
Transcriptome Analysis on the Quality of Epimedium koreanum in Different Soil Moisture Conditions at Harvesting Stage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Sample Treatment
2.3. Instruments
2.4. Experimental Methods
2.4.1. Determination of Epimedin A, Epimedin B, Epimedin C, and Icariin Content
2.4.2. Determination of MDA and Antioxidant Enzyme Levels
2.4.3. Total RNA Extraction and Transcriptome Sequencing
2.4.4. Differentially Expressed Genes, Enrichment, and Interaction Analysis
2.4.5. qRT-PCR Validation
3. Results
3.1. Content of Active Ingredients
3.2. MDA and Protective Enzyme Content
3.3. Sequencing Quality of E. koreanum RNA
3.4. Analysis of Differentially Expressed Genes
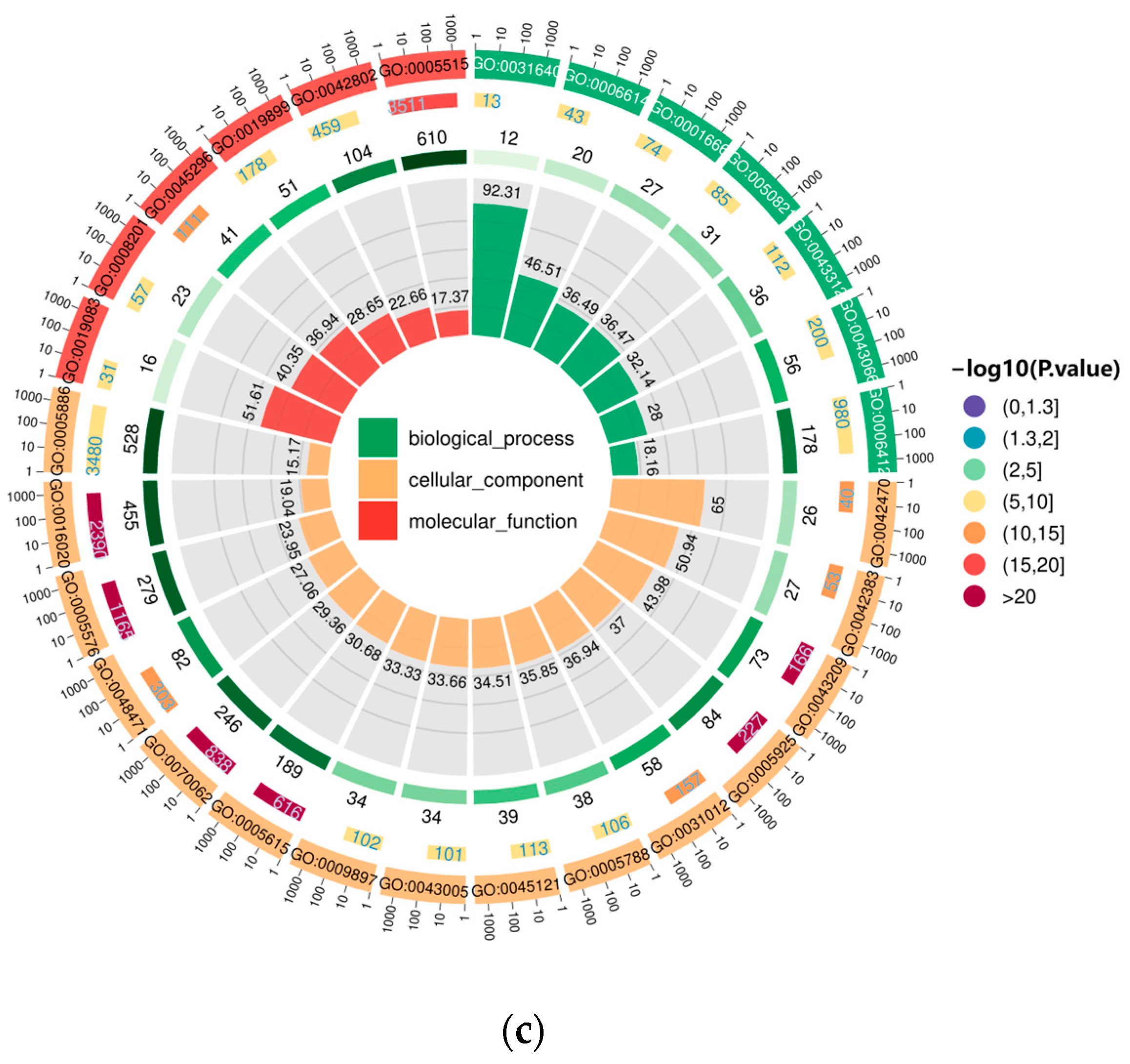
3.5. GO Enrichment Analysis
3.6. KEGG Enrichment Analysis
3.7. Gene Expression Analysis and Key Gene Screening of Flavonoid Synthesis Pathway in E. koreanum
3.8. Correlation Analysis between the Quality of E. koreanum and Influencing Factors
3.9. Analysis of Protein Interactions and Prediction of Key Proteins in Flavonoid-Synthesizing Genes in E. koreanum
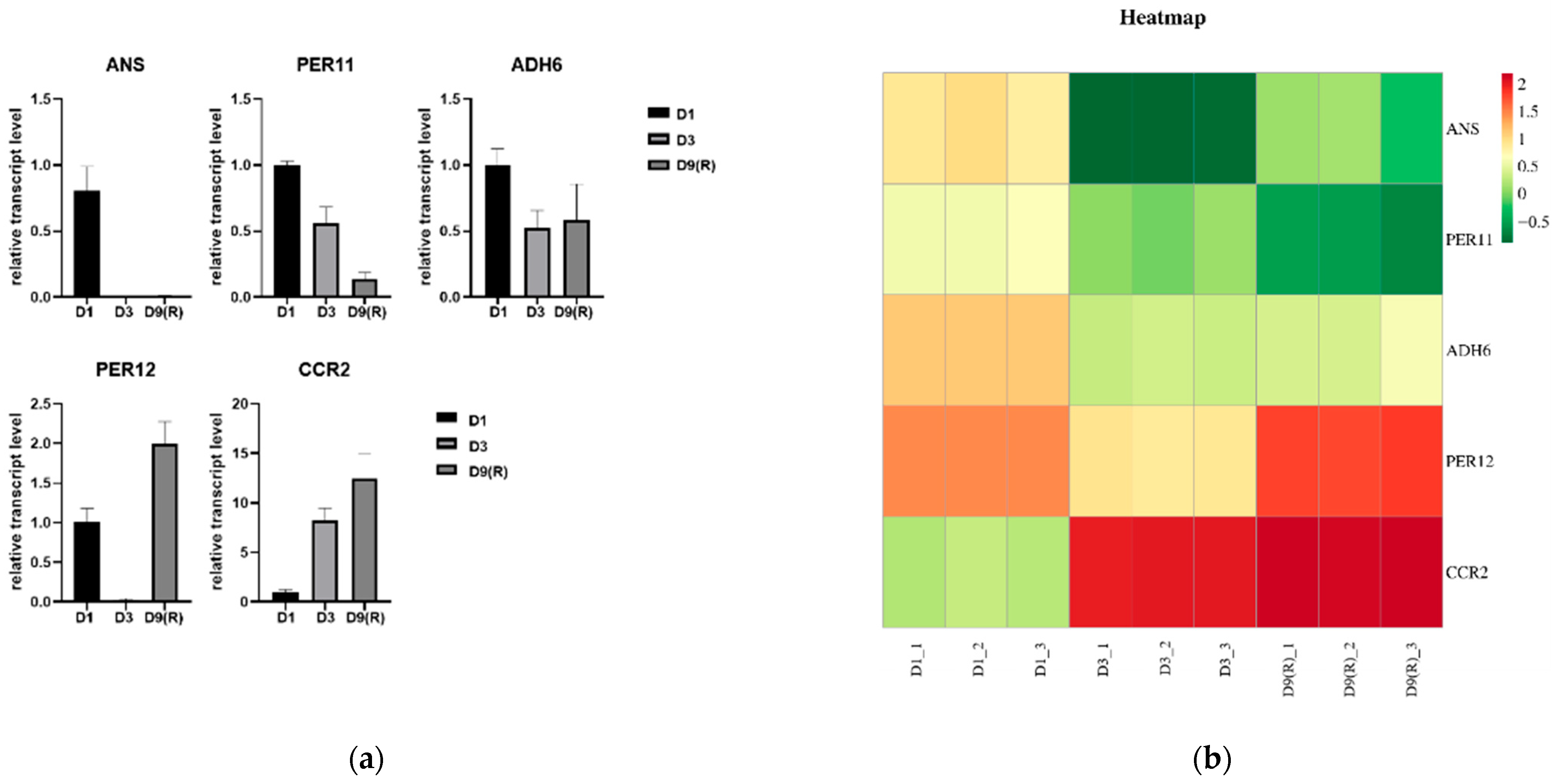
3.10. qRT-PCR Validation of RNA-seq Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Editoria Committee Flora of China. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 2001; Volume 29, pp. 278–281. ISBN 9787030072733. [Google Scholar]
- Ying, J.S. Petal evolution and geographical distribution Pattern of Epimedium (Berberidaceae). Acta Bot. Sin. 2002, 6, 481–489. [Google Scholar]
- Xu, Y.Q.; Jiang, Y.; Huang, H. Taxonomic studies of Epimedium: Progress, problems and prospects. Guangxi Bot. 2020, 40, 601–607. [Google Scholar]
- Yang, L.M. Plant Resources Science; Agriculture Press: Beijing, China, 2008; p. 104. [Google Scholar]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; Volume 1, pp. 340–342. [Google Scholar]
