
Association of FGF4L1 Retrogene Insertion with Prolapsed Gland of the Nictitans (Cherry Eye) in Dogs

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Genotyping
2.3. Clinical Information
2.4. Inclusion Criteria for Genetic Analysis
2.5. Genotype Analysis
2.6. Additional Statistical Analyses
2.7. Ethics Statements
3. Results
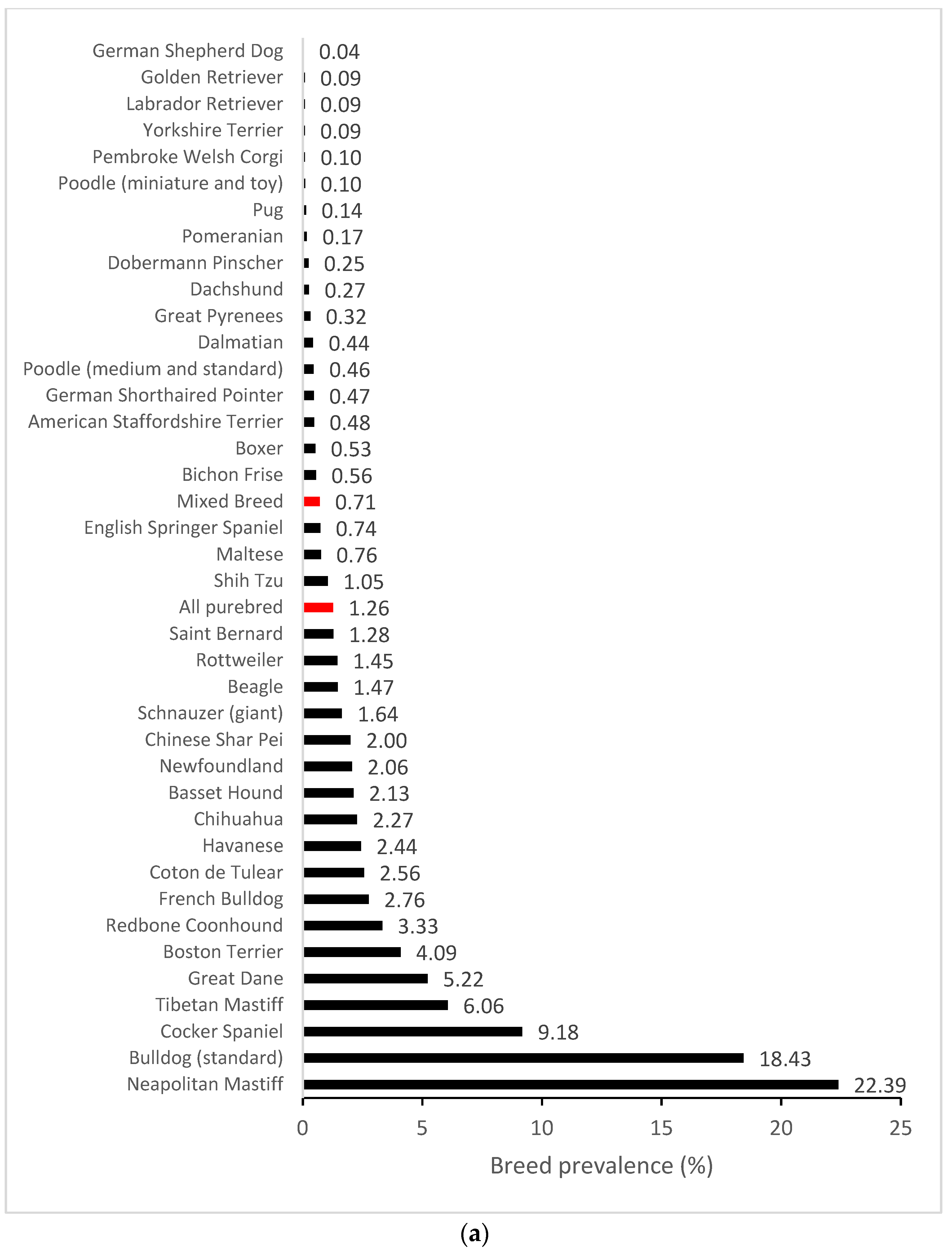
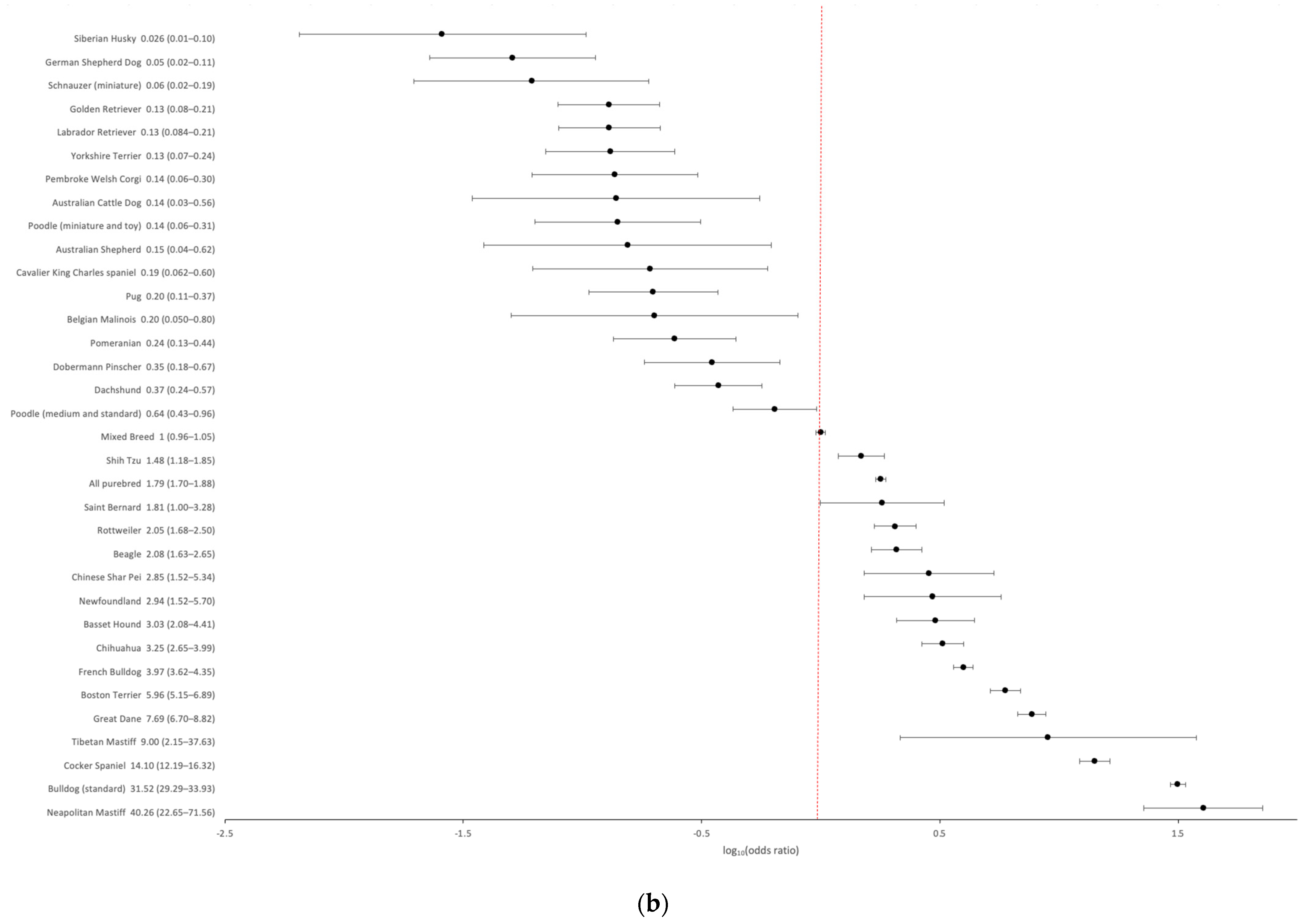
3.1. Epidemiology
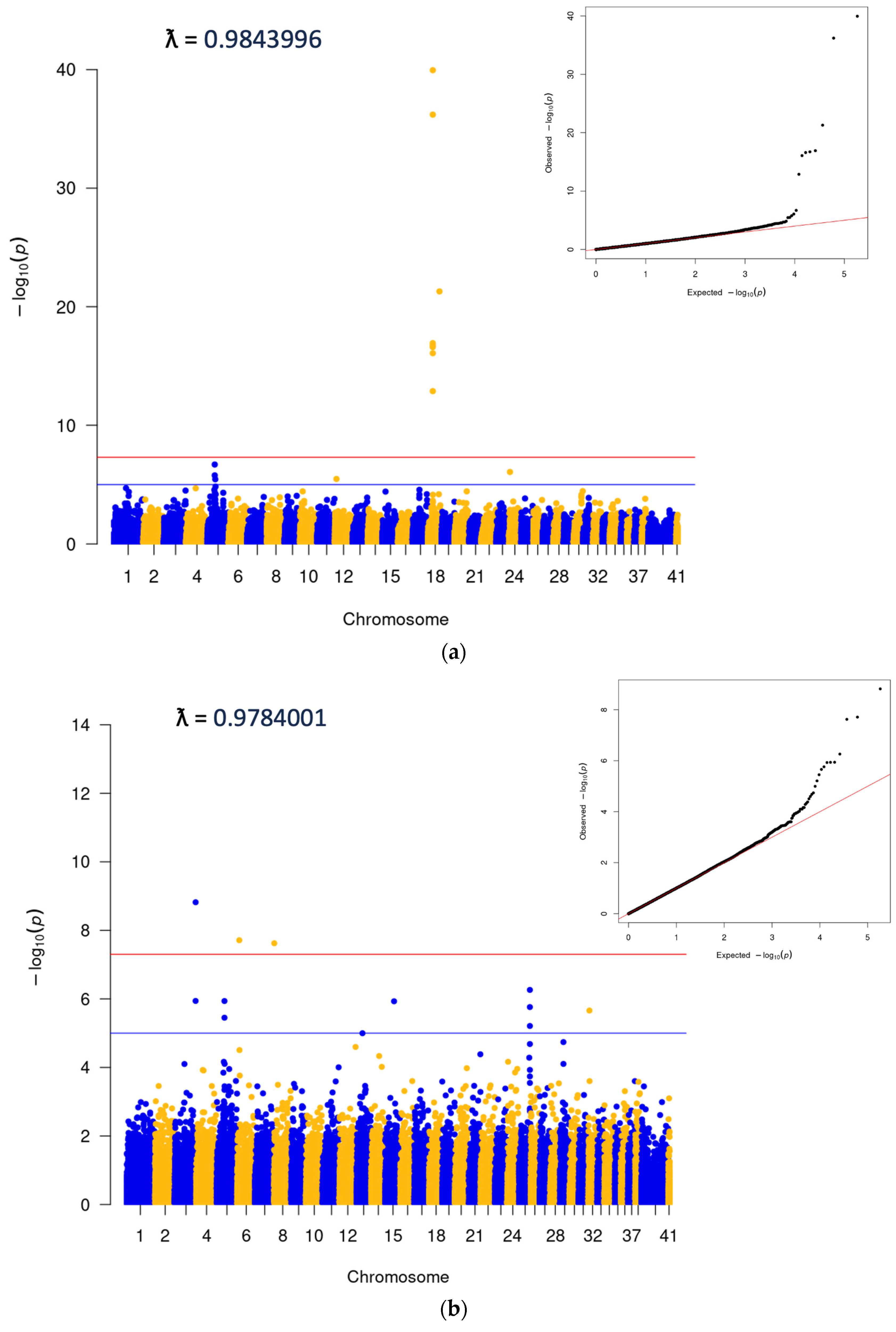
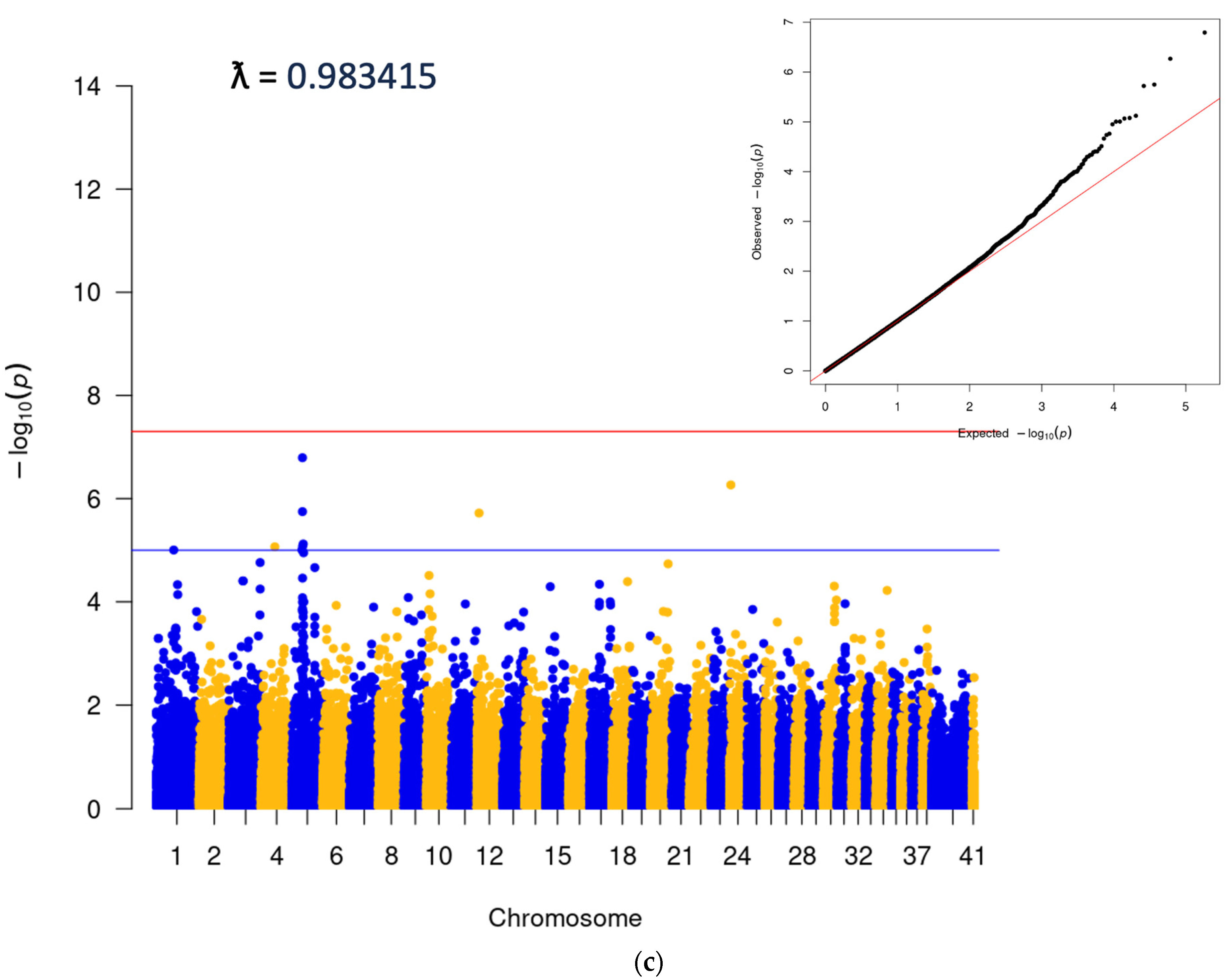
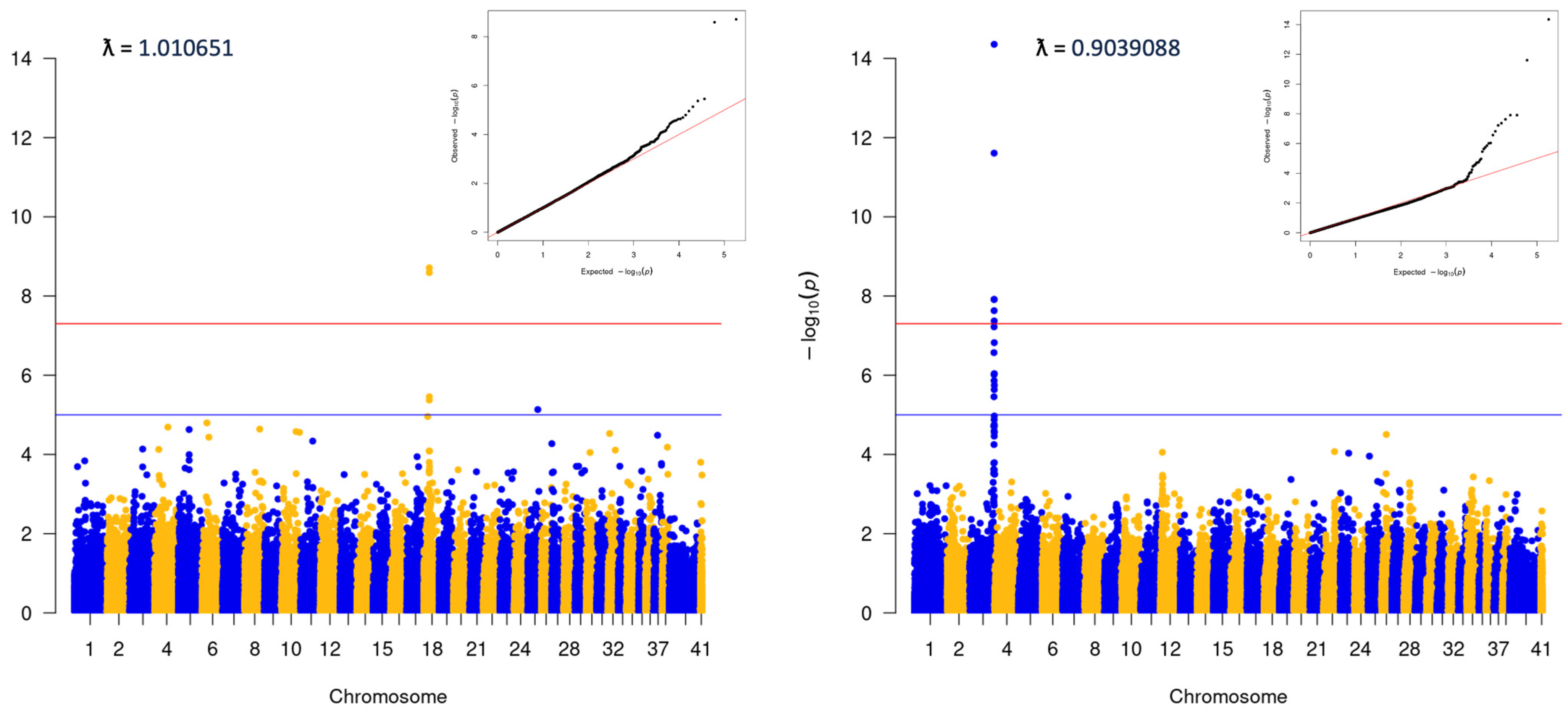
3.2. Genome-Wide Association Analysis
