
Discovery, Pathogenesis, and Complete Genome Characterization of Lates calcarifer Herpesvirus
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Barramundi Sample Collection and Preparation
2.1.1. Field Sampling Sets
2.1.2. Field Sample Collection
2.1.3. Field Sample Processing and Homogenization
2.2. Virus Culture on SBB Cells and Virus Titration
2.2.1. Sea Bass Brain (SBB) Cell Culture
2.2.2. Virus Culture
2.2.3. Endpoint Dilution/Titration Assays
2.3. Library Preparation, NGS, and Phylogenetic Inference
2.3.1. VIDISCA-NGS
2.3.2. Illumina Sequencing
2.3.3. Oxford Nanopore Sequencing
2.3.4. Genome Assembly and Comparison
2.3.5. Open Reading Frames Prediction and Annotation
2.3.6. Phylogenetic Inference
2.4. Quantitative Polymerase Chain Reaction (qPCR)
2.4.1. DNA Extraction
2.4.2. Primer and Probe Design
2.4.3. qPCR Master Mix and Program
2.4.4. Conventional PCR and Gel Electrophoresis
2.4.5. Standards for Quantification
2.5. Electron Microscopy
2.6. Experimental LCHV Infection in Barramundi
2.6.1. Experimental Infection Sample Collection
2.6.2. Experimental Infection Sample Processing and Homogenization
2.7. Histology
3. Results
3.1. Index Outbreak
3.2. Virus Culture from Affected Fish
3.3. Virus Identification
3.4. LCHV Genome Characterization and Phylogeny
3.5. Association with Disease
3.6. Experimental Infection of LCHV V511 in L. calcarifer and Reproduction of the Clinical Signs
3.7. Confirmation of LCHV Infection in Experimentally Infected Fish/Cohabitant Fish
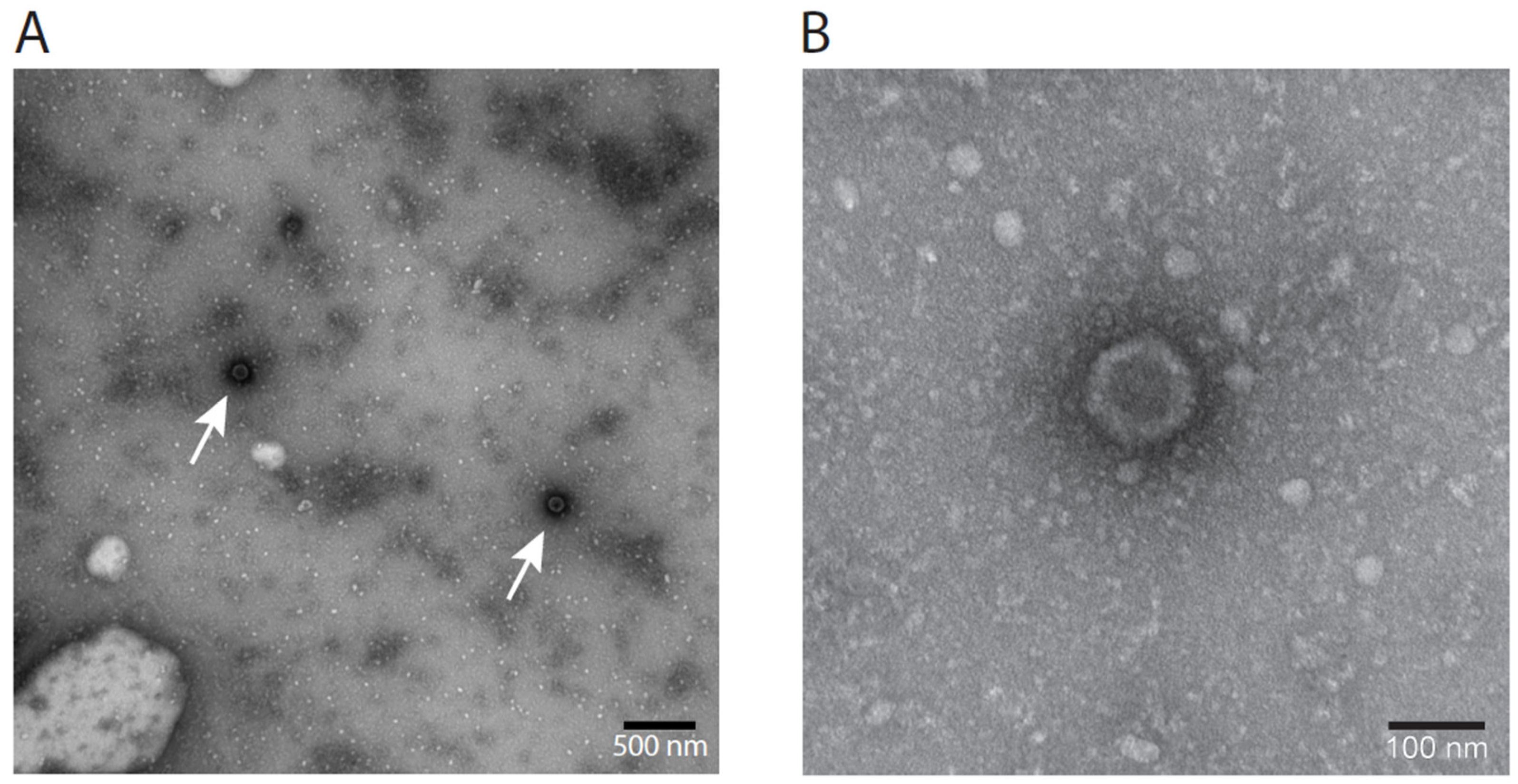
3.8. Electron Microscopy Visualization of LCHV
3.9. Histopathological Analysis of Experimentally Infected Fish
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rimmer, M.A. Lates calcarifer—Cultured Aquatic Species Fact Sheets; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; Available online: https://www.fao.org/fishery/docs/DOCUMENT/aquaculture/CulturedSpecies/file/en/en_barramundi.htm (accessed on 30 January 2024).
- Global Aquaculture Production Quantity (1950–2021). Available online: https://www.fao.org/fishery/statistics-query/en/aquaculture/aquaculture_quantity (accessed on 30 January 2024).
- Mohd Yazid, S.H.; Mohd Daud, H.; Azmai, M.N.A.; Mohamad, N.; Mohd Nor, N. Estimating the economic loss due to vibriosis in net-cage cultured asian seabass (Lates calcarifer): Evidence from the east coast of peninsular malaysia. Front. Vet. Sci. 2021, 8, 644009. [Google Scholar] [CrossRef]
- Avendano-Herrera, R.; Toranzo, A.E.; Magarinos, B. Tenacibaculosis infection in marine fish caused by tenacibaculum maritimum: A review. Dis. Aquat. Organ. 2006, 71, 255–266. [Google Scholar] [CrossRef]
- de Groof, A.; Guelen, L.; Deijs, M.; van der Wal, Y.; Miyata, M.; Ng, K.S.; van Grinsven, L.; Simmelink, B.; Biermann, Y.; Grisez, L.; et al. A novel virus causes scale drop disease in lates calcarifer. PLoS Pathog. 2015, 11, e1005074. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Kueh, S.; Chee, D.; Chen, J.; Wang, Y.H.; Tay, S.; Leong, L.N.; Ng, M.L.; Jones, J.B.; Nicholls, P.K.; Ferguson, H.W. The pathology of ‘scale drop syndrome’ in asian seabass, lates calcarifer bloch, a first description. J. Fish. Dis. 2012, 35, 19–27. [Google Scholar] [CrossRef]