- Ma, H.; He, X.; Yang, Y.; Li, M.; Hao, D.; Jia, Z. The genus Epimedium: An ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2011, 134, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Jung, H.A.; Sohn, H.S.; Kim, J.W.; Choi, J.S. Potential of Icariin Metabolites from Epimedium koreanum Nakai as Antidiabetic Therapeutic Agents. Molecules 2017, 22, 986. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.L.; Tan, J.J.; Xiong, Y.G.; Li, C.; Yang, G.; Zhang, H. Progress of dehydrated Icariin in Epimediu. Chin. Mod. Tradit. Med. 2023, 25, 2636–2642. [Google Scholar]
- Xing, Z.F.; Yu, H.; Wan, X.H. Research progress on extraction, purification, structural characteristics and biological activity of Epimedium polysaccharides. Pharm. Res. 2023, 42, 830–836. [Google Scholar]
- Kim, E.; Kim, Y.-M.; Ahn, J.; Chae, H.-S.; Chin, Y.-W.; Kim, J. Prenylated Flavonoid Glycosides with PCSK9 mRNA Expression Inhibitory Activity from the Aerial Parts of Epimedium koreanum. Molecules 2021, 26, 3590. [Google Scholar] [CrossRef]
- Long, Y.; Tao, H.; Wang, S.; Xing, B.; Wang, Z.; Liu, K.; Shao, Q.; Gao, F. Identification and Functional Validation of Two Novel Antioxidant Peptides in Saffron. Antioxidants 2024, 13, 378. [Google Scholar] [CrossRef]
- Li, Q.; Gu, L.; Song, J.; Li, C.; Zhang, Y.; Wang, Y.; Pang, Y.; Zhang, B. Physiological and transcriptome analyses highlight multiple pathways involved in drought stress in Medicago falcata. PLoS ONE 2022, 17, e0266542. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, L.B.d.S.; Tattini, M. Beyond Photoprotection: The Multifarious Roles of Flavonoids in Plant Terrestrialization. Int. J. Mol. Sci. 2022, 23, 5284. [Google Scholar] [CrossRef] [PubMed]
