
Transcriptome Analysis and Reactive Oxygen Species Detection Suggest Contrasting Molecular Mechanisms in Populus canadensis’ Response to Different Formae Speciales of Marssonina brunnea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Host and Pathogen
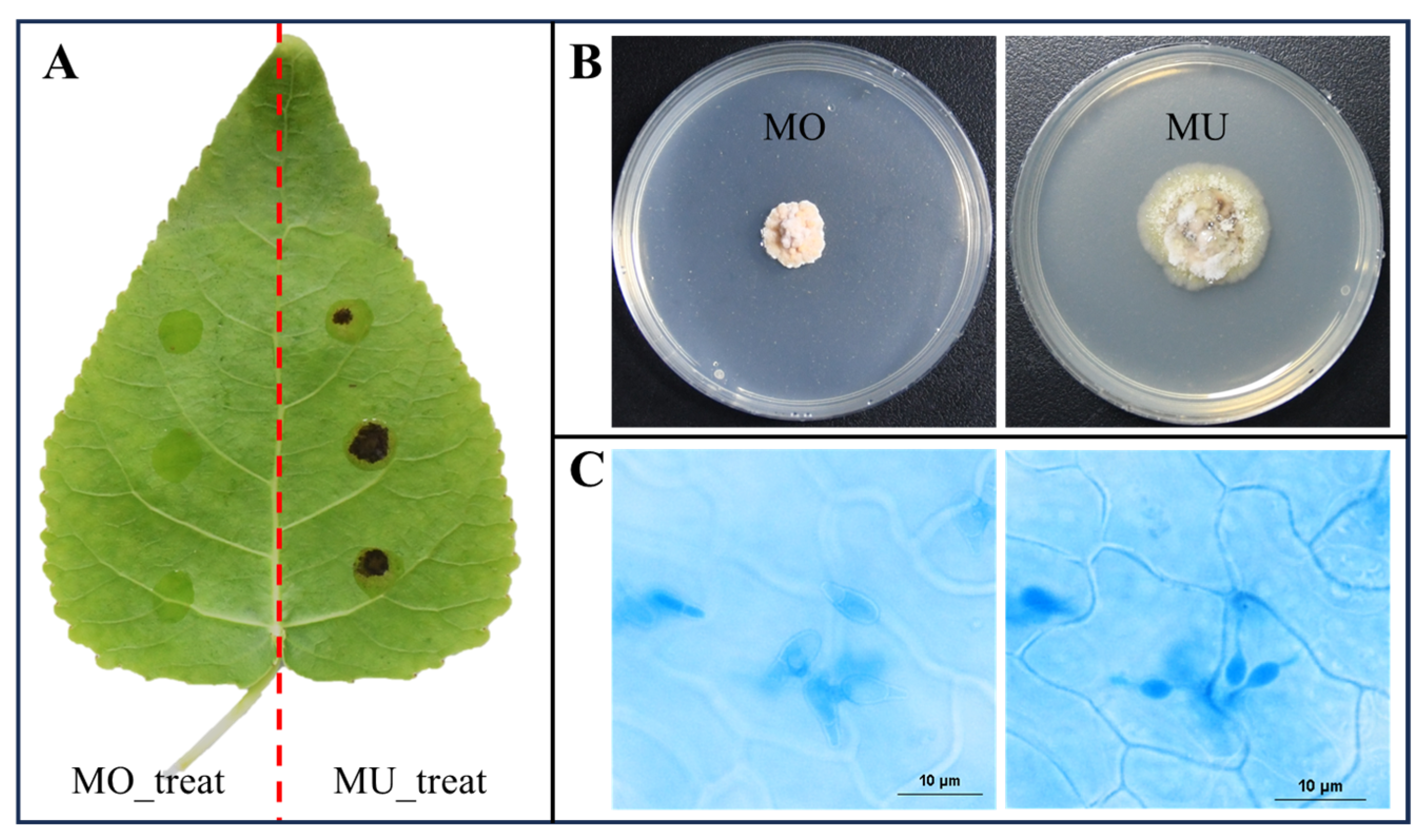
2.2. Inoculation, Microscopy, and Sample Collection
2.3. RNA Isolation and Sequencing
2.4. Read Mapping
2.5. DEG Analysis and Gene Annotation
2.6. Reverse-Transcription (qRT)-PCR
2.7. Nitroblue Tetrazolium (NBT) and 3,3-Diaminobenzidine (DAB) Staining
3. Results
3.1. Construction of the Interaction System and Formation of Penetration Structure
3.2. Summary of RNA Sequencing
3.3. Analysis of Differentially Expressed Genes (DEGs)
3.4. GO and KEGG Enrichment Analysis
3.5. Analysis of Biotic Stress Response Differentially Expressed Genes
3.6. Oxidative Burst
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keen, N.T. Gene-for-gene Complementarity in Plant-Pathogen Interactions. Annu. Rev. Genet. 1990, 24, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Orozco-Mosqueda, M.d.C.; Govindappa, M. Mechanisms of Biocontrol and Plant Growth-Promoting Activity in Soil Bacterial Species of Bacillus and Pseudomonas: A Review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Bushnell, W.R.; Roelfs, A.P. The Cereal Rusts; Harcourt Brace Jovanovich: Orlando, FL, USA, 1984; pp. 115–130. [Google Scholar] [CrossRef]
- Eriksson, J. Ueber die Specialisirung des Parasitismus bei den Getreiderostpilzen. Ber. Deutsch. Bot. Ges. 1894, 12, 292–331. [Google Scholar]
- Edel-Hermann, V.; Lecomte, C. Current Status of Fusarium oxysporum Formae Speciales and Races. Phytopathology 2018, 109, 512–530. [Google Scholar] [CrossRef]
- Coleman, J.J. The Fusarium solani Species Complex: Ubiquitous Pathogens of Agricultural Importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen Profile Update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- van Dam, P.; Fokkens, L.; Schmidt, S.M.; Linmans, J.H.J.; Kistler, H.C.; Ma, L.-J.; Rep, M. Effector Profiles Distinguish Formae Speciales of Fusarium oxysporum. Environ. Microbiol. 2016, 18, 4087–4102. [Google Scholar] [CrossRef]
- Lairini, K.; Perez-Espinosa, A.; Ruiz-Rubio, M. Tomatinase Induction in Formae Speciales of Fusarium oxysporumnon pathogenic of tomato plants. Physiol. Mol. Plant Pathol. 1997, 50, 37–52. [Google Scholar] [CrossRef]
- Kiran, K.; Rawal, H.C.; Dubey, H.; Jaswal, R.; Devanna, B.N.; Gupta, D.K.; Bhardwaj, S.C.; Prasad, P.; Pal, D.; Chhuneja, P.; et al. Draft Genome of the Wheat Rust Pathogen (Puccinia triticina) Unravels Genome-wide Structural Variations during Evolution. Genome Biol. Evol. 2016, 8, 2702–2721. [Google Scholar] [CrossRef]
