
Differences in Immune Characteristics and Related Gene Expression in Spleen among Ningxiang, Berkshire Breeds and Their Hybrid Pigs
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Measurement of Physiological and Biochemical Indicators
2.2.1. Determination of Immunological Indicators in Plasma and Spleen by ELISA
2.2.2. Measurement of Plasma Biochemical Indicators
2.3. Preparation of Samples, Construction of RNA Libraries, and Sequencing
2.4. Differential Expression Analysis and Functional Enrichment
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Statistical Analysis
3. Results
3.1. Plasma and Spleen Physiological and Biochemical Indices
3.2. Annotation of DEGs and Their Functions in Different Pig Breeds
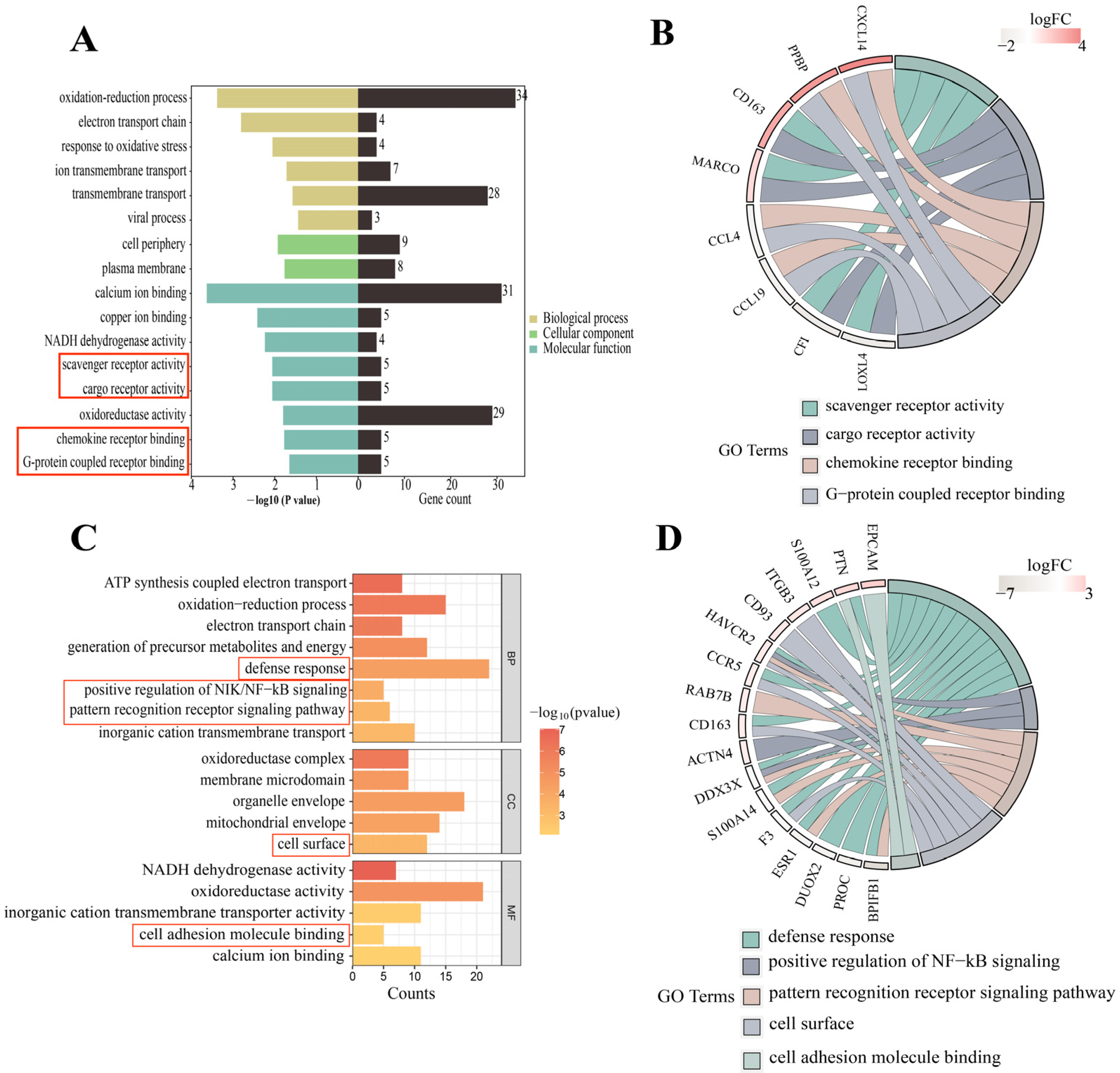
3.3. GO Enrichment Analysis
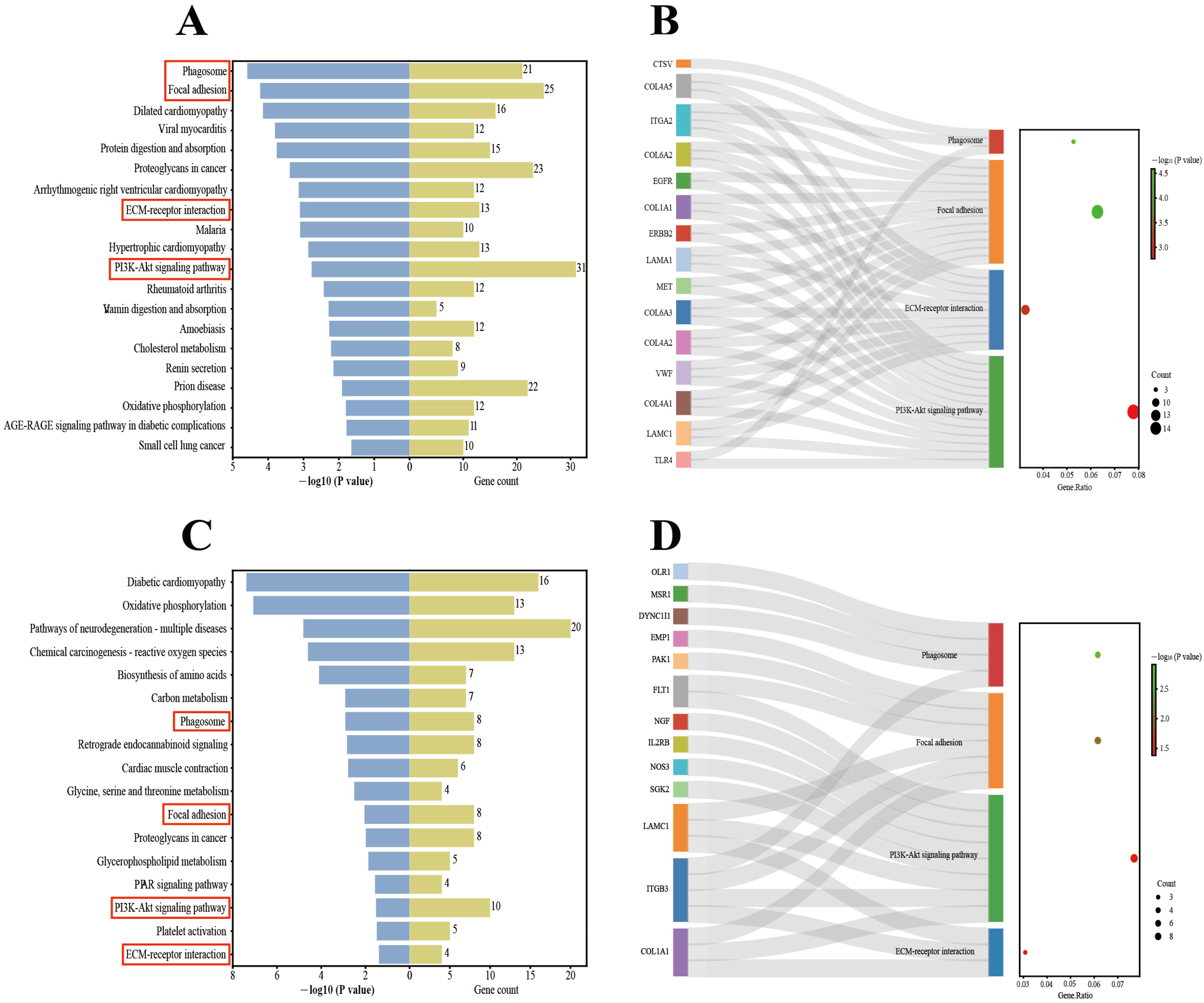
3.4. KEGG Enrichment Analysis
3.5. PPI Network and Hub Gene Identification
3.6. Validation by qRT-PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.Z.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Cen, W.M.; Xing, S.H.; Chen, J.N.; Wen, A.X.; He, T.; et al. Carcass and meat quality traits of four commercial pig crossbreeds in China. Genet. Mol. Res. 2012, 11, 4447–4455. [Google Scholar] [CrossRef]
- Li, B.; Yang, J.; He, J.; Peng, X.; Zeng, Q.; Song, Y.; Xu, K.; Ma, H. Characterization of the whole transcriptome of spleens from Chinese indigenous breed Ningxiang pig reveals diverse coding and non-coding RNAs for immunity regulation. Genomics 2021, 113, 2468–2482. [Google Scholar] [CrossRef] [PubMed]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Pittet, M.J. The Spleen in Local and Systemic Regulation of Immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef]
- Morawietz, G.; Ruehl-Fehlert, C.; Kittel, B.; Bube, A.; Keane, K.; Halm, S.; Heuser, A.; Hellmann, J. Revised guides for organ sampling and trimming in rats and mice—Part 3. A joint publication of the RITA and NACAD groups. Exp. Toxicol. Pathol. 2004, 55, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Clapperton, M.; Bishop, S.C.; Glass, E.J. Innate immune traits differ between Meishan and Large White pigs. Vet. Immunol. Immunopathol. 2005, 104, 131–144. [Google Scholar] [CrossRef]
- Liang, W.; Li, Z.; Wang, P.; Fan, P.; Zhang, Y.; Zhang, Q.; Wang, Y.; Xu, X.; Liu, B. Differences of immune responses between Tongcheng (Chinese local breed) and Large White pigs after artificial infection with highly pathogenic porcine reproductive and respiratory syndrome virus. Virus Res. 2016, 215, 84–93. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Wang, H.; Wang, H.; Liu, J.-F.; Wu, Y.; Guo, J. Transcriptomic Analysis Identifies Candidate Genes and Gene Sets Controlling the Response of Porcine Peripheral Blood Mononuclear Cells to Poly I:C Stimulation. G3 Genes Genomes Genet. 2016, 6, 1267–1275. [Google Scholar] [CrossRef]