- Um, T.; Choi, J.; Park, T.; Chung, P.J.; Jung, S.E.; Shim, J.S.; Kim, Y.S.; Choi, I.Y.; Park, S.C.; Oh, S.J.; et al. Rice microRNA171f/SCL6 module enhances drought tolerance by regulation of flavonoid biosynthesis genes. Plant Direct. 2022, 6, e374. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, M.; Cheng, L.; Yang, L.; Han, M. Changes in the Platycodin Content and Physiological Characteristics during the Fruiting Stage of Platycodon grandiflorum under Drought Stress. Sustainability 2022, 14, 6285. [Google Scholar] [CrossRef]
- Liu, H.Y.; Wang, X.D.; Wang, D.H.; Zou, Z.R.; Liang, Z.S. Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind. Crops Prod. 2011, 33, 84–88. [Google Scholar] [CrossRef]
- Wu, C.; Liu, B.; Zhang, X.; Wang, M.; Liang, H. Phytohormone Response of Drought-Acclimated Illicium difengpi (Schisandraceae). Int. J. Mol. Sci. 2023, 24, 16443. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Hu, Z.-M.; Song, W.; Wang, Z.-L.; He, J.-B.; Shi, X.-M.; Cui, Q.-H.; Qiao, X.; Ye, M. Diversity of O-Glycosyltransferases Contributes to the Biosynthesis of Flavonoid and Triterpenoid Glycosides in Glycyrrhiza uralensis. ACS Synth. Biol. 2019, 8, 1858–1866. [Google Scholar] [CrossRef]
- Shomali, A.; Das, S.; Arif, N.; Sarraf, M.; Zahra, N.; Yadav, V.; Aliniaeifard, S.; Chauhan, D.K.; Hasanuzzaman, M. Diverse Physiological Roles of Flavonoids in Plant Environmental Stress Responses and Tolerance. Plants 2022, 11, 3158. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, Y.; Lv, X.; Li, N.; Li, X.; Xing, J.; Li, C.; Hu, B. Abiotic factors and endophytes co-regulate flavone and terpenoid glycoside metabolism in Glycyrrhiza uralensis. Appl. Microbiol. Biotechnol. 2023, 107, 2671–2688. [Google Scholar] [CrossRef]
