

Predictive Role of Cluster Bean (Cyamopsis tetragonoloba) Derived miRNAs in Human and Cattle Health

 , ,
, ,
Abstract
:1. Introduction
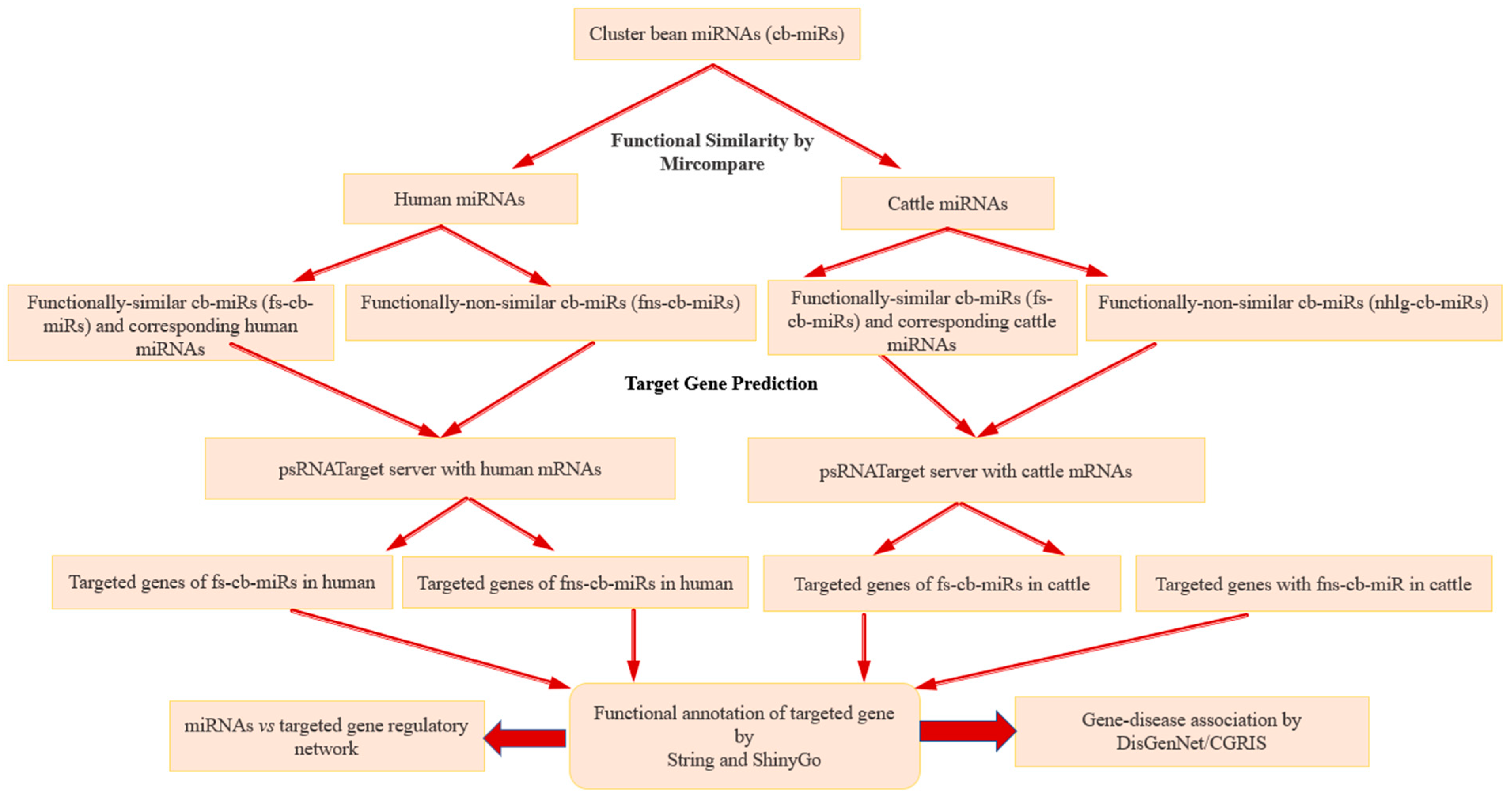
2. Material and Methods
2.1. Data Source
2.2. Cross-Kingdom miRNA Similarity
2.3. Identification of Functionally Similar cbmiRs (fs-cb-miRs) and Prediction of Potential Target Genes in Humans and Cattle
2.4. Functional Annotation and Pathway Analysis of cb-miRs’ Targeted Genes
2.5. Disease Association with cb-miRs’ Target Genes
3. Results
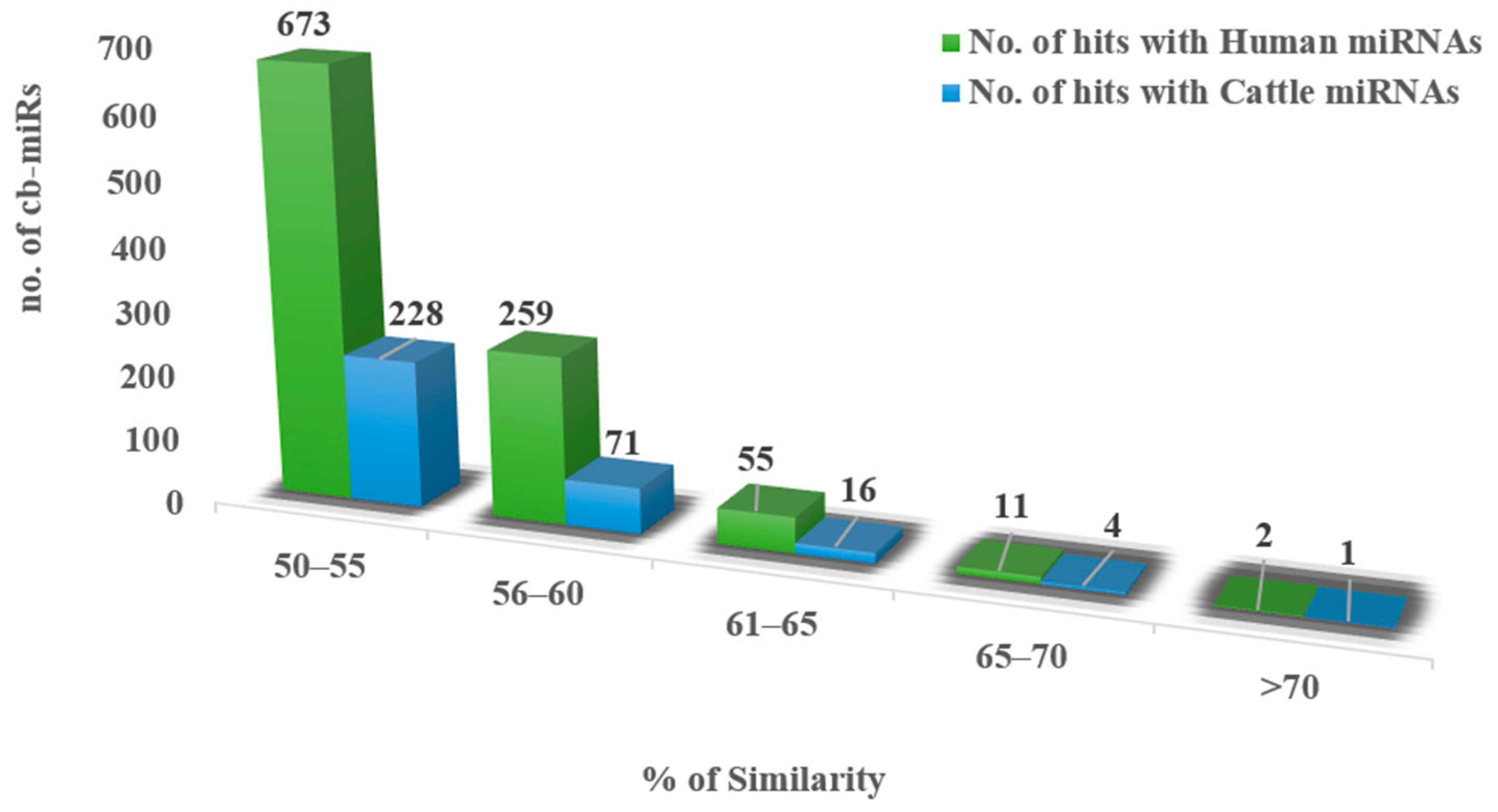
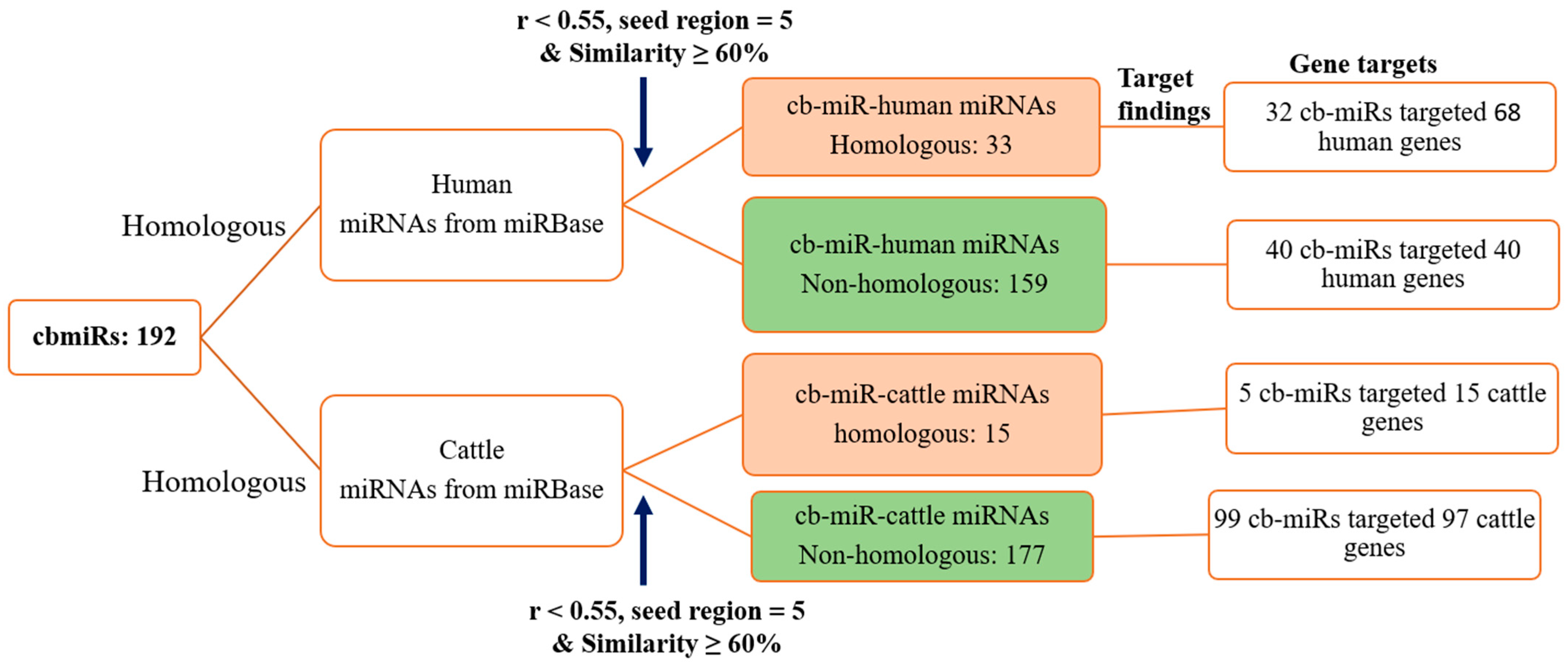
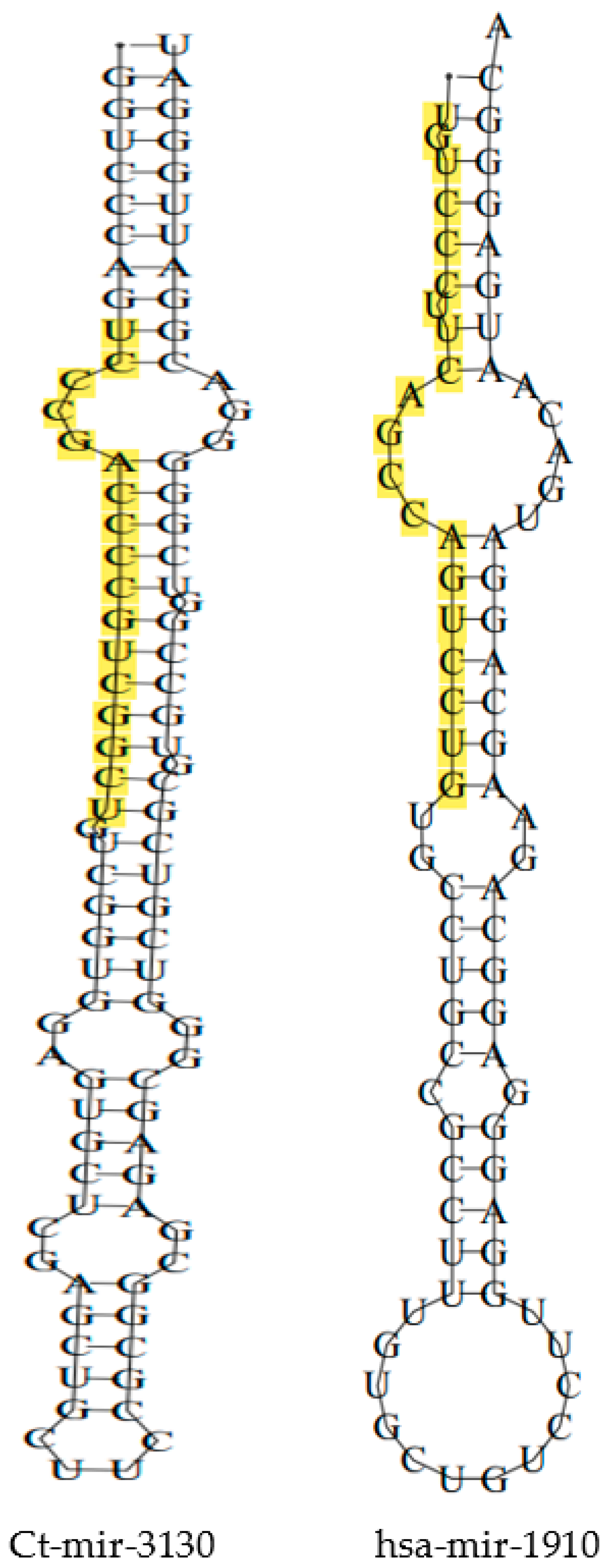
3.1. Identification of fs-cb-miRs and fns-cb-miRs to Human and Cattle miRNAs
3.2. Prediction of Target Genes of cb-miRs in Human and Cattle
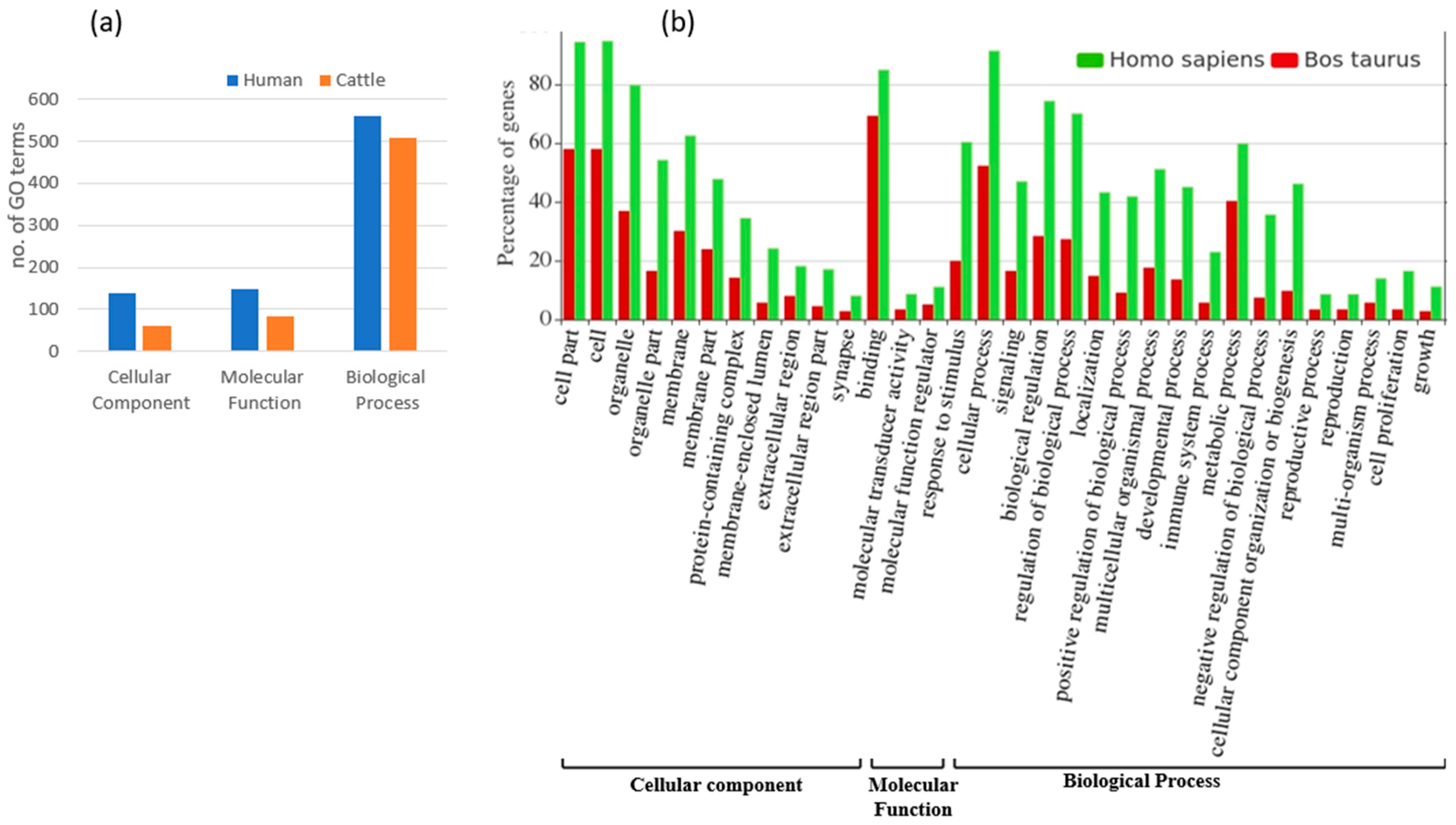
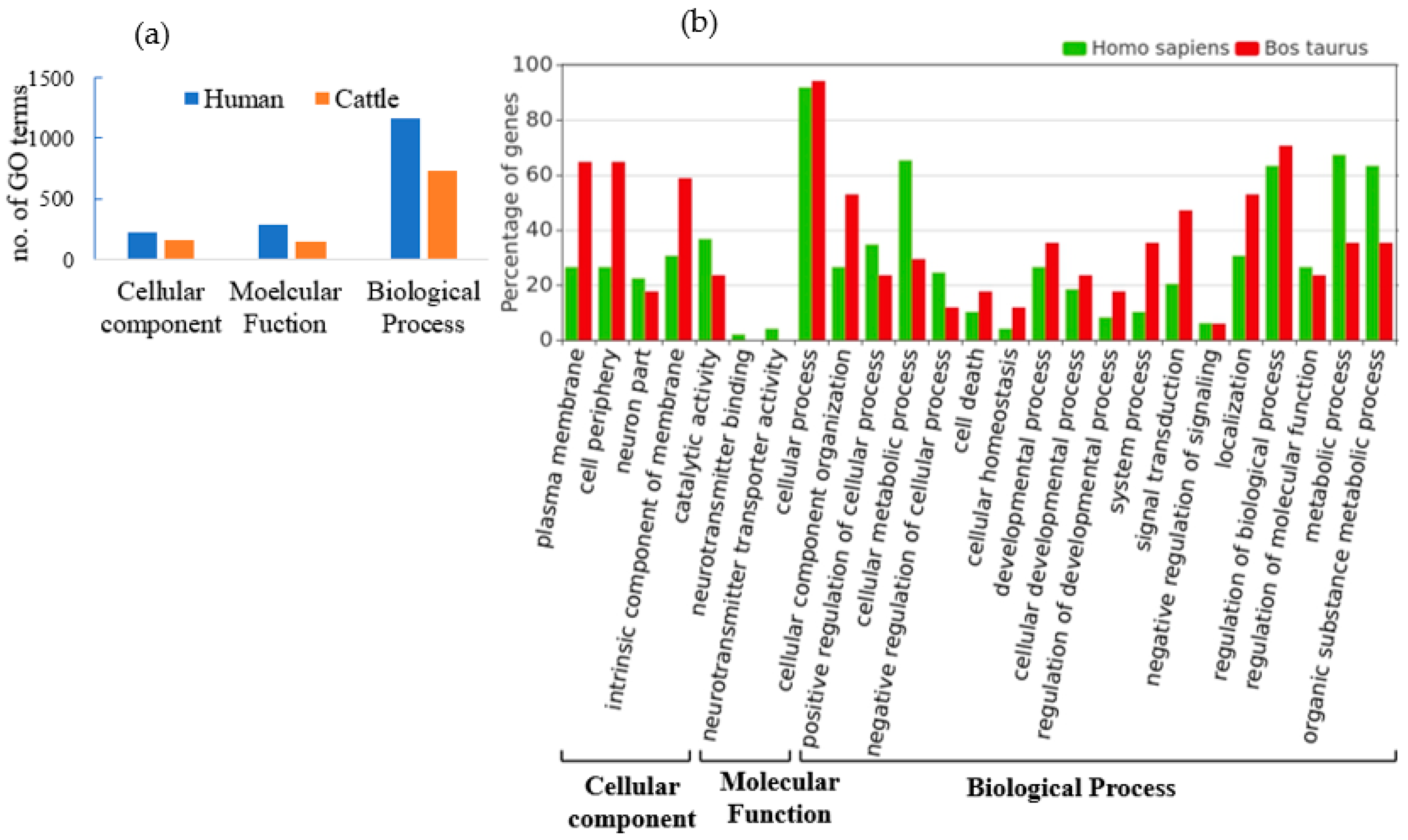
3.3. Functional Annotation and Pathway Analysis of Human and Cattle Genes Targeted by cb-miRs
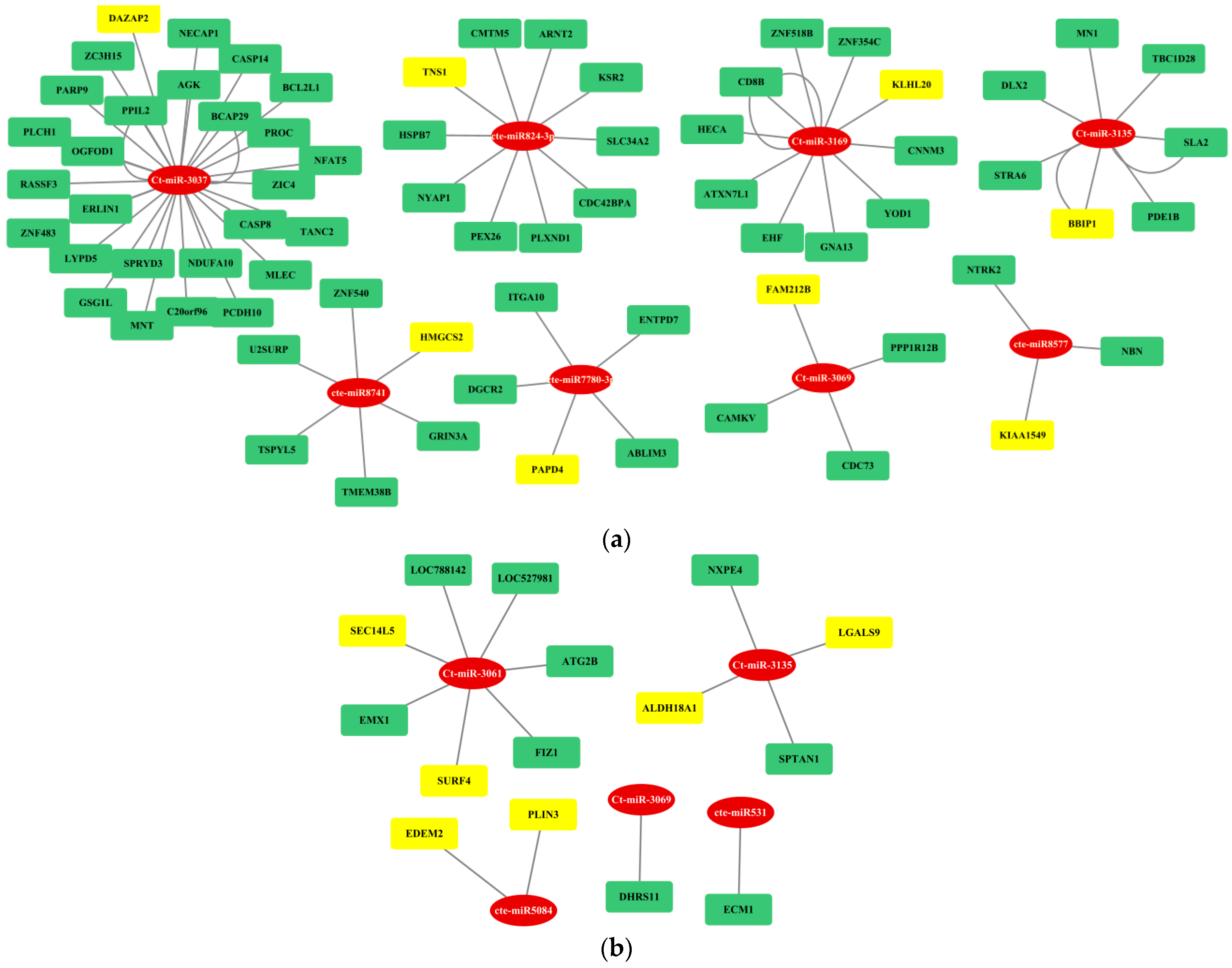
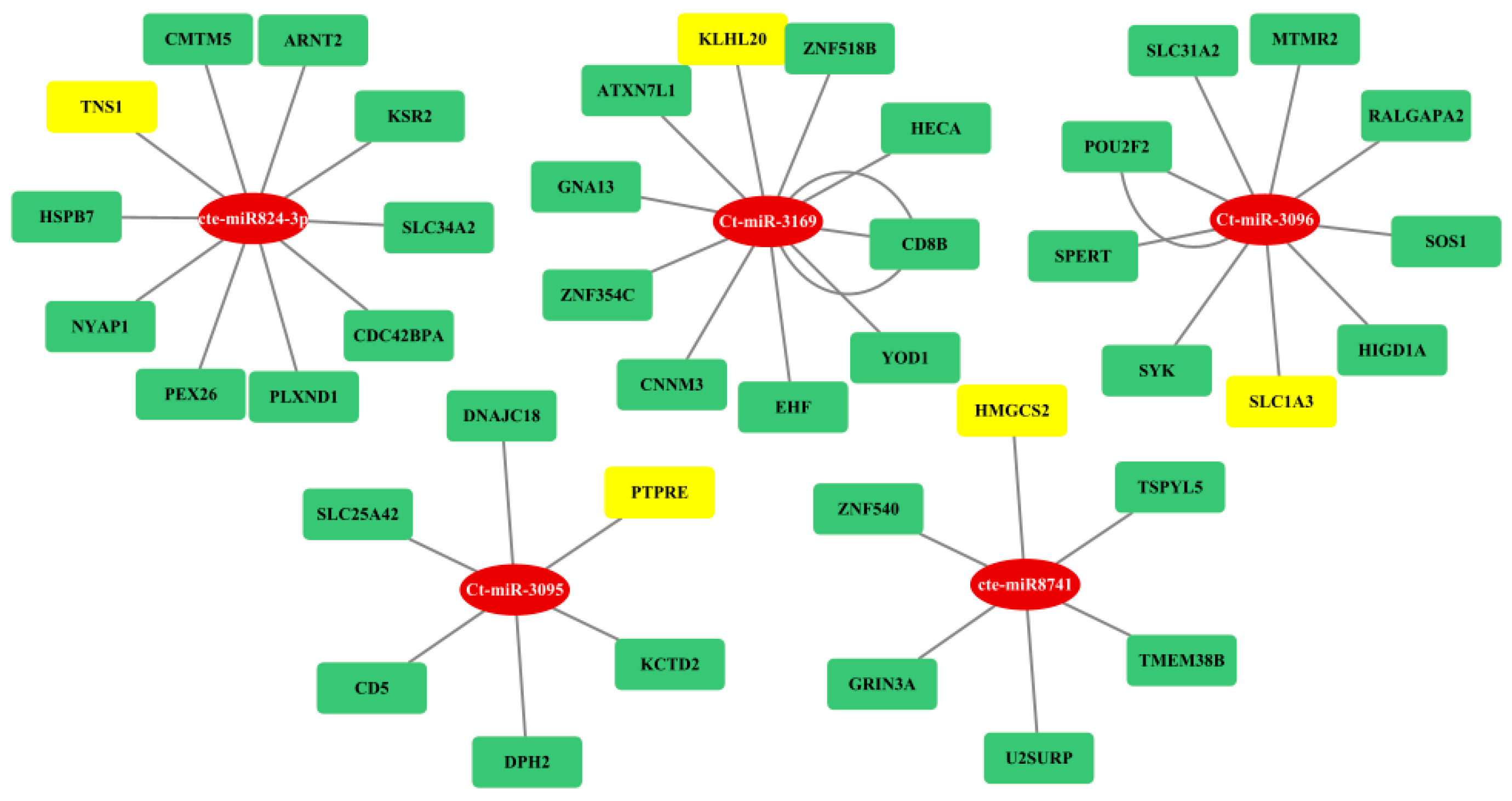
3.4. Gene Regulatory Network Analysis
3.5. Association of Target Genes of fs-cb-miRs and fns-cb-miRs with Human and Cattle Diseases
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, D.T.; Sperandio, V. Inter-kingdom signalling: Communication between bacteria and their hosts. Nat. Rev. Microbiol. 2008, 6, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Zen, K.; Zhang, J.; Zhang, C.Y.; Chen, X. New roles for microRNAs in cross-species communication. RNA Biol. 2013, 10, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Silva, M.R.; das Neves, R.F.; Cabrera-Cabrera, F.; Sanguinetti, J.; Medeiros, L.C.; Robello, C.; Naya, H.; Fernandez-Calero, T.; Souto-Padron, T.; de Souza, W.; et al. Extracellular vesicles shed by Trypanosoma cruzi are linked to small RNA pathways, life cycle regulation, and susceptibility to infection of mammalian cells. Parasitol. Res. 2014, 113, 285–304. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Wang, H.D.; Wu, J.; Ren, J.; Meng, L.; Wu, Q.; Dong, H.; Wu, J.; Kao, T.Y.; et al. Escherichia coli noncoding RNAs can affect gene expression and physiology of Caenorhabditis elegans. Nat. Commun. 2012, 3, 1073. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef]
- Ivashuta, S.I.; Petrick, J.S.; Heisel, S.E.; Zhang, Y.; Guo, L.; Reynolds, T.L.; Rice, J.F.; Allen, E.; Roberts, J.K. Endogenous small RNAs in grain: Semi-quantification and sequence homology to human and animal genes. Food Chem. Toxicol. 2009, 47, 353–360. [Google Scholar] [CrossRef]
- Bhatt, D.H.; Jha, N.; KJSr Pandya, H.A. In silico exploration of miRNA from EST data of avocado and predicting its cross-kingdom effects on human. Pharma. Innov. J. 2017, 6, 543–548. [Google Scholar]
- Köberle, V.; Pleli, T.; Schmithals, C.; Augusto Alonso, E.A.; Haupenthal, J.; Bönig, H.; Peveling-Oberhag, J.; Biondi, R.M.; Zeuzem, S.; Kronenberger, B.; et al. Differential stability of cell-free circulating microRNAs: Implications for their utilization as biomarkers. PLoS ONE 2013, 8, e75184. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sahu, S.; Kumari, P.; Gopi, S.R.; Malhotra, R.; Biswas, S. Genome-wide identification and functional annotation of miRNAs in anti-inflammatory plant and their cross-kingdom regulation in Homo sapiens. J. Biomol. Struct. Dyn. 2017, 35, 1389–1400. [Google Scholar] [CrossRef]