- Kurita, J.; Nakajima, K. Megalocytiviruses. Viruses 2012, 4, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Toh, X.; Ong, J.; Wang, Y.; Teo, X.H.; Lee, B.; Wong, P.S.; Khor, D.; Chong, S.M.; Chee, D.; et al. Detection and characterization of a novel marine birnavirus isolated from asian seabass in Singapore. Virol. J. 2019, 16, 71. [Google Scholar] [CrossRef]
- Chang, S.F.; Ng, K.S.; Grisez, L.; De Groof, A.; Vogels, W.; van der Hoek, L.; Deijs, M. Novel Fish Pathogenic Virus. International Publication. WO2018029301A1, 15 February 2018. [Google Scholar]
- Meemetta, W.; Domingos, J.A.; Dong, H.T.; Senapin, S. Development of a SYBR green quantitative PCR assay for detection of lates calcarifer herpesvirus (LCHV) in farmed barramundi. J. Virol. Methods 2020, 285, 113920. [Google Scholar] [CrossRef]
- Domingos, J.A.; Shen, X.; Terence, C.; Senapin, S.; Dong, H.T.; Tan, M.R.; Gibson-Kueh, S.; Jerry, D.R. Scale drop disease virus (SDDV) and lates calcarifer herpes virus (LCHV) coinfection downregulate immune-relevant pathways and cause splenic and kidney necrosis in barramundi under commercial farming conditions. Front. Genet. 2021, 12, 666897. [Google Scholar] [CrossRef]
- Dang, M.; Dien, T.D.; Van, U.P.; Ha, V.T.; Dung, V.V.; Hieu, N.T.D.; Hua, V.C.; Hue, N.T.K.; Giang, N.T.T.; Truong, V.H.; et al. The first description of histopathology of lates calcarifer herpesvirus (LCHV) infection in barramundi (Lates calcarifer). Aquaculture 2023, 565, 739091. [Google Scholar] [CrossRef]
- Hanson, L.; Dishon, A.; Kotler, M. Herpesviruses that infect fish. Viruses 2011, 3, 2160–2191. [Google Scholar] [CrossRef]
- Gatherer, D.; Depledge, D.P.; Hartley, C.A.; Szpara, M.L.; Vaz, P.K.; Benkő, M.; Brandt, C.R.; Bryant, N.A.; Dastjerdi, A.; Doszpoly, A.; et al. ICTV virus taxonomy profile: Herpesviridae 2021. J. Gen. Virol. 2021, 102, 001673. [Google Scholar] [CrossRef] [PubMed]
- Sano, N.; Moriwake, M.; Hondo, R.; Sano, T. Herpesvirus cyprini: A search for viral genome in infected fish by infected fish by in situ hybridization. J. Fish Dis. 2006, 16, 495–499. [Google Scholar] [CrossRef]
- Davison, A.J.; Cunningham, C.; Sauerbier, W.; McKinnell, R.G. Genome sequences of two frog herpesviruses. J. Gen. Virol. 2006, 87, 3509–3514. [Google Scholar] [CrossRef] [PubMed]
- Origgi, F.C.; Schmidt, B.R.; Lohmann, P.; Otten, P.; Akdesir, E.; Gaschen, V.; Aguilar-Bultet, L.; Wahli, T.; Sattler, U.; Stoffel, M.H. Ranid herpesvirus 3 and proliferative dermatitis in free-ranging wild common frogs (Rana temporaria). Vet. Pathol. 2017, 54, 686–694. [Google Scholar] [CrossRef]
- Origgi, F.C.; Schmidt, B.R.; Lohmann, P.; Otten, P.; Meier, R.K.; Pisano, S.R.R.; Moore-Jones, G.; Tecilla, M.; Sattler, U.; Wahli, T.; et al. Bufonid herpesvirus 1 (BfHV1) associated dermatitis and mortality in free ranging common toads (Bufo bufo) in switzerland. Sci. Rep. 2018, 8, 14737. [Google Scholar] [CrossRef]