- Choi, I.-S.; Collisson, E.W.; Maheswaran, S.K.; Yoo, H.S. Evaluation of cytokine gene expression in porcine spleen cells, peripheral blood mononuclear cells, and alveolar macrophages by competitive RT-PCR. FEMS Immunol. Med. Microbiol. 2002, 34, 119–126. [Google Scholar] [CrossRef]
- Hou, G.; Peng, W.; Wei, L.; Li, R.; Huang, X.; Yin, Y. Probiotics and Achyranthes bidentata Polysaccharides Improve Growth Performance via Promoting Intestinal Nutrient Utilization and Enhancing Immune Function of Weaned Pigs. Animals 2021, 11, 2617. [Google Scholar] [CrossRef]
- Porter, P. Transfer of immunoglobulins IgG, IgA and IgM to lacteal secretions in the parturient sow and their absorption by the neonatal piglet. Biochim. Biophys. Acta 1969, 181, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Liu, Y.; Wang, G.; He, Y.; Hu, S.; Li, Y.; Shi, W.; Wu, J.; Wang, S.; Liu, H.T.; et al. Comparative Analysis of Immune Responses in Pigs to High and Low Pathogenic Porcine Reproductive and Respiratory Syndrome Viruses Isolated in China. Transbound. Emerg. Dis. 2015, 62, e1–e10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cooper, D.K.C.; Wang, H.; Chen, P.; He, C.; Cai, Z.; Mou, L.; Luan, S.; Gao, H.J.X. Potential pathological role of pro-inflammatory cytokines (IL-6, TNF-α, and IL-17) in xenotransplantation. Xenotransplantation 2019, 26, e12502. [Google Scholar] [CrossRef]
- Standiford, T.J.; Strieter, R.M.; Chensue, S.W.; Westwick, J.; Kasahara, K.; Kunkel, S.L. IL-4 inhibits the expression of IL-8 from stimulated human monocytes. J. Immunol. 1990, 145, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- te Velde, A.A.; Huijbens, R.J.; Heije, K.; de Vries, J.E.; Figdor, C.G. Interleukin-4 (IL-4) inhibits secretion of IL-1 beta, tumor necrosis factor alpha, and IL-6 by human monocytes. Blood 1990, 76, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Tebo, J.M.; Hamilton, T.A. IL-4 suppresses cytokine gene expression induced by IFN-gamma and/or IL-2 in murine peritoneal macrophages. J. Immunol. 1992, 148, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.H.; Jones, C.A.; Finlay-Jones, J.J. Interleukin-4 suppression of monocyte tumour necrosis factor-alpha production. Dependence on protein synthesis but not on cyclic AMP production. Immunology 1992, 76, 560–565. [Google Scholar] [PubMed]
