardiovascular disease | multiple | [72] | |||
| rs1554483 | GC + GG | Overweight, obesity | Caucasian | [73] | |
| rs4864548 | AG + AA | Overweight, obesity | Caucasian | [73] | |
| rs13113518 | CT AM | Higher campesterol levels | Caucasian | [74] | |
| rs35115774 | C- AM | Lower campesterol and sitosterol levels | Caucasian | [74] | |
| rs6832769 | AG AM | Lower campesterol and sitosterol levels | Caucasian | [74] | |
| rs3749474 | TC + TT DM | Higher energy intake, decreased serum levels of IL-6 and MCP1 | predominantly Caucasian | [69] | |
| rs6811520 | TT RM | Higher incidence of myocardial infarction | Caucasian (Croatian) | [75] | |
| rs13124436 | GG RM | Higher incidence of myocardial infarction | Caucasian (Croatian) | [75] | |
| BMAL1 | rs6486121 | TC + CC DM | Higher campesterol levels | Caucasian | [74] |
| rs3789327 | AG + GG DM | Lower incidence of myocardial infarction | Caucasian (Croatian) | [75] | |
| rs12363415 | AG + GG DM | Lower incidence of myocardial infarction | Caucasian (Croatian) | [75] | |
| rs3816358 | TT + TG | Lack of overnight blood pressure decrease | Chinese | [67] | |
| rs7950226 | GG RM | Lower risk for MetS comorbidities | multiple | [76] | |
| CRY1 | rs2078074 | CC RM | Higher sitosterol levels | Caucasian | [74] |
| rs2287161 | CC | Higher carbohydrate intake | Spanish and Northamerican (predominatly Caucasian) | [77] | |
| RORββ | rs1410225 | TT | Presence of overnight blood pressure decrease | Chinese | [67] |
| RORα | rs10519096 | AG + AA | Lack of overnight blood pressure decrease | Chinese | [67] |
| REV-ERBα | rs2314339 | AG + AA | Lower probability of abdominal obesity, more physical activity | Spanish and Northamerican (predominatly Caucasian) | [78] |
| NPAS2 | rs3888170 | CT + CC | Lack of overnight blood pressure decrease | Chinese | [67] |
| rs2305160 | GA + AA | Decreased levels of free and bioavailable testosterone | Chinese | [66] |
| Disorder | Gene | Mutation | Modification | Phenotype | Population | Reference |
|---|---|---|---|---|---|---|
| (F)DSPD | PER2 | G−>A | Val1205Met | sleep–wake phase delay, idiopathic hypersomnia | Japanese | [95] |
| PER3 | 4 VNTR | / | association with evening preference | [92] | ||
| 5 VNTR | / | association with morning preference | [92] | |||
| delayed sleep phase, association with diurnal preference | predominantly Caucasian | [93] | ||||
| CLOCK | 3111T/C | / | evening preference, significantly delayed sleep onset, shorter sleep time and greater daytime sleepiness in CC hmozygotes | Japanese | [96] | |
| CRY1 | A−>C | ∆ exon 11 | enhanced interaction with CLOCK and BMAL1, long-period behavioral and body temperature rhythms with diminished amplitudes | [97] | ||
| CKIɛ | G−>A | Ser408Asp | protective effect of A allele against DSPD | Japanese | [98] | |
| (F)ASPD | PER2 | A−>G | Ser662Gly | advance of sleep, temperature, and melatonin rhythms | A single family with ASPD | [99] |
| PER3 | C−>G | Pro415Ala | habitual early spontaneous awakening | A single family with ASPD | [100] | |
| A−>G | His417Arg | habitual early spontaneous awakening | A single family with ASPD | [100] | ||
| CRY2 | G−>A | Ala260Thr | alternation of CRY2 conformation, which results in increase in accessibility and affinity for an E3 ubiquitin ligase FBXL3 and consequently CRY2 degradation | A single family with ASPD | [101] | |
| TIMELESS | C−>T | Arg1081X | destabilization of CRY1/2 and PER1/2 heterodimer, a shortened circadian period or altered entrainment | A single family with ASPD | [102] | |
| CKIδ | A−>G | Thr44Ala | a shorter circadian period | A single family with ASPD | [103] | |
| FRT | PER3 | C−>G | Pro864Ala | G allele is more common in evening types and in FRT individuals | Japanese | [104] |
| N-24 | CKIɛ | G−>A | Ser408Asp | protective effect of A allele against N-24 | Japanese | [98] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gršković, P.; Korać, P. Circadian Gene Variants in Diseases. Genes 2023, 14, 1703. https://doi.org/10.3390/genes14091703
Gršković P, Korać P. Circadian Gene Variants in Diseases. Genes. 2023; 14(9):1703. https://doi.org/10.3390/genes14091703
Chicago/Turabian StyleGršković, Paula, and Petra Korać. 2023. "Circadian Gene Variants in Diseases" Genes 14, no. 9: 1703. https://doi.org/10.3390/genes14091703
APA StyleGršković, P., & Korać, P. (2023). Circadian Gene Variants in Diseases. Genes, 14(9), 1703. https://doi.org/10.3390/genes14091703