- Zhou, L.K.; Zhou, Z.; Jiang, X.M.; Zheng, Y.; Chen, X.; Fu, Z.; Xiao, G.; Zhang, C.Y.; Zhang, L.K.; Yi, Y. Absorbed plant MIR2911 in honeysuckle decoction inhibits SARS-CoV-2 replication and accelerates the negative conversion of infected patients. Cell Discov. 2020, 6, 54. [Google Scholar] [CrossRef]
- Ji, L.; Chen, X. Regulation of small RNA stability: Methylation and beyond. Cell Res. 2012, 22, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Minutolo, A.; Potestà, M.; Gismondi, A.; Pirrò, S.; Cirilli, M.; Gattabria, F.; Galgani, A.; Sessa, L.; Mattei, M.; Canini, A.; et al. Olea europaea small RNA with functional homology to human miR34a in cross-kingdom interaction of anti-tumoral response. Sci. Rep. 2018, 8, 12413. [Google Scholar] [CrossRef]
- Lamonte, G.; Philip, N.; Reardon, J.; Lacsina, J.R.; Majoros, W.; Chapman, L.; Thornburg, C.D.; Telen, M.J.; Ohler, U.; Nicchitta, C.V.; et al. Translocation of sickle cell erythrocyte microRNAs into Plasmodium falciparum inibits parasite translation and contributes to malaria resistance. Cell Host Microbe 2012, 12, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; Van Eijndhoven, M.A.J.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Würdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef]
- Pastrello, C.; Tsay, M.; McQuaid, R.; Abovsky, M.; Pasini, E.; Shirdel, E.; Angeli, M.; Tokar, T.; Jamnik, J.; Kotlyar, M.; et al. Circulating plant miRNAs can regulate human gene expression in vitro. Sci. Rep. 2016, 6, 32773. [Google Scholar] [CrossRef]
- Wang, M.L.; Morris, J.B. Flavonoid content in seeds of guar germplasm using HPLC. Plant Genet. Res. 2007, 5, 96–99. [Google Scholar] [CrossRef]
- Gaikwad, K.; Ramakrishna, G.; Srivastava, H.; Saxena, S.; Kaila, T.; Tyagi, A.; Sharma, P.; Sharma, S.; Sharma, R.; Mahla, H.R.; et al. The chromosome-scale genome assembly of cluster bean provides molecular insight into edible gum (galactomannan) biosynthesis family genes. Sci. Rep. 2023, 13, 9941. [Google Scholar] [CrossRef]
- Mukhtar, H.M.; Ansari, S.H.; Bhat, Z.A.; Naved, T. Antihyperglycemic activity of Cyamopsis tetragonoloba beans on blood glucose levels in alloxan-induced diabetic rats. Pharm. Biol. 2006, 44, 10–13. [Google Scholar] [CrossRef]
- Tyagi, A.; Nigam, D.; SV, A.M.; Solanke, A.U.; Singh, N.K.; Sharma, T.R.; Gaikwad, K. Genome-wide discovery of tissue-specific miRNAs in clusterbean (Cyamopsis tetragonoloba) indicates their association with galactomannan biosynthesis. Plant Biotechnol. J. 2018, 16, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.; Rao, A.R.; Pandey, J.; Gaikwad, K.; Ghoshal, S.; Mohapatra, T. Genome-wide identification and characterization of lncRNAs and miRNAs in cluster bean (Cyamopsis tetragonoloba). Gene 2018, 667, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. PsRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Pirrò, S.; Minutolo, A.; Galgani, A.; Potestà, M.; Colizzi, V.; Montesano, C. Bioinformatics prediction and experimental validation of microRNAs involved in cross-kingdom interaction. J. Comput. Biol. 2016, 23, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.J.; Chakraborty, S.; Deb, B.; Rajwanshi, R. Computational identification and functional annotation of microRNAs and their targets from expressed sequence tags (ESTs) and genome survey sequences (GSSs) of coffee (Coffea arabica L.). Plant Gene 2016, 1, 30–42. [Google Scholar] [CrossRef]