- Liang, Y.; Wei, K.; Wei, F.; Qin, S.; Deng, C.; Lin, Y.; Li, M.; Gu, L.; Wei, G.; Miao, J.; et al. Integrated transcriptome and small RNA sequencing analyses reveal a drought stress response network in Sophora tonkinensis. BMC Plant Biol. 2021, 21, 566. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Wang, Y.; Ding, Z.; Zhao, L. Global Transcriptional Analysis Reveals the Complex Relationship between Tea Quality, Leaf Senescence and the Responses to Cold-Drought Combined Stress in Camellia sinensis. Front. Plant Sci. 2016, 7, 1858. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wei, W.; Pang, X.; Wang, X.; Zhang, H.; Dong, B.; Xing, Y.; Li, X.; Wang, M. Comparative transcriptome profiling of a desert evergreen shrub, Ammopiptanthus mongolicus, in response to drought and cold stresses. BMC Genom. 2014, 15, 671. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; Li, P.; Wang, T.; Zhang, F.-J.; Huang, X.-X.; Hou, B.-K. The maize secondary metabolism glycosyltransferase UFGT2 modifies flavonols and contributes to plant acclimation to abiotic stresses. Ann. Bot. 2018, 122, 1203–1217. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Chen, X.R.; Wang, J.P.; Cui, W.Q.; Xing, X.X.; Chen, X.Y.; Ding, W.Y.; God’spower, B.O.; Eliphaz, N.; Sun, M.Q.; et al. Transcriptomic analysis reveals flavonoid biosynthesis of Syringa oblata Lindl. in response to different light intensity. BMC Plant Biol. 2019, 19, 487. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Gradient Phase A (%) | Gradient Phase C (%) |
|---|---|---|
| 0~20 | 24→26 | 76→74 |
| 20~30 | 26→30 | 74→70 |
| 30~45 | 30→45 | 70→55 |
| 45~50 | 45→60 | 55→40 |
| 50~60 | 60→24 | 40→76 |
| Epimedin A | Epimedin B | Epimedin C | Icariin |
|---|---|---|---|
 |  |  |  |
| Name | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|
| 18S | CGCTGCGTTCTTCATCGTT | TTGGGTAGTCGGGCTGGTC |
| ANS | ACCAAGGCAGCATTGTTAGC | TGAACAAGGCGGGTAGTAACC |
| PER11 | AGCATTGGTTCCCATGCAAC | ACAGCACACTCCATCTCCTTC |
| ADH6 | TCACTTCGGCTTGCTATTCG | TCGTCAGTGGCGATGAAATG |
| OMT | ATTCCGTCATCACTGCCTCTAG | ACTTGCAAACAGGTGCCAAG |
| CCR2 | TGTCCTAAAGGCATGCGTTG | AAACAGAACAGCGGCAATGG |