- Kim, J.J.; Yang, J.S.; Manson, K.H.; Weiner, D.B. Modulation of antigen-specific cellular immune responses to DNA vaccination in rhesus macaques through the use of IL-2, IFN-gamma, or IL-4 gene adjuvants. Vaccine 2001, 19, 2496–2505. [Google Scholar] [CrossRef]
- Zhang, X.; Li, G.; Gao, L.; Mu, L.; Zhang, L.; Cong, Y.; Ding, Z. Positive inductive effect of IL-18 on virus-specific immune responses induced by PRRSV-GP5 DNA vaccine in swine. Res. Vet. Sci. 2013, 94, 346–353. [Google Scholar] [CrossRef]
- Toyoshima, S.; Wakamatsu, E.; Ishida, Y.; Obata, Y.; Kurashima, Y.; Kiyono, H.; Abe, R. The spleen is the site where mast cells are induced in the development of food allergy. Int. Immunol. 2017, 29, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Miossec, P.; Kolls, J.K. Targeting IL-17 and TH17 cells in chronic inflammation. Nat. Rev. Drug Discov. 2012, 11, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Qian, Y. IL-17 family cytokines mediated signaling in the pathogenesis of inflammatory diseases. Cell Signal. 2013, 25, 2335–2347. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, Q.D.; Li, K. Studies on Some lmmune Traits of Tongcheng, Swedish Landrace, BritishLarge White and Their Three-way Crossbred Pigs. Acta Vet. Zootech. Sin. 2005, 36, 735–738. (In Chinese) [Google Scholar]
- Schoenborn, J.R.; Wilson, C.B. Regulation of interferon-gamma during innate and adaptive immune responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar]
- Liu, Y.; Xu, J.; Fu, W.; Weng, Z.; Niu, X.; Liu, J.; Ding, X.; Zhang, Q. Tissues Expression, Polymorphisms of IFN Regulatory Factor 6 (IRF6) Gene and Their Associated with Immune Traits in Three Pig Populations. Asian-Australas. J. Anim. Sci. 2012, 25, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhou, M.; Gong, X.; Zhou, Y.; Chen, J.; Ma, J.; Zhang, P. Starch-protein interaction effects on lipid metabolism and gut microbes in host. Front. Nutr. 2022, 9, 1018026. [Google Scholar] [CrossRef]
- Oltean, M.; Gavrea, R.; Dumitrache, M.; Băguţ, T.; Gherman, C.M.; Cozma, V.; Györke, A. Characterization of host-parasite interactions during the experimental Trichinella spiralis infection in pigs. Helminthologia 2012, 49, 139–146. [Google Scholar] [CrossRef]
- Zhao, K.; Yin, H.; Yan, H.; Tang, W.; Diao, H.; Wang, Q.; Qi, R.; Liu, J. Dietary Supplementation of Lactobacillus johnsonii RS-7 Improved Antioxidant and Immune Function of Weaned Piglets. Animals 2023, 13, 1595. [Google Scholar] [CrossRef]
- Ding, Y.Y.; Zhu, M.Y.; Ding, J. Comparison and Analysis of Serum Biochemical indexes between Huoshou Black Pig and Berkshire Pig x Huoshou Black Pig at Different Days. Southwest China J. Agric. Sci. 2020, 33, 193–197. (In Chinese) [Google Scholar]