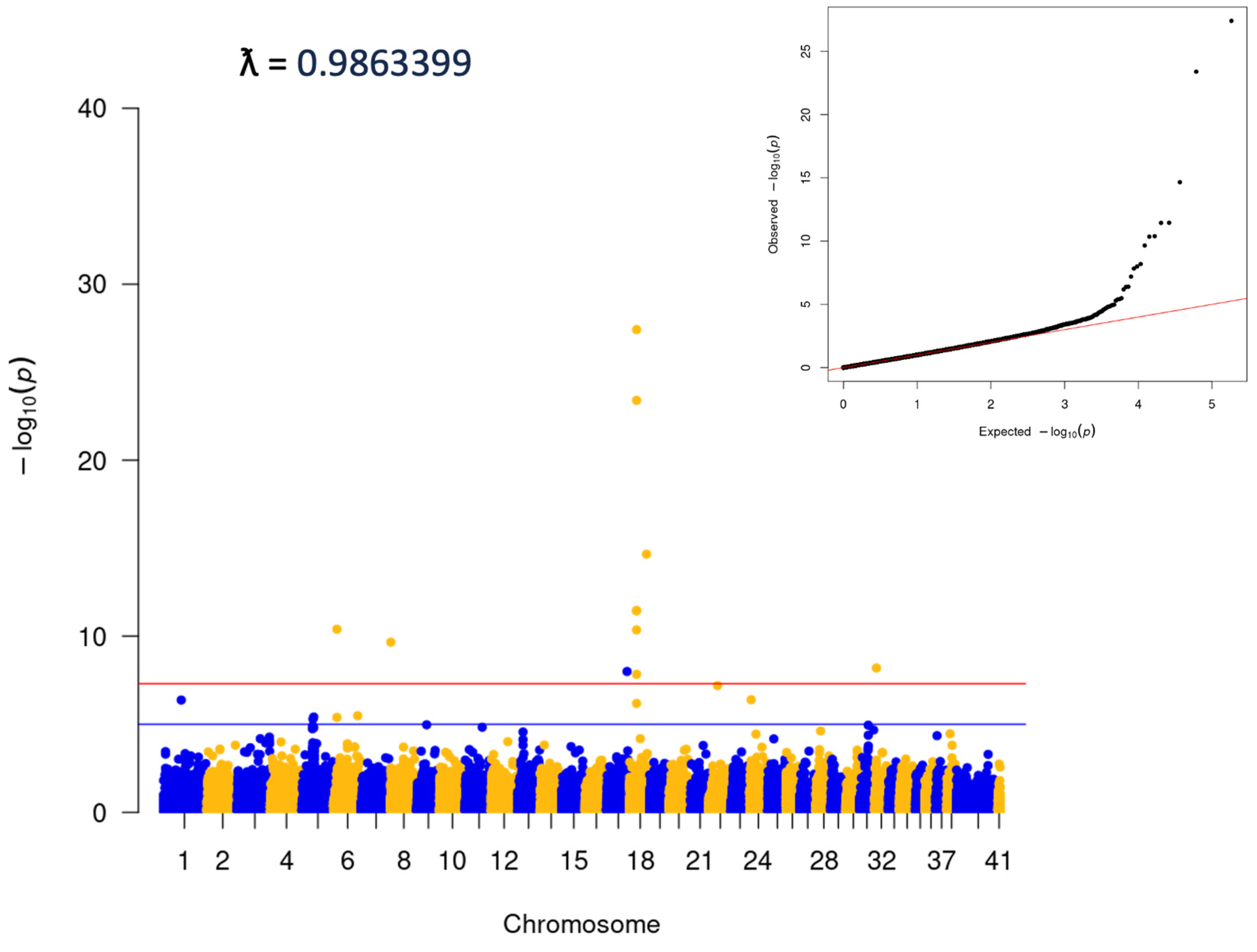
3.3. Association between Cherry Eye and the FGF4L1 Retrogene Variant in Mixed Breed Dogs
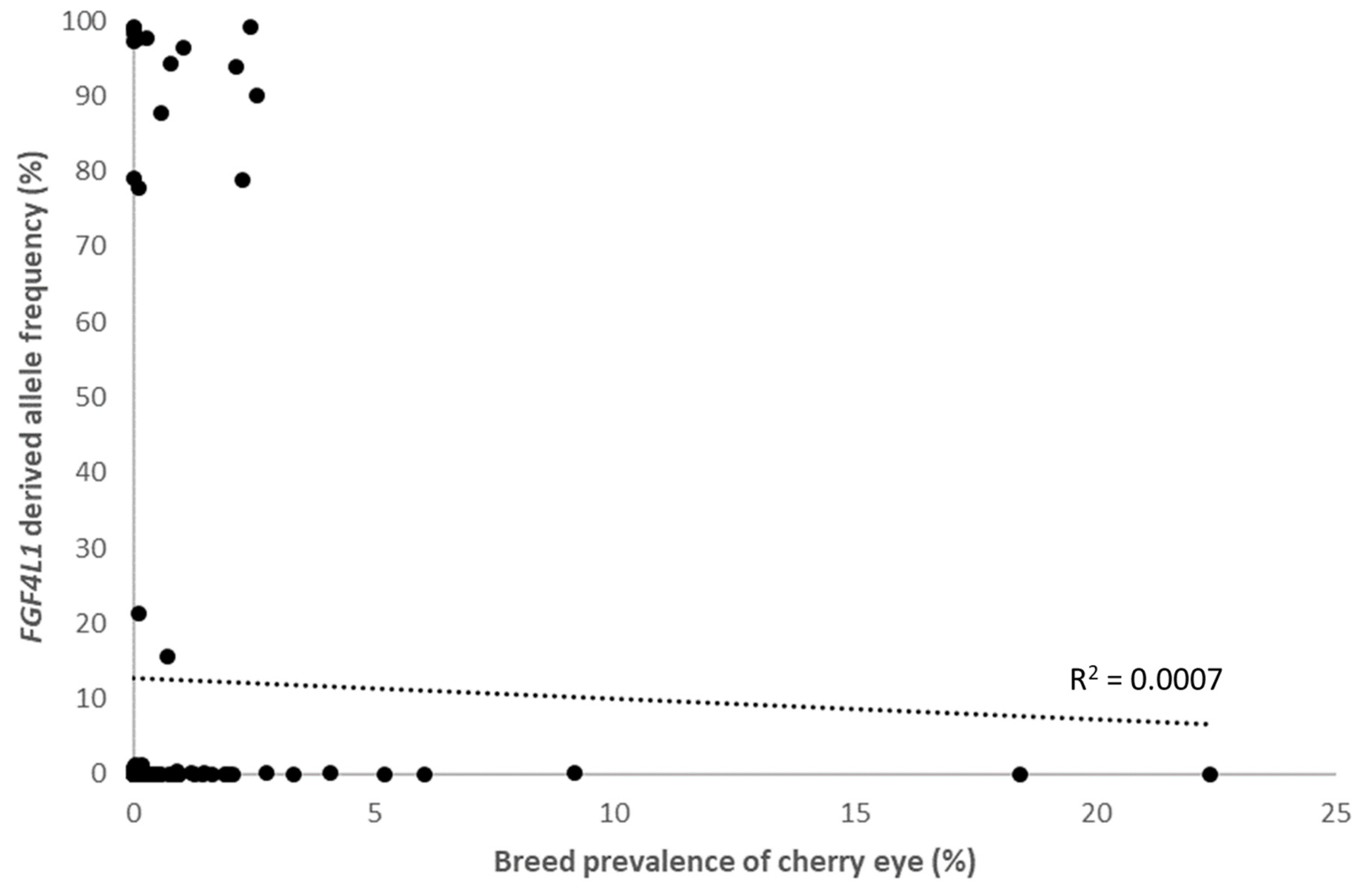
3.4. Within Breed FGF4L1 Derived Allele Frequency and Clinical Prevalence of Cherry Eye
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dugan, S.J.; Severin, G.A.; Hungerford, L.L.; Whiteley, H.E.; Roberts, S.M. Clinical and Histologic Evaluation of the Prolapsed Third Eyelid Gland in Dogs. J. Am. Vet. Med. Assoc. 1992, 201, 1861–1867. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, T.; Brehm, H.; Amselgruber, W.M. The Cartilage of the Third Eyelid: A Comparative Macroscopical and Histological Study in Domestic Animals. Ann. Anat.-Anat. Anz. 2001, 183, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Sandmeyer, L.S.; Levitt, S.; Leis, M.L. Diagnostic Ophthalmology. Can. Vet. J. 2022, 63, 979–980. [Google Scholar] [PubMed]
- O’Neill, D.G.; Yin, Y.; Tetas Pont, R.; Brodbelt, D.C.; Church, D.B.; Pegram, C.; Mustikka, M. Breed and Conformational Predispositions for Prolapsed Nictitating Membrane Gland (PNMG) in Dogs in the UK: A VetCompass Study. PLoS ONE 2022, 17, e0260538. [Google Scholar] [CrossRef]
- Gelatt, K.N.; Ben-Shlomo, G.; Gilger, B.C.; Hendrix, D.V.H.; Kern, T.J.; Plummer, C.E. Veterinary Ophthalmology, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; Volume 1. [Google Scholar]
- Edelmann, M.L.; Miyadera, K.; Iwabe, S.; Komáromy, A.M. Investigating the Inheritance of Prolapsed Nictitating Membrane Glands in a Large Canine Pedigree. Vet. Ophthalmol. 2013, 16, 416–422. [Google Scholar] [CrossRef]
- Fossum, T.W.; Hedlund, C.S. Small Animal Surgery, 3rd ed.; Mosby/Elsevier: St. Louis, MO, USA, 2007. [Google Scholar]