- Supriya, P.; Kumar, A.; Archak, S.; Bhat, K.V. Computational identification of microRNAs and their target genes in sesame (Sesamum indicum L.). Indian J. Genet. Plant Breed. 2022, 82, 469–473. [Google Scholar]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 15, 2628–2629. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Kanehisa, M. The KEGG database. Novartis Found. Symp. 2002, 247, 91–101. [Google Scholar] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zhang, Y.; Cui, H.; Liu, J.; Wu, Y.; Cheng, Y.; Xu, H.; Huang, X.; Li, S.; Zhou, A.; et al. WEGO 2.0: A web tool for analyzing and plotting GO annotations, 2018 update. Nucleic Acids Res. 2018, 46, W71–W75. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef] [PubMed]
- Strande, N.T.; Riggs, E.R.; Buchanan, A.H.; Ceyhan-Birsoy, O.; DiStefano, M.; Dwight, S.S.; Goldstein, J.; Ghosh, R.; Seifert, B.A.; Sneddon, T.P.; et al. Evaluating the clinical validity of gene-disease associations: An evidence-based framework developed by the clinical genome resource. Am. J. Hum. Genet. 2017, 100, 895–906. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, S.; Ayachit, G.; Bhairappanavar, S.B.; Ansari, A.; Sharma, P.; Soni, S.; Das, J. Cross-kingdom regulation of putative miRNAs derived from happy tree in cancer pathway: A systems biology approach. Int. J. Mol. Sci. 2017, 18, 1191. [Google Scholar] [CrossRef]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef]
- Li, J.; Hu, W.X.; Luo, S.Q.; Xiong, D.H.; Sun, S.; Wang, Y.P.; Bu, X.F.; Liu, J.; Hu, J. Promoter methylation induced epigenetic silencing of DAZAP2, a downstream effector of p38/MAPK pathway, in multiple myeloma cells. Cell Signal 2019, 60, 136–145. [Google Scholar] [CrossRef]
- Chen, Z.; Picaud, S.; Filippakopoulos, P.; D’Angiolella, V.; Bullock, A.N. Structural Basis for Recruitment of DAPK1 to the KLHL20 E3 Ligase. Structure 2019, 27, 1395–1404.e4. [Google Scholar] [CrossRef] [PubMed]
- Kotepui, M.; Thawornkuno, C.; Chavalitshewinkoon-Petmitr, P.; Punyarit, P.; Petmitr, S. Quantitative Real-Time RT-PCR of ITGA7, SVEP1, TNS1, LPHN3, SEMA3G, KLB and MMP13 mRNA Expression in Breast Cancer. Asian Pac. J. Cancer Prev. 2012, 13, 5879–5882. [Google Scholar] [CrossRef] [PubMed]
- Scheidecker, S.; Etard, C.; Pierce, N.W.; Geoffroy, V.; Schaefer, E.; Muller, J.; Chennen, K.; Flori, E.; Pelletier, V.; Poch, O.; et al. Exome sequencing of Bardet-Biedl syndrome patient identifies a null mutation in the BBSome subunit BBIP1 (BBS18). J. Med. Genet. 2014, 51, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Reid, E.S.; Papandreou, A.; Drury, S.; Boustred, C.; Yue, W.W.; Wedatilake, Y.; Beesley, C.; Jacques, T.S.; Anderson, G.; Abulhoul, L.; et al. Advantages and pitfalls of an extended gene panel for investigating complex neurometabolic phenotypes. Brain J. Neurol. 