- Deutz, N.E.P.; Reijven, P.L.M.; Athanasas, G.; Soeters, P.B. Post-operative changes in hepatic, intestinal, splenic and muscle fluxes of amino acids and ammonia in pigs. Clin. Sci. 1992, 83, 607–614. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Xiang, X.; Zhang, Y.; Li, Q.; Wei, J.; Liu, K.; Shao, D.; Li, B.; Olszewski, M.A.; Ma, Z.; Qiu, Y. Expression profile of porcine scavenger receptor A and its role in bacterial phagocytosis by macrophages. Dev. Comp. Immunol. 2020, 104, 103534. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Qiu, H.; Zhang, Q.; Peng, Z.; Liu, B. Association of two porcine reproductive and respiratory syndrome virus (PRRSV) receptor genes, CD163 and SN with immune traits. Mol. Biol. Rep. 2012, 39, 3971–3976. [Google Scholar] [CrossRef] [PubMed]
- Burkard, C.; Opriessnig, T.; Mileham, A.J.; Stadejek, T.; Ait-Ali, T.; Lillico, S.G.; Whitelaw, C.B.A.; Archibald, A.L. Pigs Lacking the Scavenger Receptor Cysteine-Rich Domain 5 of CD163 Are Resistant to Porcine Reproductive and Respiratory Syndrome Virus 1 Infection. J. Virol. 2018, 92, e00415-18. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Martínez, P.; Yuste, M.; Poderoso, T.; Alonso, F.; Domínguez, J.; Ezquerra, A.; Revilla, C. Phenotypic and functional heterogeneity of CD169+ and CD163+ macrophages from porcine lymph nodes and spleen. Dev. Comp. Immunol. 2014, 44, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Ritter, M.; Orsó, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor CD163 expression in human monocytes and macrophages by pro- and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Aruffo, A.; Bowen, M.A.; Patel, D.D.; Haynes, B.F.; Starling, G.C.; Gebe, J.A.; Bajorath, J. CD6—Ligand interactions: A paradigm for SRCR domain function? Immunol. Today 1997, 18, 498–504. [Google Scholar] [CrossRef]
- Kraal, G.; van der Laan, L.J.; Elomaa, O.; Tryggvason, K. The macrophage receptor MARCO. Microbes Infect. 2000, 2, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Charo, I.F.; Ransohoff, R.M. The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Nalbantoglu, I.; Aitken, J.D.; Uchiyama, R.; Su, Y.; Doho, G.H.; Vijay-Kumar, M.; Gewirtz, A.T. Cytosolic flagellin receptor NLRC4 protects mice against mucosal and systemic challenges. Mucosal Immunol. 2012, 5, 288–298. [Google Scholar] [CrossRef]
- Lu, J.; Chatterjee, M.; Schmid, H.; Beck, S.; Gawaz, M. CXCL14 as an emerging immune and inflammatory modulator. J. Inflamm. 2016, 13, 1. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. The Chemokine Superfamily Revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef]