- Nirmaladevi, D.; Venkataramana, M.; Srivastava, R.K.; Uppalapati, S.R.; Gupta, V.K.; Yli-Mattila, T.; Clement Tsui, K.M.; Srinivas, C.; Niranjana, S.R.; Chandra, N.S. Molecular Phylogeny, Pathogenicity and Toxigenicity of Fusarium oxysporum f. sp. lycopersici. Sci. Rep. 2016, 6, 21367. [Google Scholar] [CrossRef]
- Li, C. Two Specialized Forms of Marssonina populi (Lib.) Magn. J. Nanjing For. Univ. 1984, 27, 10–17. [Google Scholar] [CrossRef]
- Han, Z.; Li, C.; Huang, M. Comparative Studies of Isolates of Marssonina brunnea in China. Sci. Silvae Sin. 1998, 34, 59–65. [Google Scholar] [CrossRef]
- Zhang, Y.; He, W.; Yan, D.-H. Histopathologic Characterization of the Process of Marssonina brunnea Infection in Poplar Leaves. Can. J. For. Res. 2018, 48, 1302–1310. [Google Scholar] [CrossRef]
- Zhu, S.; Cao, Y.Z.; Jiang, C.; Tan, B.Y.; Wang, Z.; Feng, S.; Zhang, L.; Su, X.H.; Brejova, B.; Vinar, T.; et al. Sequencing the Genome of Marssonina brunnea Reveals Fungus-Poplar Co-evolution. BMC Genom. 2012, 13, 382. [Google Scholar] [CrossRef]
- Jiang, C.; He, B.; Huang, R.; Huang, M.; Xu, L. Identification and Functional Analysis of LysM Effectors from Marssonina brunnea. Australas. Plant Pathol. 2014, 43, 615–622. [Google Scholar] [CrossRef]
- Cheng, Q.; Cao, Y.; Jiang, C.; Xu, L.a.; Wang, M.; Zhang, S.; Huang, M. Identifying Secreted Proteins of Marssonina brunnea by Degenerate PCR. Proteomics 2010, 10, 2406–2417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, L.; Yan, D.-H.; He, W. Genome-Wide Transcriptome Analysis Reveals the Comprehensive Response of Two Susceptible Poplar Sections to Marssonina brunnea Infection. Genes 2018, 9, 154. [Google Scholar] [CrossRef]
- Chen, C.; Yao, Y.; Zhang, L.; Xu, M.; Jiang, J.; Dou, T.; Lin, W.; Zhao, G.; Huang, M.; Zhou, Y. A Comprehensive Analysis of the Transcriptomes of Marssonina brunnea and Infected Poplar Leaves to Capture Vital Events in Host-Pathogen Interactions. PLoS ONE 2015, 10, e0134246. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, Q.; Cui, R.; Xu, X.; Zhu, F.; Cheng, Q.; Li, X. High-Throughput Sequencing Reveals the Regulatory Networks of Transcriptome and Small RNAs during the Defense Against Marssonina brunnea in Poplar. Front. Plant Sci. 2021, 12, 719549. [Google Scholar] [CrossRef]
- Li, L.; Sun, W.; Zhou, P.; Wei, H.; Wang, P.; Li, H.; Rehman, S.; Li, D.; Zhuge, Q. Genome-Wide Characterization of Dirigent Proteins in Populus: Gene Expression Variation and Expression Pattern in Response to Marssonina brunnea and Phytohormones. Forests 2021, 12, 507. [Google Scholar] [CrossRef]
- Erickson, J.E.; Stanosz, G.R.; Kruger, E.L. Photosynthetic Consequences of Marssonina Leaf Spot Differ between two Poplar Hybrids. New Phytol. 2004, 161, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, Y.; Zhao, Y.; Xu, W.; Li, Y.; Xie, J.; Zhang, D. Key Genes and Genetic Interactions of Plant-Pathogen Functional Modules in Poplar Infected by Marssonina brunnea. Mol. Plant-Microbe Interact. 2020, 33, 1080–1090. [Google Scholar] [CrossRef]