- Mazzucchelli, S.; Vaillant, M.D.; Wéverberg, F.; Arnold-Tavernier, H.; Honegger, N.; Payen, G.; Vanore, M.; Liscoet, L.; Thomas, O.; Clerc, B.; et al. Retrospective Study of 155 Cases of Prolapse of the Nictitating Membrane Gland in Dogs. Vet. Rec. 2012, 170, 443. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.V.; Duddy, J.M.; McClurg, K. Prolapse of the Gland of the Third Eyelid in Dogs: A Retrospective Study of 89 Cases (1980 to 1990). J. Am. Anim. Hosp. Assoc. 1993, 29, 56–60. [Google Scholar]
- White, C.; Brennan, M. An Evidence-Based Rapid Review of Surgical Techniques for Correction of Prolapsed Nictitans Glands in Dogs. Vet. Sci. 2018, 5, 75. [Google Scholar] [CrossRef]
- Donner, J.; Kaukonen, M.; Anderson, H.; Möller, F.; Kyöstilä, K.; Sankari, S.; Hytönen, M.; Giger, U.; Lohi, H. Genetic Panel Screening of Nearly 100 Mutations Reveals New Insights into the Breed Distribution of Risk Variants for Canine Hereditary Disorders. PLoS ONE 2016, 11, e0161005. [Google Scholar] [CrossRef]
- Garrigan, D.; Huff, J.; Chodroff Foran, R. BCSYS: An Accurate and Scalable Local Ancestry Classifier. Available online: https://www.wisdompanel.com/downloads/wp-breed-detection.pdf (accessed on 1 December 2023).