- Kanikarla Marie, P.; Fowlkes, N.W.; Afshar-Kharghan, V.; Martch, S.L.; Sorokin, A.V.; Shen, J.P.; Morris, V.K.; Dasari, A.; You, N.Y.; Sood, A.K.; et al. The Provocative Roles of Platelets in Liver Disease and Cancer. Front. Oncol. 2021, 11, 643815. [Google Scholar] [CrossRef]
- Laing, K.J.; Secombes, C.J. Chemokines. Dev. Comp. Immunol. 2004, 28, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Förster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Blander, J.M.; Medzhitov, R. Regulation of phagosome maturation by signals from toll-like receptors. Science 2004, 304, 1014–1018. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Zhou, S.; Wang, Y.; Shi, X.; Liu, H. Cadmium exposure activates the PI3K/AKT signaling pathway through miRNA-21, induces an increase in M1 polarization of macrophages, and leads to fibrosis of pig liver tissue. Ecotoxicol. Environ. Saf. 2021, 228, 113015. [Google Scholar] [CrossRef]
- Liang, W.; Ji, L.; Zhang, Y.; Zhen, Y.; Zhang, Q.; Xu, X.; Liu, B. Transcriptome Differences in Porcine Alveolar Macrophages from Tongcheng and Large White Pigs in Response to Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Infection. Int. J. Mol. Sci. 2017, 18, 1475. [Google Scholar] [CrossRef]
- Gao, Y.; Wahlberg, P.; Marthey, S.; Esquerré, D.; Jaffrézic, F.; Lecardonnel, J.; Hugot, K.; Rogel-Gaillard, C. Analysis of porcine MHC using microarrays. Vet. Immunol. Immunopathol. 2012, 148, 78–84. [Google Scholar] [CrossRef]
- Mair, K.H.; Sedlak, C.; Käser, T.; Pasternak, A.; Levast, B.; Gerner, W.; Saalmüller, A.; Summerfield, A.; Gerdts, V.; Wilson, H.L.; et al. The porcine innate immune system: An update. Dev. Comp. Immunol. 2014, 45, 321–343. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Hu, R.; Tang, T.; Tang, W.; Huang, C. Review of the correlation between Chinese medicine and intestinal microbiota on the efficacy of diabetes mellitus. Front. Endocrinol. 2022, 13, 1085092. [Google Scholar] [CrossRef]
- Park, B.-J.; Ahn, H.-S.; Han, S.-H.; Go, H.-J.; Kim, D.-H.; Choi, C.; Jung, S.; Myoung, J.; Lee, J.-B.; Park, S.-Y.; et al. Analysis of the Immune Responses in the Ileum of Gnotobiotic Pigs Infected with the Recombinant GII.p12_GII.3 Human Norovirus by mRNA Sequencing. Viruses 2021, 13, 92. [Google Scholar] [CrossRef]
- Pathak, S.K.; Kumar, A.; Bhuwana, G.; Sah, V.; Upmanyu, V.; Tiwari, A.K.; Sahoo, A.P.; Sahoo, A.R.; Wani, S.A.; Panigrahi, M.; et al. RNA Seq analysis for transcriptome profiling in response to classical swine fever vaccination in indigenous and crossbred pigs. Funct. Integr. Genom. 