2016, 139, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Ulveling, D.; Le Clerc, S.; Cobat, A.; Labib, T.; Noirel, J.; Laville, V.; Coulonges, C.; Carpentier, W.; Nalpas, B.; Heim, M.H.; et al. A new 3p25 locus is associated with liver fibrosis progression in human immunodeficiency virus/hepatitis C virus-coinfected patients. Hepatology 2016, 64, 1462–1472. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, Y.; Kawai, Y.; Naito, T.; Hirano, A.; Umeno, J.; Fuyuno, Y.; Liu, Z.; Li, D.; Nakano, T.; Izumiyama, Y.; et al. A Genome-wide Association Study Identifying RAP1A as a Novel Susceptibility Gene for Crohn’s Disease in Japanese Individuals. J. Crohn’s Colitis 2021, 13, 648–658. [Google Scholar] [CrossRef]
- Appay, R.; Fina, F.; Macagno, N.; Padovani, L.; Colin, C.; Barets, D.; Ordioni, J.; Scavarda, D.; Giangaspero, F.; Badiali, M.; et al. Duplications of KIAA1549 and BRAF screening by Droplet Digital PCR from formalin-fixed paraffin-embedded DNA is an accurate alternative for KIAA1549-BRAF fusion detection in pilocytic astrocytomas. Mod. Pathol. 2018, 31, 1490–1501. [Google Scholar] [CrossRef]
- Vine, A.E.; McQuillin, A.; Bass, N.J.; Pereira, A.; Kandaswamy, R.; Robinson, M.; Lawrence, J.; Anjorin, A.; Sklar, P.; Gurling, H.M.; et al. No evidence for excess runs of homozygosity in bipolar disorder. Psychiatr. Genet. 2009, 19, 165–170. [Google Scholar] [CrossRef]
- Han, X.; Huang, S.; Xue, P.; Fu, J.; Liu, L.; Zhang, C.; Yang, L.; Xia, L.; Sun, L.; Huang, S.K.; et al. LncRNA PTPRE-AS1 modulates M2 macrophage activation and inflammatory diseases by epigenetic promotion of PTPRE. Sci. Adv. 2019, 5, eaax9230. [Google Scholar] [CrossRef]
- Fan, S.J.; Snell, C.; Turley, H.; Li, J.L.; McCormick, R.; Perera, S.M.; Heublein, S.; Kazi, S.; Azad, A.; Wilson, C.; et al. PAT4 levels control amino-acid sensitivity of rapamycin-resistant mTORC1 from the Golgi and affect clinical outcome in colorectal cancer. Oncogene 2016, 35, 3004–3015. [Google Scholar] [CrossRef]
- AbdulHameed, M.D.; Tawa, G.J.; Kumar, K.; Ippolito, D.L.; Lewis, J.A.; Stallings, J.D.; Wallqvist, A. Systems level analysis and identification of pathways and networks associated with liver fibrosis. PLoS ONE 2014, 9, e112193. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, S.; Sidiq, T.; Yousuf, S.; van den Elsen, P.J.; Kobayashi, K.S. Class I transactivator, NLRC5: A central player in the MHC class I pathway and cancer immune surveillance. Immunogenetics 2019, 71, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Plenge, R.M.; Seielstad, M.; Padyukov, L.; Lee, A.T.; Remmers, E.F.; Ding, B.; Liew, A.; Khalili, H.; Chandrasekaran, A.; Davies, L.R.; et al. TRAF1-C5 as a risk locus for rheumatoid arthritis—A genomewide study. N. Engl. J. Med. 2007, 357, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Bradbury, N.A. Lemur Tyrosine Kinase 2, a novel target in prostate cancer therapy. Oncotarget 2015, 6, 14233–14246. [Google Scholar] [CrossRef] [PubMed]
- Schomisch, S.J.; Murdock, D.G.; Hedayati, N.; Carino, J.L.; Lesnefsky, E.J.; Cmolik, B.L. Cardioplegia prevents ischemia-induced transcriptional alterations of cytoprotective genes in rat hearts: A DNA microarray study. J. Thorac. Cardiovasc. Surg. 2005, 130, 1151. [Google Scholar] [CrossRef] [PubMed]