- Turner, S. qqman: An R Package for Visualizing GWAS Results Using Q-Q and Manhattan Plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Zhou, X.; Stephens, M. Genome-Wide Efficient Mixed-Model Analysis for Association Studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Shaun Purcell PLINK 1.9. Available online: http://pngu.mgh.harvard.edu/purcell/plink/ (accessed on 1 December 2023).
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An Expressed Fgf4 Retrogene Is Associated with Breed-Defining Chondrodysplasia in Domestic Dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Plassais, J.; Kim, J.; Davis, B.W.; Karyadi, D.M.; Hogan, A.N.; Harris, A.C.; Decker, B.; Parker, H.G.; Ostrander, E.A. Whole Genome Sequencing of Canids Reveals Genomic Regions under Selection and Variants Influencing Morphology. Nat. Commun. 2019, 10, 1489. [Google Scholar] [CrossRef]
- Farooq, M.; Khan, A.W.; Kim, M.S.; Choi, S. The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells 2021, 10, 3242. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.-R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.-H.; Shin, U.S.; Kim, H.-W. Fibroblast Growth Factors: Biology, Function, and Application for Tissue Regeneration. J. Tissue Eng. 2010, 1, 218142. [Google Scholar] [CrossRef]
- Martínez, S.; Fajardo, R.; Valdés, J.; Ulloa-Arvizu, R.; Alonso, R. Histopathologic Study of Long-Bone Growth Plates Confirms the Basset Hound as an Osteochondrodysplastic Breed. Can. J. Vet. Res. 2007, 71, 66–69. [Google Scholar]
- Rasmussen, P.G.; Reimann, I. Dysostosis Enchondralis of the Ulnar Bone in the Basset Hound. Acta Vet. Scand. 1977, 18, 31–39. [Google Scholar] [CrossRef]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 Retrogene on CFA12 Is Responsible for Chondrodystrophy and Intervertebral Disc Disease in Dogs. Proc. Natl. Acad. Sci. USA 2017, 114, 11476–11481. [Google Scholar] [CrossRef]
- Shiang, R.; Thompson, L.M.; Zhu, Y.-Z.; Church, D.M.; Fielder, T.J.; Bocian, M.; Winokur, S.T.; Wasmuth, J.J. Mutations in the Transmembrane Domain of FGFR3 Cause the Most Common Genetic Form of Dwarfism, Achondroplasia. Cell 1994, 78, 335–342. [Google Scholar] [CrossRef]
- Grillo, L.; Greco, D.; Pettinato, R.; Avola, E.; Potenza, N.; Castiglia, L.; Spalletta, A.; Amata, S.; Di Benedetto, D.; Luciano, D.; et al. Increased FGF3 and FGF4 Gene Dosage Is a Risk Factor for Craniosynostosis. Gene 2014, 534, 435–439. [Google Scholar] [CrossRef]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR Signaling in Health and Disease. Signal Transduct. Target. Ther. 2020, 5, 181. [Google Scholar] [CrossRef]
- Minowada, G.; Jarvis, L.A.; Chi, C.L.; Neubüser, A.; Sun, X.; Hacohen, N.; Krasnow, M.A.; Martin, G.R. Vertebrate Sprouty Genes Are Induced by FGF Signaling and Can Cause Chondrodysplasia When Overexpressed. Development 1999, 126, 4465–4475. [Google Scholar] [CrossRef]
- Guo, C.; Degnin, C.R.; Laederich, M.B.; Lunstrum, G.P.; Holden, P.; Bihlmaier, J.; Krakow, D.; Cho, Y.-J.; Horton, W.A. Sprouty 2 Disturbs FGFR3 Degradation in Thanatophoric Dysplasia Type II: A Severe Form of Human Achondroplasia. Cell Signal. 2008, 20, 1471–1477. [Google Scholar] [CrossRef]