2017, 17, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Hardbower, D.M.; Singh, K.; Asim, M.; Verriere, T.G.; Olivares-Villagómez, D.; Barry, D.P.; Allaman, M.M.; Washington, M.K.; Peek, R.M., Jr.; Piazuelo, M.B.; et al. EGFR regulates macrophage activation and function in bacterial infection. J. Clin. Investig. 2016, 126, 3296–3312. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Li, J.; Xu, P.; Wang, L.; Feng, M.; Chen, D.; Yu, X.; Lu, Y. Molecular biology of BPIFB1 and its advances in disease. Ann. Transl. Med. 2020, 8, 651. [Google Scholar] [CrossRef] [PubMed]
- Ninsuwon, J.; Waiyamitra, P.; Roongsitthichai, A.; Surachetpong, W. Expressions of miR-155 and miR-181 and predictions of their structures and targets in pigs (Sus scrofa). Vet. World 2020, 13, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ouyang, M.; Van den Dries, K.; McGhee, E.J.; Tanaka, K.; Anderson, M.D.; Groisman, A.; Goult, B.T.; Anderson, K.I.; Schwartz, M.A. Correction: Talin tension sensor reveals novel features of focal adhesion force transmission and mechanosensitivity. J. Cell Biol. 2016, 214, 231. [Google Scholar] [CrossRef] [PubMed]
- Korytina, G.F.; Akhmadishina, L.Z.; Kochetova, O.V.; Aznabaeva, Y.G.; Zagidullin, S.Z.; Victorova, T.V. The Role of Serum Amyloid A1, Adhesion Molecules, Chemokines, and Chemokine Receptors Genes in Chronic Obstructive Pulmonary Disease. Russ. J. Genet. 2019, 55, 105–113. [Google Scholar] [CrossRef]
- Tang, Q.-h.; Zhang, Y.-m.; Xu, Y.-z.; He, L.; Dai, C.; Sun, P. Up-regulation of integrin beta3 expression in porcine vascular endothelial cells cultured in vitro by classical swine fever virus. Vet. Immunol. Immunopathol. 2010, 133, 237–242. [Google Scholar] [CrossRef]
- Li, W.; Wang, G.; Liang, W.; Kang, K.; Guo, K.; Zhang, Y. Integrin β3 Is Required in Infection and Proliferation of Classical Swine Fever Virus. PLoS ONE 2014, 9, e110911. [Google Scholar] [CrossRef]
- Yang, C.; Lan, R.; Wang, X.; Zhao, Q.; Li, X.; Bi, J.; Wang, J.; Yang, G.; Lin, Y.; Liu, J.; et al. Integrin β3, a RACK1 interacting protein, is critical for porcine reproductive and respiratory syndrome virus infection and NF-κB activation in Marc-145 cells. Virus Res. 2020, 282, 197956. [Google Scholar] [CrossRef]
- Rottman, J.B.; Ganley, K.P.; Williams, K.C.; Wu, L.; Mackay, C.R.; Ringler, D.J. Cellular localization of the chemokine receptor CCR5. Correlation to cellular targets of HIV-1 infection. Am. J. Pathol. 1997, 151, 1341–1351. [Google Scholar] [PubMed]
- Zhen, Y.; Wang, F.; Liang, W.; Liu, J.; Gao, G.; Wang, Y.; Xu, X.; Su, Q.; Zhang, Q.; Liu, B. Identification of Differentially Expressed Non-coding RNA in Porcine Alveolar Macrophages from Tongcheng and Large White Pigs Responded to PRRSV. Sci. Rep. 2018, 8, 15621. [Google Scholar] [CrossRef]