- Cusanovich, D.A.; Billstrand, C.; Zhou, X.; Chavarria, C.; De Leon, S.; Michelini, K.; Pai, A.A.; Ober, C.; Gilad, Y. The combination of a genome-wide association study of lymphocyte count and analysis of gene expression data reveals novel asthma candidate genes. Hum. Mol. Genet. 2012, 21, 2111–2123. [Google Scholar] [CrossRef] [PubMed]
- Makise, N.; Sekimizu, M.; Kobayashi, E.; Yoshida, H.; Fukayama, M.; Kato, T.; Kawai, A.; Ichikawa, H.; Yoshida, A. Low-grade endometrial stromal sarcoma with a novel MEAF6-SUZ12 fusion. Virchows Arch. Int. J. Pathol. 2019, 475, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.D.; Jen, J.C.; Choi, S.Y.; Shin, J.H.; Kim, H.S.; Kim, H.J.; Kim, J.S.; Choi, J.H. Late-onset episodic ataxia associated with SLC1A3 mutation. J. Hum. Genet. 2017, 62, 443–446. [Google Scholar] [CrossRef]
- Albash, B.; Imtiaz, F.; Al-Zaidan, H.; Al-Manea, H.; Banemai, M.; Allam, R.; Al-Suheel, A.; Al-Owain, M. Novel PHKG2 mutation causing GSD IX with prominent liver disease: Report of three cases and review of literature. Eur. J. Pediatr. 2014, 173, 647–653. [Google Scholar] [CrossRef]
- Ogushi, K.; Hattori, A.; Suzuki, E.; Shima, H.; Izawa, M.; Yagasaki, H.; Horikawa, R.; Uetake, K.; Umezawa, A.; Ishii, T.; et al. DNA Methylation Status of SHOX-Flanking CpG Islands in Healthy Individuals and Short Stature Patients with Pseudoautosomal Copy Number Variations. Cytogenet. Genome Res. 2019, 158, 56–62. [Google Scholar] [CrossRef]
- Rosales-Reynoso, M.A.; Ochoa-Hernández, A.B.; Aguilar-Lemarroy, A.; Jave-Suárez, L.F.; Troyo-Sanromán, R.; Barros-Núñez, P. Gene expression profiling identifies WNT7A as a possible candidate gene for decreased cancer risk in fragile X syndrome patients. Arch. Med. Res. 2010, 41, 110–118.e2. [Google Scholar] [CrossRef] [PubMed]
- Cali, F.; Ragalmuto, A.; Chiavetta, V.; Calabrese, G.; Fichera, M.; Vinci, M.; Ruggeri, G.; Schinocca, P.; Sturnio, M.; Romano, S.; et al. Novel deletion of the E3A ubiquitin protein ligase gene detected by multiplex ligation-dependent probe amplification in a patient with Angelman syndrome. Exp. Mol. Med. 2010, 12, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Tickoo, M.; Gupta, R.; Vaswani, M.; Ambekar, A.; Grover, T.; Sharma, A. Association of serotonin and GABA pathway gene polymorphisms with alcohol dependence: A preliminary study. Asian J. Psychiatry 2019, 39, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Loor, J.J.; Dann, H.M.; Everts, R.E.; Oliveira, R.; Green, C.A.; Guretzky, N.A.; Rodriguez-Zas, S.L.; Lewin, H.A.; Drackley, J.K. Temporal gene expression profiling of liver from periparturient dairy cows reveals complex adaptive mechanisms in hepatic function. Physiol. Genom. 2005, 23, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lee, J.E.; Kang, J.W.; Shin, H.Y.; Lee, J.B.; Jin, D.I. Endoplasmic reticulum (ER) stress and unfolded protein response (UPR) in mammalian oocyte maturation and preimplantation embryo development. Int. J. Mol. Sci. 2019, 20, 409. [Google Scholar] [CrossRef] [PubMed]
- Goo, Y.A. Stromal Mesenchyme Cell Genes in Prostate Cancer Development: Epigenetic Markers for Cancer and Potential Targets for Therapy. Univ. Washington Report, 2006. Available online: https://apps.dtic.mil/sti/citations/ADA467588(accessed on 29 June 2023).