- Liao, Y.; Cui, R.; Xu, X.; Cheng, Q.; Li, X. Jasmonic Acid- and Ethylene-Induced Mitochondrial Alternative Oxidase Stimulates Marssonina brunnea Defense in Poplar. Plant Cell Physiol. 2020, 61, 2031–2042. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, Y.; Movahedi, A.; Wei, H.; Zhuge, Q. Thaumatin-like Protein(Pe-TLP) Acts as a Positive Factor in Transgenic Poplars Enhanced Resistance to Spots Disease. Physiol. Mol. Plant Pathol. 2020, 112, 101512. [Google Scholar] [CrossRef]
- Dixon, R.A.; Liu, C.; Jun, J.H. Metabolic Engineering of Anthocyanins and Condensed Tannins in Plants. Curr. Opin. Biotechnol. 2013, 24, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ran, L.; Hou, Y.; Tian, Q.; Li, C.; Liu, R.; Fan, D.; Luo, K. The Transcription Factor MYB115 Contributes to the Regulation of Proanthocyanidin Biosynthesis and Enhances Fungal Resistance in Poplar. New Phytol. 2017, 215, 351–367. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A Refined Protein Classification and Annotation Framework Applicable to Multi-Omics Data Analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef]
- de Wit, P.J.G.M. Molecular Characterization of Gene-for-Gene Systems in Plant-Fungus Interactions and the Application of Avirulence Genes in Control of Plant Pathogens. Annu. Rev. Phytopathol. 1992, 30, 391–418. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.P.; Moscou, M.J.; Bogdanove, A.J.; Whitham, S.A. Transcript Profiling in Host-Pathogen Interactions. Annu. Rev. Phytopathol. 2007, 45, 329–369. [Google Scholar] [CrossRef]
- Liu, J.; Yu, Z.; Yang, Y.; Ma, Y.; Yu, X.; Wei, W.; Song, C.; Guo, X.; Yang, X. Gene Profiling of Compatible and Incompatible Interactions between Potato and Phytophthora infestans. Am. J. Potato Res. 2020, 97, 376–392. [Google Scholar] [CrossRef]
- Coram, T.E.; Wang, M.; Chen, X. Transcriptome Analysis of the Wheat-Puccinia striiformis f. sp. tritici Interaction. Mol. Plant Pathol. 2008, 9, 157–169. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant-pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Ravi, B.; Foyer, C.H.; Pandey, G.K. The Integration of Reactive Oxygen Species (ROS) and Calcium Signalling in Abiotic Stress Responses. Plant Cell Environ. 2023, 46, 1985–2006. [Google Scholar] [CrossRef]
- Liu, Y.; He, C. Regulation of Plant Reactive Oxygen Species (ROS) in Stress Responses: Learning from AtRBOHD. Plant Cell Rep. 2016, 35, 995–1007. [Google Scholar] [CrossRef]
- Greenberg, J.T.; Yao, N. The Role and Regulation of Programmed Cell Death in plant-pathogen interactions. Cell. Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef]