- Kim, S.; Lim, B.; Mattoo, S.u.S.; Oh, E.Y.; Jeong, C.-G.; Kim, W.-I.; Lee, K.-T.; Lee, S.-M.; Kim, J.-M. Comprehensive Transcriptomic Comparison between Porcine CD8− and CD8+ gamma Delta T Cells Revealed Distinct Immune Phenotype. Animals 2021, 11, 2165. [Google Scholar] [CrossRef]
- Wu, L.; Paxton, W.A.; Kassam, N.; Ruffing, N.; Rottman, J.B.; Sullivan, N.J.; Choe, H.; Sodroski, J.G.; Newman, W.; Koup, R.A.; et al. CCR5 Levels and Expression Pattern Correlate with Infectability by Macrophage-tropic HIV-1, In Vitro. J. Exp. Med. 1997, 185, 1681–1692. [Google Scholar] [CrossRef]
- Hariharan, D.; Douglas, S.D.; Lee, B.; Lai, J.-p.; Campbell, D.E.; Ho, W. Interferon-gamma upregulates CCR5 expression in cord and adult blood mononuclear phagocytes. Blood 1999, 93, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Patterson, B.K.; Czerniewski, M.A.; Andersson, J.; Sullivan, Y.B.; Su, F.; Jiyamapa, D.; Burki, Z.G.; Landay, A.L. Regulation of CCR5 and CXCR4 expression by type 1 and type 2 cytokines: CCR5 expression is downregulated by IL-10 in CD4-positive lymphocytes. Clin. Immunol. 1999, 91, 254–262. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Range | Sensitivity |
|---|---|---|
| IL-2 | 10~320 pg/mL | 1.0 pg/mL |
| IL-4 | 2~64 pg/mL | 0.1 pg/mL |
| IL-6 | 25~800 pg/mL | 1.0 pg/mL |
| IL-17 | 1.5~48 pg/mL | 0.1 pg/mL |
| IgA | 25~800 µg/mL | 1.0 µg/mL |
| TNF-α | 6.25~200 pg/mL | 1.0 pg/mL |
| IFN-γ | 1.25~40 pg/mL | 0.1 pg/mL |
| Gene | Primer Sequence (5′-3′) | Annealing Temperature (°C) | Amplicon Length (bp) | GenBank No. |
|---|---|---|---|---|
| CD163 | F: ATAGAGCTTGATGGGTCCGAGG | 60 | 321 | XM_021091123.1 |
| R: GCTCAACATTCTTGGGTCCTGG | ||||
| DYNC1I1 | F: GCTGGGGAAAGGAAGTGCTA | 60 | 186 | XM_021102406.1 |
| R: GTCAGACATGCTGGCTTCCT | ||||
| TLR4 | F: CAGATAAGCGAGGCCGTCATT | 60 | 113 | NM_001293316.1 |
| R: TTGCAGCCCACAAAAAGCA | ||||
| CXCL14 | F: GAATTCATGAGGCTCCTGGCGGCCG | 60 | 312 | NM_001244128.1 |
| R: GGATCCCTATTCTTCGTAGACCCT | ||||
| GPC1 | F: CTTAGTGCTGCTTTGCTTTTCAT | 60 | 155 | XM_047443961.1 |
| R: AGGGTTATTATGGGGTGGACTT | ||||
| KCNA3 | F: GAGCTGCACTGACCAGTAGG | 60 | 607 | XM_021086763.1 |
| R: GTGCTGGCAGATGGATCACT | ||||
| KCTD17 | F: GAGCTCACACAGATGGTATCC | 60 | 95 | XM_021091531.1 |
| R: TGGTCCTCACTCCCATAGTT | ||||
| ITGB3 | F: CACGGGCAAGTACTGTGAGT | 60 | 123 | NM_214002.1 |
| R: TAGAAGCCAGTCCAGTCGGA | ||||
| S100A14 | F: AAACTGGAGCTGTCTGTCGG | 60 | 173 | NM_001190168.1 |
| R: TGTAAATGCCCAAGCACCCA | ||||
| EPCAM | F: TGCTCTTTGAATGCGCTTGG | 60 | 172 | NM_214419.1 |
| R: AGAGCCCATCGTTGTTCTGG | ||||
| HAVCR2 | F: TTCGACGGGAGCAGTAAAGC | 60 | 105 | XM_003134109.4 |