- van Beurden, S.J.; Bossers, A.; Voorbergen-Laarman, M.H.A.; Haenen, O.L.M.; Peters, S.; Abma-Henkens, M.H.C.; Peeters, B.P.H.; Rottier, P.J.M.; Engelsma, M.Y. Complete genome sequence and taxonomic position of anguillid herpesvirus 1. J. Gen. Virol. 2010, 91, 880–887. [Google Scholar] [CrossRef]
- DDavison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative genomics of carp herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef]
- Waltzek, T.B.; Subramaniam, K.; Doszpoly, A.; Hughes, J.; Vučak, M.; Davison, A.J. Genome sequence of white sturgeon herpesvirus 1 isolated from farmed white sturgeon (Acipenser transmontanus). Microbiol. Resour. Announc. 2023, 12, e0057123. [Google Scholar] [CrossRef]
- Walker, L.; Subramaniam, K.; Viadanna, P.; Vann, J.; Marcquenski, S.; Godard, D.; Kieran, E.; Frasca, S.; Popov, V.; Kerr, K.; et al. Characterization of an alloherpesvirus from wild lake sturgeon Acipenser fulvescens in Wisconsin (USA). Dis. Aquat. Organ. 2022, 149, 83–96. [Google Scholar] [CrossRef]
- Johnston, A.E.; Shavalier, M.A.; Scribner, K.T.; Soto, E.; Griffin, M.J.; Waldbieser, G.C.; Richardson, B.M.; Winters, A.D.; Yun, S.; Baker, E.A.; et al. First isolation of a herpesvirus (family Alloherpesviridae) from great lakes lake sturgeon (Acipenser fulvescens). Animals 2022, 12, 3230. [Google Scholar] [CrossRef]
- Du, K.; Stöck, M.; Kneitz, S.; Klopp, C.; Woltering, J.M.; Adolfi, M.C.; Feron, R.; Prokopov, D.; Makunin, A.; Kichigin, I.; et al. The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization. Nat. Ecol. Evol. 2020, 4, 841–852. [Google Scholar] [CrossRef]
- Davison, A.J. Channel catfish virus: A new type of herpesvirus. Virology 1992, 186, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Borzák, R.; Haluk, T.; Bartha, D.; Doszpoly, A. Complete genome sequence and analysis of ictalurid herpesvirus 2. Arch. Virol. 2018, 163, 1083–1085. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, R.N.; Polchana, J.; Kanchanakhan, S.; Davison, A.J.; Waltzek, T.B.; Subramaniam, K. Genome sequence of a novel alloherpesvirus isolated from glass catfish (Kryptopterus bicirrhis). Genome Announc. 2018, 6, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Doszpoly, A.; Somogyi, V.; LaPatra, S.E.; Benkő, M. Partial genome characterization of acipenserid herpesvirus 2: Taxonomical proposal for the demarcation of three subfamilies in Alloherpesviridae. Arch. Virol. 2011, 156, 2291–2296. [Google Scholar] [CrossRef]
- Gilad, O.; Yun, S.; Andree, K.B.; Adkison, M.A.; Zlotkin, A.; Bercovier, H.; Eldar, A.; Hedrick, R.P. Initial characteristics of koi herpesvirus and development of a polymerase chain reaction assay to detect the virus in koi, Cyprinus carpio koi. Dis. Aquat. Organ. 2002, 48, 101–108. [Google Scholar] [CrossRef]
- Wolf, K.; Darlington, R.W. Channel catfish virus: A new herpesvirus of ictalurid fish. J. Virol. 1971, 8, 525–533. [Google Scholar] [CrossRef]