- Shetty, N.P.; Jorgensen, H.J.L.; Jensen, J.D.; Collinge, D.B.; Shetty, H.S. Roles of Reactive Oxygen Species in Interactions between Plants and Pathogens. Eur. J. Plant Pathol. 2008, 121, 267–280. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, C.; Cheng, Y.; Chen, X.; Han, Q.; Huang, L.; Wei, G.; Kang, Z. Histological and Cytological Characterization of Adult Plant Resistance to Wheat Stripe Rust. Plant Cell Rep. 2012, 31, 2121–2137. [Google Scholar] [CrossRef]
- Barna, B.; Fodor, J.; Harrach, B.D.; Pogány, M.; Király, Z. The Janus Face of Reactive Oxygen Species in Resistance and Susceptibility of Plants to Necrotrophic and Biotrophic Pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive Oxygen Species as Signals that Modulate Plant Stress Responses and Programmed Cell Death. Bioessays 2006, 28, 1091–1101. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SampleID | Clean Reads | Q20 | Q30 | Input Reads | Mapped Reads | Mapped Rate |
|---|---|---|---|---|---|---|
| control_8hpi_1 | 41,519,136 | 97.83% | 93.10% | 41,519,136 | 35,888,940 | 86.44% |
| control_8hpi_2 | 38,736,712 | 97.86% | 93.15% | 38,736,712 | 33,457,893 | 86.37% |
| control_8hpi_3 | 37,580,346 | 97.75% | 92.78% | 37,580,346 | 32,366,453 | 86.13% |
| MO_8hpi_1 | 43,733,376 | 97.55% | 92.37% | 43,733,376 | 37,744,736 | 86.31% |
| MO_8hpi_2 | 44,294,756 | 97.58% | 92.47% | 44,294,756 | 38,352,233 | 86.58% |
| MO_8hpi_3 | 38,157,696 | 98.12% | 93.76% | 38,157,696 | 33,351,061 | 87.40% |
| MU_8hpi_1 | 43,474,240 | 97.68% | 92.84% | 43,474,240 | 33,134,118 | 76.22% |
| MU_8hpi_2 | 47,154,366 | 97.60% | 92.63% | 47,154,366 | 36,744,352 | 77.92% |
| MU_8hpi_3 | 42,583,954 | 97.68% | 92.82% | 42,583,954 | 36,758,674 | 86.32% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Tian, L. Transcriptome Analysis and Reactive Oxygen Species Detection Suggest Contrasting Molecular Mechanisms in Populus canadensis’ Response to Different Formae Speciales of Marssonina brunnea. Genes 2024, 15, 116. https://doi.org/10.3390/genes15010116
Zhang Y, Tian L. Transcriptome Analysis and Reactive Oxygen Species Detection Suggest Contrasting Molecular Mechanisms in Populus canadensis’ Response to Different Formae Speciales of Marssonina brunnea. Genes. 2024; 15(1):116. https://doi.org/10.3390/genes15010116
Chicago/Turabian StyleZhang, Yanfeng, and Longyan Tian. 2024. "Transcriptome Analysis and Reactive Oxygen Species Detection Suggest Contrasting Molecular Mechanisms in Populus canadensis’ Response to Different Formae Speciales of Marssonina brunnea" Genes 15, no. 1: 116. https://doi.org/10.3390/genes15010116
APA StyleZhang, Y., & Tian, L. (2024). Transcriptome Analysis and Reactive Oxygen Species Detection Suggest Contrasting Molecular Mechanisms in Populus canadensis’ Response to Different Formae Speciales of Marssonina brunnea. Genes, 15(1), 116. https://doi.org/10.3390/genes15010116