| R: CAAGGGCAGGACACAGTCAA | ||||
| BPIFB1 | F: ACTCTTCGTGTCCAGTTTCTACT | 60 | 159 | NM_001101032.1 |
| R: TTCTGGACCGAGCCTGAGA | ||||
| β-Actin | F: GCGTAGCATTTGCTGCATGA | 60 | 252 | XM 003357928.4 |
| R: GCGTGTGTGTAACTAGGGGT |
| Items | (Mean ± SD) | ||
|---|---|---|---|
| Ningxiang | Berkshire | F1 Offspring of Ningxiang and Berkshires Pigs (F1 Offspring) | |
| plasma | |||
| IgA (ug/mL) | 732.87 ± 55.55 a | 653.13 ± 24.41 b | 726.54 ± 68.17 a |
| IL-2 (pg/mL) | 308.57 ± 23.18 a | 307.30 ± 22.41 a | 277.48 ± 20.83 b |
| IL-6 (pg/mL) | 806.59 ± 42.16 a | 751.83 ± 54.18 b | 768.85 ± 35.15 b |
| IFN-γ (pg/mL) | 36.96 ± 2.94 a | 33.31 ± 4.44 b | 37.64 ± 3.76 a |
| spleen | |||
| IL-4 (pg/mL) | 60.32 ± 4.45 a | 55.91 ± 4.56 b | 57.01 ± 5.02 b |
| IL-6 (pg/mL) | 755.00 ± 19.54 a | 740.01 ± 16.52 b | 750.82 ± 14.54 a,b |
| IL-17 (pg/mL) | 43.69 ± 2.41 a | 40.77 ± 3.27 b | 41.10 ± 4.75 b |
| IFN-γ (pg/mL) | 42.01 ± 1.86 a | 38.94 ± 3.21 b | 40.95 ± 1.53 a |
| TNF-α (pg/mL) | 195.08 ± 9.82 b | 203.01 ± 12.99 a | 207.59 ± 19.45 a |
| Items | (Mean ± SD) | ||
|---|---|---|---|
| Ningxiang | Berkshire | F1 Offspring | |
| ALB (g/L) | 60.02 ± 2.27 b | 70.86 ± 9.55 a | 60.66 ± 3.99 b |
| ALT (U/L) | 59.04 ± 11.48 b | 88.09 ± 23.96 a | 55.91 ± 13.58 b |
| ALP (U/L) | 31.29 ± 22.19 a | 27.57 ± 30.39 a | 5.29 ± 4.27 b |
| GLU (mmol/L) | 11.89 ± 2.27 a | 9.53 ± 1.86 b | 9.84 ± 3.15 b |
| CHOL (mmol/L) | 3.35 ± 0.59 a | 2.90 ± 0.63 b | 2.63 ± 0.27 b |
| LDL-C3 (mmol/L) | 2.24 ± 0.35 a | 1.78 ± 0.72 b | 1.56 ± 0.24 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, G.; Zhang, Y.; Gao, H.; Fu, Y.; Chen, Y.; Yin, Y.; Xu, K. Differences in Immune Characteristics and Related Gene Expression in Spleen among Ningxiang, Berkshire Breeds and Their Hybrid Pigs. Genes 2024, 15, 205. https://doi.org/10.3390/genes15020205
Song G, Zhang Y, Gao H, Fu Y, Chen Y, Yin Y, Xu K. Differences in Immune Characteristics and Related Gene Expression in Spleen among Ningxiang, Berkshire Breeds and Their Hybrid Pigs. Genes. 2024; 15(2):205. https://doi.org/10.3390/genes15020205
Chicago/Turabian StyleSong, Gang, Yuebo Zhang, Hu Gao, Yawei Fu, Yue Chen, Yulong Yin, and Kang Xu. 2024. "Differences in Immune Characteristics and Related Gene Expression in Spleen among Ningxiang, Berkshire Breeds and Their Hybrid Pigs" Genes 15, no. 2: 205. https://doi.org/10.3390/genes15020205
APA StyleSong, G., Zhang, Y., Gao, H., Fu, Y., Chen, Y., Yin, Y., & Xu, K. (2024). Differences in Immune Characteristics and Related Gene Expression in Spleen among Ningxiang, Berkshire Breeds and Their Hybrid Pigs. Genes, 15(2), 205. https://doi.org/10.3390/genes15020205