- Hasoon, M.F.; Daud, H.M.; Abdullah, A.A.; Arshad, S.S.; Bejo, H.M. Development and partial characterization of new marine cell line from brain of asian sea bass lates calcarifer for virus isolation. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 16–25. [Google Scholar] [CrossRef]
- Chi, S.C.; Wu, Y.C.; Cheng, T.M. Persistent infection of betanodavirus in a novel cell line derived from the brain tissue of barramundi lates calcarifer. Dis. Aquat. Organ. 2005, 65, 91–98. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Boom, R.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Wertheim-van Dillen, P.M.; van der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef]
- de Vries, M.; Deijs, M.; Canuti, M.; van Schaik, B.D.; Faria, N.R.; van de Garde, M.D.; Jachimowski, L.C.; Jebbink, M.F.; Jakobs, M.; Luyf, A.C.; et al. A sensitive assay for virus discovery in respiratory clinical samples. PLoS ONE 2011, 6, e16118. [Google Scholar] [CrossRef]
- Endoh, D.; Mizutani, T.; Kirisawa, R.; Maki, Y.; Saito, H.; Kon, Y.; Morikawa, S.; Hayashi, M. Species-independent detection of rna virus by representational difference analysis using non-ribosomal hexanucleotides for reverse transcription. Nucleic Acids Res. 2005, 33, e65. [Google Scholar] [CrossRef]
- Edridge, A.W.D.; Deijs, M.; Namazzi, R.; Cristella, C.; Jebbink, M.F.; Maurer, I.; A Kootstra, N.; Buluma, L.R.; van Woensel, J.B.M.; de Jong, M.D.; et al. Novel orthobunyavirus identified in the cerebrospinal fluid of a ugandan child with severe encephalopathy. Clin. Infect. Dis. 2019, 68, 139–142. [Google Scholar] [CrossRef]
- De Coster, W.; Rademakers, R. Nanopack2: Population-scale evaluation of long-read sequencing data. Bioinformatics 2023, 39, btad311. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Pfaff, F.; Neubauer-Juric, A.; Krebs, S.; Hauser, A.; Singer, S.; Blum, H.; Hoffmann, B. Full genome sequence of bovine alphaherpesvirus 2 (BoHV-2). Arch. Virol. 2021, 166, 639–643. [Google Scholar] [CrossRef]
- Needleman, S.B.; Wunsch, C.D. A general method applicable to the search for similarities in the amino acid sequence of two proteins. J. Mol. Biol. 1970, 48, 443–453. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. Eggnog-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. Orthofinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef]
- Buchfink, B.; Reuter, K.; Drost, H.-G. Sensitive protein alignments at tree-of-life scale using diamond. Nat. Methods 2021, 18, 366–368. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. Mafft online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. Iq-tree 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Wildy, P.; Russell, W.; Horne, R. The morphology of herpes virus. Virology 1960, 12, 204–222. [Google Scholar] [CrossRef]
- Evans, A.S. Causation and disease: The henle-koch postulates revisited. Yale J. Biol. Med. 1976, 49, 175–195. [Google Scholar]
- Lepa, A.; Siwicki, A.K. Fish herpesvirus diseases: A short review of current knowledge. Acta Veterinaria Brno 2012, 81, 383–389. [Google Scholar] [CrossRef]
- Corbeil, S. Abalone viral ganglioneuritis. Pathogens 2020, 9, 720. [Google Scholar] [CrossRef]