- Wurmser, C.; Tardif, S.; Price, M.; Küry, P. Developmental disorders associated with EMX1 mutations. Mol. Syndromol. 2017, 8, 41–47. [Google Scholar]
- Rupp, R.; Hernandez, A.; Mallard, B.A.; Brockman, F.J. Association of variation in the SEC14L5 gene with susceptibility to mastitis in dairy cattle. J. Dairy Sci. 2019, 102, 8083–8094. [Google Scholar]
- Van der Stede, Y.; Vanderbeke, E.; Bauwens, W.; Mintiens, K.; De Leeuw, I. Association between DHRS11 and susceptibility to bovine viral diarrhea virus infection in cattle. Vet. Res. 2018, 49, 1–8. [Google Scholar]
- McGill, J.L.; Kelly, S.M.; Guerra-Maupome, M.; Prewitt, A.K.; Bass, B.E. Galectin-9 upregulation in the lungs of cattle with bovine respiratory disease contributes to neutrophil and CD4+ T cell recruitment. Vet. Immunol. Immunopathol. 2020, 228, 110090. [Google Scholar]
- Kametani, Y.; Kawamoto, T.; Furuichi, T.; Ikegawa, S.; Okazaki, Y. A novel homozygous ALDH18A1 mutation in a patient with hereditary spastic paraplegia and developmental delay. Gene 2014, 544, 137–142. [Google Scholar]
- Casas, E.; Lunstra, D.D.; Stone, R.T.; Franke, D.E.; Kappes, S.M.; Beever, J.E. A missense mutation in the bovine SPTAN1 gene causes congenital cerebellar ataxia in Angus cattle. BMC Genet. 2003, 4, 18. [Google Scholar]
- Kim, J.H.; Yoon, D.H.; Kim, B.H.; Lim, D.J.; Kim, Y.M.; Lee, S.S. Associations of SNPs in genes related to reproduction and milk production traits in Holstein cows. Asian-Australas. J. Anim. Sci. 2021, 34, 1306–1313. [Google Scholar]
- Neibergs, H.L.; Seabury, C.M.; Wojtowicz, A.J.; Wang, Z.; Scraggs, E.; Kiser, J.N.; Neupane, M.; Womack, J.E.; Eenennaam, A.V.; Hagevoort, G.R.; et al. Susceptibility loci revealed for bovine respiratory disease complex in pre-weaned holstein calves. BMC Genom. 2014, 15, 1164. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Pal, K.; Sharma, A.K.; Dutta, S.K.; Lau, J.S.; Yan, I.K.; Wang, E.; Elkhanany, A.; Alkharfy, K.M.; Sanyal, A.; et al. GAIP interacting protein C-terminus regulates autophagy and exosome biogenesis of pancreatic cancer through metabolic pathways. PLoS ONE 2014, 9, e114409. [Google Scholar] [CrossRef] [PubMed]
- Hojo, T.; Al-Zi’abi, M.O.; Komiyama, J.; Manabe, N.; Acosta, T.J.; Okuda, K. Expression and localization of cFLIP, an anti-apoptotic factor, in the bovine corpus luteum. J. Reprod. Dev. 2010, 56, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Groneberg, D.A.; Quarcoo, D.; Frossard, N.; Fischer, A. Neurogenic mechanisms in bronchial inflammatory diseases. Allergy 2004, 59, 1139–1152. [Google Scholar] [CrossRef]
- Hayes, H.C.; Popescu, P.; Dutrillaux, B. Comparative gene mapping of lactoperoxidase, retinoblastoma, and alpha-lactalbumin genes in cattle, sheep, and goats. Mamm. Genome 1993, 4, 593–597. [Google Scholar] [CrossRef]
- Rossier, B.C. Epithelial Sodium Channel (ENaC) and the Control of Blood Pressure. Curr. Opin. Pharmacol. 2015, 21, 76–87. [Google Scholar] [CrossRef]
- Jiang, M.; Sang, X.; Hong, Z. Beyond nutrients: Food-derived microRNAs provide cross-kingdom regulation. Bioessays 2012, 34, 280–284. [Google Scholar] [CrossRef]
- Zhang, S.; Sang, X.; Hou, D.; Chen, J.; Gu, H.; Zhang, Y.; Li, J.; Yang, D.; Zhu, H.; Yang, X.; et al. Plant-derived RNAi therapeutics: A strategic inhibitor of HBsAg. Biomaterials 2019, 210, 83–93. [Google Scholar] [CrossRef]
- Li, D.; Yang, J.; Yang, Y.; Liu, J.; Li, H.; Li, R.; Cao, C.; Shi, L.; Wu, W.; He, K. A timely review of cross-kingdom regulation of plant-derived MicroRNAs. Front. Genet. 2021, 12, 613197. [Google Scholar] [CrossRef] [PubMed]
- Saiyed, A.N.; Vasavada, A.R.; Johar, S.K. Recent trends in miRNA therapeutics and the application of plant miRNA for prevention and treatment of human diseases. Future J. Pharm. Sci. 2022, 1, 24. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Sanchita, R.S.; Singh, R.; Srivastava, G.; Sharma, A. Comparative Study of Withanolide Biosynthesis-Related miRNAs in Root and Leaf Tissues of Withania somnifera. Appl. Biochem. Biotechnol. 2018, 185, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Philip, A.; Ferro, V.A.; Tate, R.J. Determination of the potential bioavailability of plant microRNAs using a simulated human digestion process. Mol. Nutr. Food Res. 2015, 59, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Zhuang, X.; Wang, Q.; Jiang, H.; Deng, Z.B.; Wang, B.; Zhang, L.; Kakar, S.; Jun, Y.; Miller, D.; et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. Mol. Nutr. Food Res. 2014, 58, 1561–1573. [Google Scholar] [CrossRef] [PubMed]
- Beltrami, C.; Clayton, A.; Newbury, L.J.; Corish, P.; Jenkins, R.H.; Phillips, A.O.; Fraser, D.J.; Bowen, T. Stabilization of urinary microRNAs by association with exosomes and argonaute 2 protein. Non-Coding RNA 2015, 1, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Min, H.; Yoon, S. Got target?: Computational methods for microRNA target prediction and their extension. Exp. Mol. Med. 2010, 42, 233–244. [Google Scholar] [CrossRef]
- Patel, M.; Mangukia, N.; Jha, N.; Gadhavi, H.; Shah, K.; Patel, S.; Mankad, A.; Pandya, H.; Rawal, R. Computational identification of miRNA and their cross kingdom targets from expressed sequence tags of Ocimum basilicum. Mol. Biol. Rep. 2019, 1, 2979–2995. [Google Scholar] [CrossRef]
- Bellato, M.; De Marchi, D.; Gualtieri, C.; Sauta, E.; Magni, P.; Macovei, A.; Pasotti, L. A bioinformatics approach to explore microRNAs as tools to bridge pathways between plants and animals. Is DNA damage response (DDR) a potential target process? Front. Plant Sci. 2019, 10, 1535. [Google Scholar] [CrossRef]
- Gismondi, A.; Nanni, V.; Monteleone, V.; Colao, C.; Di Marco, G.; Canini, A. Plant miR171 modulates mTOR pathway in HEK293 cells by targeting GNA12. Mol. Biol. Rep. 2021, 48, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Halder, L.D.; Babych, S.; Palme, D.I.; Mansouri-Ghahnavieh, E.; Ivanov, L.; Ashonibare, V.; Langenhorst, D.; Prusty, B.; Rambach, G.; Wich, M.; et al. Candida albicans induces cross-kingdom miRNA trafficking in human monocytes to promote fungal growth. mBio 2022, 13, e03563-21. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romo, D.; Hernández-Vásquez, C.I.; Pereyra-Alférez, B.; García-García, J.H. Identification of potential target genes in Homo sapiens, by miRNA of Triticum aestivum: A cross kingdom computational approach. Non-Coding RNA Res. 2022, 7, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Karlebach, G.; Shamir, R. Modelling and analysis of gene regulatory networks. Nat. Rev. Mol. Cell Biol. 2008, 9, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Park, G.C.; Kim, J.M.; Shin, S.C.; Cheon, Y.I.; Sung, E.S.; Lee, M.; Lee, J.C.; Lee, B.J. Tensin Regulates Fundamental Biological Processes by Interacting with Integrins of Tonsil-Derived Mesenchymal Stem Cells. Cells 2022, 11, 2333. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Chen, L.; Guo, S. Knockdown of SLC34A2 inhibits cell proliferation, metastasis, and elevates chemosensitivity in glioma. J. Cell. Biochem. 2019, 120, 10205–10214. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, L.J.; Fan, C.C.; Chang, H.C.; Shih, H.A.; Min, M.Y.; Chang, M.S. Important roles of Vilse in dendritic architecture and synaptic plasticity. Sci. Rep. 2017, 7, 45646. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, Z.; Bassel-Duby, R.; Olson, E.N. Genetic and epigenetic regulation of cardiomyocytes in development, regeneration and disease. Development 2018, 145, dev171983. [Google Scholar] [CrossRef]