| Sample | Raw Reads | Raw Bases | Valid Reads | Valid Bases | Valid% | Q20% | GC% |
|---|---|---|---|---|---|---|---|
| D1-1 | 43,154,254 | 6.47 G | 42,501,058 | 6.31 G | 98.49 | 98.06 | 45.29 |
| D1-2 | 47,879,254 | 7.18 G | 47,119,460 | 6.99 G | 98.41 | 97.99 | 45.03 |
| D1-3 | 48,121,936 | 7.22 G | 47,358,136 | 7.02 G | 98.41 | 98.05 | 45.30 |
| D3-1 | 40,412,608 | 6.06 G | 39,812,314 | 5.91 G | 98.51 | 98.18 | 44.91 |
| D3-2 | 39,276,616 | 5.89 G | 38,687,458 | 5.74 G | 98.50 | 98.22 | 44.91 |
| D3-3 | 39,017,882 | 5.85 G | 38,438,222 | 5.71 G | 98.51 | 98.18 | 44.93 |
| D9(R)-1 | 45,177,958 | 6.78 G | 44,464,904 | 6.60 G | 98.42 | 98.1 | 44.83 |
| D9(R)-2 | 49,464,974 | 7.42 G | 48,634,656 | 7.21 G | 98.32 | 98.09 | 45.12 |
| D9(R)-3 | 43,015,912 | 6.45 G | 42,379,574 | 6.29 G | 98.52 | 98.11 | 44.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, D.; Wu, F.; Huang, X.; Chai, X.; Yang, L. Transcriptome Analysis on the Quality of Epimedium koreanum in Different Soil Moisture Conditions at Harvesting Stage. Genes 2024, 15, 528. https://doi.org/10.3390/genes15050528
Zhang Y, Wang D, Wu F, Huang X, Chai X, Yang L. Transcriptome Analysis on the Quality of Epimedium koreanum in Different Soil Moisture Conditions at Harvesting Stage. Genes. 2024; 15(5):528. https://doi.org/10.3390/genes15050528
Chicago/Turabian StyleZhang, Yonggang, Dantong Wang, Feng Wu, Xiangdi Huang, Xiaowei Chai, and Limin Yang. 2024. "Transcriptome Analysis on the Quality of Epimedium koreanum in Different Soil Moisture Conditions at Harvesting Stage" Genes 15, no. 5: 528. https://doi.org/10.3390/genes15050528
APA StyleZhang, Y., Wang, D., Wu, F., Huang, X., Chai, X., & Yang, L. (2024). Transcriptome Analysis on the Quality of Epimedium koreanum in Different Soil Moisture Conditions at Harvesting Stage. Genes, 15(5), 528. https://doi.org/10.3390/genes15050528