- Savin, K.W.; Cocks, B.G.; Wong, F.; Sawbridge, T.; Cogan, N.; Savage, D.; Warner, S. A neurotropic herpesvirus infecting the gastropod, abalone, shares ancestry with oyster herpesvirus and a herpesvirus associated with the amphioxus genome. Virol. J. 2010, 7, 308. [Google Scholar] [CrossRef]
- Stehle, T.; Schulz, G.E. Refined structure of the complex between guanylate kinase and its substrate GMP at 2.0 a resolution. J. Mol. Biol. 1992, 224, 1127–1141. [Google Scholar] [CrossRef] [PubMed]
- Mabrok, M.; Algammal, A.M.; Sivaramasamy, E.; Hetta, H.F.; Atwah, B.; Alghamdi, S.; Fawzy, A.; Avendaño-Herrera, R.; Rodkhum, C. Tenacibaculosis caused by Tenacibaculum maritimum: Updated knowledge of this marine bacterial fish pathogen. Front. Cell Infect. Microbiol. 2022, 12, 1068000. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Post-Challenge | |||||
|---|---|---|---|---|---|
| 4 | 7 | 11 | 14 | 17 or 18 a | |
| Intraperitoneal injection | |||||
| Gills | 0 b | nt | nt | nt | 36,214 |
| Heart | nt | nt | nt | nt | 5154 |
| Intestine | nt | nt | nt | nt | 3481 |
| Brain | nt | nt | nt | nt | 582 |
| Liver | nt | nt | nt | nt | 1628 |
| Skin | nt | nt | nt | nt | 39,032 |
| Spleen | nt | nt | nt | nt | nt |
| Kidney | 30,167 | 19,204 | 262 | 46 | 214 |
| Serum | nt | nt | nt | nt | 21,056 |
| Cohabitant infection | |||||
| Gills | 403 | 80 | 0 | 0 | 15,060 |
| Heart | 533 | 31 | 0 | 0 | 2930 |
| Intestine | 2731 | 7 | 0 | 0 | 29 |
| Brain | 234 | 65 | 20 | 50 | 25 |
| Liver | 836 | 0 | 0 | 0 | 162 |
| Skin | 748,800 | 7 | 7 | 831 | 16,111 |
| Spleen | nt | 0 | 0 | 0 | nt |
| Kidney | 12 | 85 | 0 | 0 | 6 |
| Serum | 0 | 0 | 0 | 0 | 730 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simmelink, B.; Coolen, J.P.M.; Vogels, W.; Deijs, M.; van der Last-Kempkes, J.L.M.; Ng, K.S.; Chang, S.F.; Gevers, K.; Harkema, L.; van der Hoek, L.; et al. Discovery, Pathogenesis, and Complete Genome Characterization of Lates calcarifer Herpesvirus. Genes 2024, 15, 264. https://doi.org/10.3390/genes15030264
Simmelink B, Coolen JPM, Vogels W, Deijs M, van der Last-Kempkes JLM, Ng KS, Chang SF, Gevers K, Harkema L, van der Hoek L, et al. Discovery, Pathogenesis, and Complete Genome Characterization of Lates calcarifer Herpesvirus. Genes. 2024; 15(3):264. https://doi.org/10.3390/genes15030264
Chicago/Turabian StyleSimmelink, Bartjan, Jordy P. M. Coolen, Wannes Vogels, Martin Deijs, Jessica L. M. van der Last-Kempkes, Kah Sing Ng, Siow Foong Chang, Koen Gevers, Liesbeth Harkema, Lia van der Hoek, and et al. 2024. "Discovery, Pathogenesis, and Complete Genome Characterization of Lates calcarifer Herpesvirus" Genes 15, no. 3: 264. https://doi.org/10.3390/genes15030264
APA StyleSimmelink, B., Coolen, J. P. M., Vogels, W., Deijs, M., van der Last-Kempkes, J. L. M., Ng, K. S., Chang, S. F., Gevers, K., Harkema, L., van der Hoek, L., & de Groof, A. (2024). Discovery, Pathogenesis, and Complete Genome Characterization of Lates calcarifer Herpesvirus. Genes, 15(3), 264. https://doi.org/10.3390/genes15030264