- Plikus, M.V.; Wang, X.; Sinha, S.; Forte, E.; Thompson, S.M.; Herzog, E.L.; Driskell, R.R.; Rosenthal, N.; Biernaskie, J.; Horsley, V. Fibroblasts: Origins, Definitions, and Functions in Health and Disease. Cell 2021, 184, 3852–3872. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Case Population Characteristics | Odds Ratio (95% CI) | p-Value |
|---|---|---|---|
| Sex | |||
| Female | 2942 (43.57%) | 1 | |
| Male | 3811 (56.43%) | 1.10 (1.05–1.15) | <0.001 |
| Neuter | |||
| Intact | 2773 (41.06%) | 1 | |
| Neutered/spayed | 3980 (58.94%) | 0.79 (0.76–0.83) | <10−16 |
| Breed | |||
| Mixed Breed | 3826 (56.66%) | 1 | |
| Purebred | 2927 (43.34%) | 1.79 (1.70–1.88) | <0.0001 |
| Skull shape | |||
| Mesocephalic | 522 (7.73%) | 1 | |
| Brachycephalic | 2072 (30.68%) | 7.60 (6.90–8.36) | <10−16 |
| Dolichocephalic | 332 (4.92%) | 1.52 (1.32–1.74) | <10−8 |
| Uncategorized (mixed breed) | 3827 (56.67%) | 1.61 (1.47–1.77) | <10−16 |
| Age at first diagnosis | |||
| Average | 0.83 years | ||
| Median | 0.48 years | ||
| Range | 0.07–15.36 years | ||
| 0–1 years | 4743 (79.31%) | 28.63 (23.13–35.44) | <10−16 |
| 1–2 years | 774 (12.94%) | 4.87 (3.90–6.08) | <10−16 |
| 2–3 years | 252 (4.21%) | 1.97 (1.54–2.52) | <10−7 |
| 3–4 years | 86 (1.44%) | 1 | |
| 4–5 years | 39 (0.65%) | 2.33 (1.60–3.41) | <0.0001 |
| >5 years | 86 (1.44%) | 4.34 (3.22–5.85) | <10−16 |
| Group | 0 | 1 | Odds Ratio (95% CI) | 2 | Odds Ratio (95% CI) | Total | Chi Square p-Value |
|---|---|---|---|---|---|---|---|
| All | |||||||
| Cases Controls | 4445 (66.06%) 565,269 (74.27%) | 753 (11.19%) 83,819 (11.01%) | 1.14 (1.06–1.23) | 1531 (22.75%) 112,045 (14.72%) | 1.74 (1.64–1.84) | 6729 761,133 | <10−16 |
| Purebred | |||||||
| Cases Controls | 2659 (91.00%) 184,865 (81.09%) | 39 (1.33%) 8209 (3.60%) | 0.33 (0.24–0.45) | 224 (7.67%) 34,915 (15.31%) | 0.45 (0.39–0.51) | 2922 227,989 | <10−16 |
| Mixed Breed | |||||||
| Cases Controls | 1786 (46.91%) 380,404 (71.35%) | 714 (18.75%) 75,610 (14.18%) | 2.01 (1.84–2.19) | 1307 (34.33%) 77,130 (14.47%) | 3.61 (3.36–3.88) | 3807 533,144 | <10−16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freyer, J.; Labadie, J.D.; Huff, J.T.; Denyer, M.; Forman, O.P.; Chodroff Foran, R.; Donner, J. Association of FGF4L1 Retrogene Insertion with Prolapsed Gland of the Nictitans (Cherry Eye) in Dogs. Genes 2024, 15, 198. https://doi.org/10.3390/genes15020198
Freyer J, Labadie JD, Huff JT, Denyer M, Forman OP, Chodroff Foran R, Donner J. Association of FGF4L1 Retrogene Insertion with Prolapsed Gland of the Nictitans (Cherry Eye) in Dogs. Genes. 2024; 15(2):198. https://doi.org/10.3390/genes15020198
Chicago/Turabian StyleFreyer, Jamie, Julia D. Labadie, Jason T. Huff, Michael Denyer, Oliver P. Forman, Rebecca Chodroff Foran, and Jonas Donner. 2024. "Association of FGF4L1 Retrogene Insertion with Prolapsed Gland of the Nictitans (Cherry Eye) in Dogs" Genes 15, no. 2: 198. https://doi.org/10.3390/genes15020198
APA StyleFreyer, J., Labadie, J. D., Huff, J. T., Denyer, M., Forman, O. P., Chodroff Foran, R., & Donner, J. (2024). Association of FGF4L1 Retrogene Insertion with Prolapsed Gland of the Nictitans (Cherry Eye) in Dogs. Genes, 15(2), 198. https://doi.org/10.3390/genes15020198