- Vos, M.J.; Zijlstra, M.P.; Kanon, B.; van Waarde-Verhagen, M.A.; Brunt, E.R.; Oosterveld-Hut, H.M.; Carra, S.; Sibon, O.C.; Kampinga, H.H. HSPB7 is the most potent polyQ aggregation suppressor within the HSPB family of molecular chaperones. Hum. Mol. Genet. 2010, 19, 4677–4693. [Google Scholar] [CrossRef]
- Shao, L.; Cui, Y.; Li, H.; Liu, Y.; Zhao, H.; Wang, Y.; Zhang, Y.; Ng, K.M.; Han, W.; Ma, D.; et al. CMTM5 exhibits tumor suppressor activities and is frequently silenced by methylation in carcinoma cell lines. Clin. Cancer Res. 2007, 13, 5756–5762. [Google Scholar] [CrossRef]
- Oh, W.J.; Gu, C. The role and mechanism-of-action of Sema3E and Plexin-D1 in vascular and neural development. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2013; Volume 24, pp. 156–162. [Google Scholar]
- Bruick, R.K.; McKnight, S.L. Differential interaction of the hypoxia-inducible factor-alpha (HIF-alpha) subunits with ARNT2. Mol. Cell Biol. 1995, 15, 4073–4082. [Google Scholar]
- East, M.P.; Christopher, R.M.A. CDC42BPA/MRCK [alpha]: A kinase target for brain, ovarian and skin cancers. Nat. Rev. Drug Discov. 2021, 20, 167–168. [Google Scholar] [CrossRef]
- Gadhavi, H.; Patel, M.; Mangukia, N.; Shah, K.; Bhadresha, K.; Patel, S.K.; Rawal, R.M.; Pandya, H.A. Transcriptome-wide miRNA identification of Bacopa monnieri: A cross-kingdom approach. Plant Signal. Behav. 2020, 2, 1699265. [Google Scholar] [CrossRef]
- Trivedi, T.S.; Mangukia, N.; Bhavsar, M.; Mankad, A.U.; Rawal, R.M.; Patel, S.K. A novel insight of Picrorhiza kurroa miRNAs in human cystic fibrosis: A transcriptome-wide cross-kingdom study. Hum. Gene 2023, 35, 201153. [Google Scholar] [CrossRef]
- Zhao, X.; Mo, D.; Li, A.; Gong, W.; Xiao, S.; Zhang, Y.; Li, J. Identification and characterization of long non-coding RNAs in subcutaneous adipose tissue from castrated and intact full-sib pair Huai pig breeds. Genomics 2018, 110, 190–199. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Gene | KEGG Id | Pathway |
|---|---|---|
| HMGCS2 | hsa00072 | Synthesis and degradation of ketone bodies |
| hsa00280 | Valine, leucine and isoleucine degradation | |
| hsa00650 | Butanoate metabolism | |
| hsa00900 | Terpenoid backbone biosynthesis | |
| hsa01100 | Metabolic pathways | |
| hsa03320 | PPAR signaling pathway | |
| UBE2K | hsa04120 | Ubiquitin mediated proteolysis |
| MYH14 | hsa04270 | Vascular smooth muscle contraction |
| hsa05130 | Pathogenic Escherichia coli infection | |
| hsa04530 | Tight junction | |
| LCP2 | hsa04810 | Regulation of actin cytoskeleton |
| hsa04015 | Rap1 signaling pathway | |
| hsa04380 | Osteoclast differentiation | |
| hsa05135 | Yersinia infection | |
| hsa04611 | Platelet activation | |
| hsa04650 | Natural killer cell mediated cytotoxicity | |
| hsa04660 | T cell receptor signaling pathway | |
| hsa04664 | Fc epsilon RI signaling pathway | |
| GABRA6 | hsa04723 | Retrograde endocannabinoid signaling |
| hsa04727 | GABAergic synapse | |
| hsa04742 | Taste transduction | |
| hsa05032 | Morphine addiction | |
| hsa05033 | Nicotine addiction | |
| hsa04080 | Neuroactive ligand-receptor interaction |
| Targeted Gene | KEGG Id | Pathway |
|---|---|---|
| ALDH18A1 | bta00330 | Arginine and proline metabolism |
| bta01100 | Metabolic pathways | |
| bta01230 | Biosynthesis of amino acids | |
| ATG2B | bta04136 | Autophagy—other |
| bta04140 | Autophagy—animal | |
| bta05010 | Alzheimer disease | |
| bta05014 | Amyotrophic lateral sclerosis | |
| bta05016 | Huntington disease | |
| bta05017 | Spinocerebellar ataxia | |
| DHRS11 | bta00140 | Steroid hormone biosynthesis |
| EDEM2 | bta04141 | Protein processing in endoplasmic reticulum |
| SPTAN1 | bta04210 | Apoptosis |
| Targeted Gene | KEGG Id | Pathway |
|---|---|---|
| ABI2 | hsa04810 | Regulation of actin cytoskeleton |
| COL22A1 | hsa04974 | Protein digestion and absorption |
| DNTT | hsa04640 | Hematopoietic cell lineage |
| hsa03450 | Non-homologous end-joining | |
| FLI1 | hsa05202 | Transcriptional misregulation in cancer |
| FMO3 | hsa00982 | Drug metabolism—cytochrome P450 |
| GABRA6 | hsa04727 | GABAergic synapse |
| hsa05032 | Morphine addiction | |
| hsa04080 | Neuroactive ligand-receptor interaction | |
| hsa05033 | Nicotine addiction | |
| hsa04723 | Retrograde endocannabinoid signaling | |
| hsa04742 | Taste transduction | |
| HMGCS2 | hsa00650 | Butanoate metabolism |
| hsa01100 | Metabolic pathways | |
| hsa03320 | PPAR signaling pathway | |
| hsa00072 | Synthesis and degradation of ketone bodies | |
| hsa00900 | Terpenoid backbone biosynthesis | |
| hsa00280 | Valine, leucine and isoleucine degradation | |
| LCP2 | hsa04664 | Fc epsilon RI signaling pathway |
| hsa04650 | Natural killer cell mediated cytotoxicity | |
| hsa04380 | Osteoclast differentiation | |
| hsa04611 | Platelet activation | |
| hsa04015 | Rap1 signaling pathway | |
| hsa04660 | T cell receptor signaling pathway | |
| hsa05135 | Yersinia infection | |
| MYH14 | hsa05130 | Pathogenic Escherichia coli infection |
| hsa04810 | Regulation of actin cytoskeleton | |
| hsa04530 | Tight junction | |
| hsa04270 | Vascular smooth muscle contraction | |
| PDSS2 | hsa00900 | Terpenoid backbone biosynthesis |
| PHKA1 | hsa04(020,922,910) | signaling pathway (Calcium, Glucagon, Insulin) |
| PPP2R5C | hsa04261 | Adrenergic signaling in cardiomyocytes |
| hsa04152 | AMPK signaling pathway | |
| hsa04728 | Dopaminergic synapse | |
| hsa05165 | Human papillomavirus infection | |
| hsa03015 | mRNA surveillance pathway | |
| hsa04114 | Oocyte meiosis | |
| hsa04151 | PI3K-Akt signaling pathway | |
| hsa04071 | Sphingolipid signaling pathway | |
| SLC1A3 | hsa04724 | Glutamatergic synapse |
| hsa05016 | Huntington disease | |
| hsa04721 | Synaptic vesicle cycle | |
| SLC36A4 | hsa04974 | Protein digestion and absorption |
| ST8SIA1 | hsa00604 | Glycosphingolipid biosynthesis |
| hsa01100 | Metabolic pathways | |
| UBE2K | hsa04120 | Ubiquitin mediated proteolysis |
| Targeted Gene | KEGG Id | Pathway |
|---|---|---|
| RB1 | bta01522, bta04110, bta0421 bta04934, bta05160, bta0516, bta05200 | Endocrine resistance, Cell cycle, Cellular senescence, Cushing syndrome, Hepatitis B, Hepatitis C, Pathways in cancer |
| SCNN1B | bta04742, bta04960 | Taste transduction, Aldosterone-regulated sodium reabsorption |
| CFLAR | bta04064, bta04140, bta04210 bta04217, bta04668, bta05142, bta05160 | NF-kappa B signaling pathway, Autophagy—animal, Apoptosis, Necroptosis, TNF signaling pathway, Chagas disease, Hepatitis C |
| COL4A4 | bta04933, bta05146, bta04512, bta04510, bta05200, bta04151, bta04974, bta04926 | AGE-RAGE signaling pathway in diabetic complications, Amoebiasis, ECM-receptor interaction, Focal adhesion, Pathways in cancer, PI3K-Akt signaling pathway Protein digestion and absorption, Relaxin signaling pathway, |
| TRPA1 | bta04750 | Inflammatory mediator regulation of TRP channels |
| FAT4 | bta04392 | Hippo signaling pathway—multiple species |
| ITGB7 | bta05412, bta04514, bta05414, bta05410, bta04672, bta04810, bta05202 | Arrhythmogenic right ventricular cardiomyopathy, Cell adhesion molecules, Dilated cardiomyopathy, Hypertrophic cardiomyopathy, Intestinal immune network for IgA production, Regulation of actin cytoskeleton, Transcriptional misregulation in cancer |
| PIP5K1A | bta05231, bta04144, bta04666, bta00562, bta01100, bta04070, bta04072, bta05135 | Choline metabolism in cancer, Endocytosis, Fc gama R-mediated phagocytosis, Inositol phosphate metabolism, Metabolic pathways, Phosphatidylinositol signaling system, Phospholipase D signaling pathway, Yersinia infection |
| LOC533983 | bta04740, bta04975, bta00561 | Olfactory transduction, Fat digestion and absorption, Glycerolipid metabolism |
| cb-miR Id | Targeted Gene | Association Type | Disease | References |
|---|---|---|---|---|
| Ct-miR-3037 | DAZAP2 | Posttranslational Modification | Multiple myeloma | [40] |
| Ct-miR-3169 | KLHL20 | Biomarker | Alzheimer’s disease | [41] |
| Cte-miR824-3p | TNS1 | Genetic Variation | Malignant tumor of breast | [42] |
| Ct-miR-3135 | BBIP1 | Biomarker | Bardet-Biedl syndrome 18 | [43] |
| Cte-miR8741 | HMGCS2 | Biomarker | 3-hydroxy-3-methylglutaryl-CoA synthase deficiency | [44] |
| Cte-miR7780-3p | PAPD4 | Biomarker | Hepatitis C virus infection | [45] |
| Ct-miR-3069 | FAM212B | Genetic Variation | Crohn’s disease | [46] |
| Cte-miR8577 | KIAA1549 | Genetic Variation | Pilocytic astrocytoma | [47] |
| Cb-miR Id | Targeted Gene | Association Type | Disease | References |
|---|---|---|---|---|
| Ct-miR-3034, 3061 | DNTT, UHMK1 | Biomarker | Schizophrenia | [48] |
| Ct-miR-3095 | PTPRE | Biomarker | Asthma | [49] |
| Cte-miR5084 cte-miR04 | CNRIP1, SLC36A4 | Biomarker | Colorectal Carcinoma | [50] |
| Ct-miR-3035, 3094 Cte-miR8713 | CLEC4G, LCP2, ST8SIA1 | Biomarker | Liver carcinoma | [51] |
| Cte-miR5644, 824-3p | NLRC5, TNS1 | Altered Expression | Malignant neoplasm | [52] |
| Ct-miR-3007 | NRM | Genetic Variation | Rheumatoid Arthritis | [53] |
| Cte-miR8577 | KIAA1549 | Genetic Variation | Pilocytic Astrocytoma | [47] |
| Cte-miR1134 | LMTK2 | Biomarker | Malignant neoplasm of prostate | [54] |
| Cte-miR531 | SLC6A6 | Biomarker | Myocardial Ischemia | [55] |
| Ct-miR-3104 | TYW1 | Genetic Variation | Lymphocyte Count measurement | [56] |
| Cte-miR117 | SUZ12 | Genetic Variation | Endometrial Stromal Sarcoma | [57] |
| Ct-miR-3096 | SLC1A3 | Causal Mutation | Episodic Ataxia, TYPE 6 | [58] |
| Ct-miR-3041 | PHKA1 | Genetic Variation | Glycogen Storage Disease, Type IXD | [59] |
| Ct-miR-3027 | SHOX | Genetic Variation | Leri-Weill dyschondrosteosis | [60] |
| Ct-miR-3015 | PPP2R5C | Biomarker | Neoplasm | [61] |
| Ct-miR-3139 | PDSS2 | Causal Mutation | Coenzyme Q10 Deficiency | [44] |
| Cte-miR168 | UBE2K | Genetic Variation | Angelman Syndrome | [62] |
| Cte-miR8741 | HMGCS2 | Genetic Variation | 3-Hydroxy-3-Methylglutaryl-CoA Synthase 2 Deficiency | [44] |
| Ct-miR-3097 | GABRA6 | Biomarker | Alcoholic Intoxication, Chronic | [63] |
| Ct-miR-3169 | KLHL20 | Biomarker | Alzheimer’s Disease | [41] |
| Cb-miR Id | Gene | Description | References |
|---|---|---|---|
| Cte-miR5084 | PLIN3 | Associated with various metabolic diseases, primarily expressed in adipose tissue. | [64] |
| EDEM2 | Mutations leads genetic disorder, bovine osteoporosis. | [65] | |
| Cte-miR531 | ECM1 | Mutations leads genetic disorder, bovine hereditary angioneurotic edema (HANE). | [66] |
| Ct-miR-3061 | SURF4 | In relation to milk production traits and mastitis resistance. | [67] |
| SEC14L5 | Variations/mutation leads susceptibility to mastitis, lead to reduced milk production and quality and involved in regulating immune function in response to bacterial infection. | [68] | |
| Ct-miR-3069 | DHRS11 | Variations/mutation lead susceptibility to infectious diseases like bovine viral diarrhea virus in cattle. | [69] |
| Ct-miR-3135 | LGALS9 | Expressed in bovine respiratory disease (BRD, and may be involved in the development and progression of BRD. | [70] |
| ALDH18A1 | Mutations lead Hyperprolinemia type II, rare autosomal recessive disorder. | [71] | |
| SPTAN1 | Mutations leads Cerebellar abiotrophy, neurological disorder that affects the cerebellum. | [72] | |
| NXPE4 | SNP (single nucleotide polymorphism) in the NXPE4 gene was significantly associated with milk yield and fat content in milk. | [73] | |
| FIZ1 | Variations lead BRD is a multifactorial disease. | [74] | |
| Ct-miR3033 | GIPC3 | Potential role in cell growth, differentiation and survival. | [75] |
| cb-miR Id | Gene | Description | References |
|---|---|---|---|
| Cte-miR824-3p | CFLAR | Malignant glioma, cancer | [76] |
| Cte-miR5644 | TRPA1 | Respiratory diseases | [77] |
| Ct-miR-3017 | RB1 | Mutations lead sporadic cases of retinoblastoma | [78] |
| Ct-miR-3145 | SCNN1B | Dysfunctions in SCNN1B renal diseases | [79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahu, S.; Rao, A.R.; Sahu, T.K.; Pandey, J.; Varshney, S.; Kumar, A.; Gaikwad, K. Predictive Role of Cluster Bean (Cyamopsis tetragonoloba) Derived miRNAs in Human and Cattle Health. Genes 2024, 15, 448. https://doi.org/10.3390/genes15040448
Sahu S, Rao AR, Sahu TK, Pandey J, Varshney S, Kumar A, Gaikwad K. Predictive Role of Cluster Bean (Cyamopsis tetragonoloba) Derived miRNAs in Human and Cattle Health. Genes. 2024; 15(4):448. https://doi.org/10.3390/genes15040448
Chicago/Turabian StyleSahu, Sarika, Atmakuri Ramakrishna Rao, Tanmaya Kumar Sahu, Jaya Pandey, Shivangi Varshney, Archna Kumar, and Kishor Gaikwad. 2024. "Predictive Role of Cluster Bean (Cyamopsis tetragonoloba) Derived miRNAs in Human and Cattle Health" Genes 15, no. 4: 448. https://doi.org/10.3390/genes15040448
APA StyleSahu, S., Rao, A. R., Sahu, T. K., Pandey, J., Varshney, S., Kumar, A., & Gaikwad, K. (2024). Predictive Role of Cluster Bean (Cyamopsis tetragonoloba) Derived miRNAs in Human and Cattle Health. Genes, 15(4), 448. https://doi.org/10.3390/genes15040448