
Presence of a Prophage Determines Temperature-Dependent Capsule Production in Streptococcus pyogenes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Quantitation of S. pyogenes Capsule
2.3. Targeted Insertional Disruption of the L897_07695 Gene
2.4. Passaging Cells for Generating Spontaneous Capsule Thermoregulation Mutants
2.5. Whole Genome Sequencing
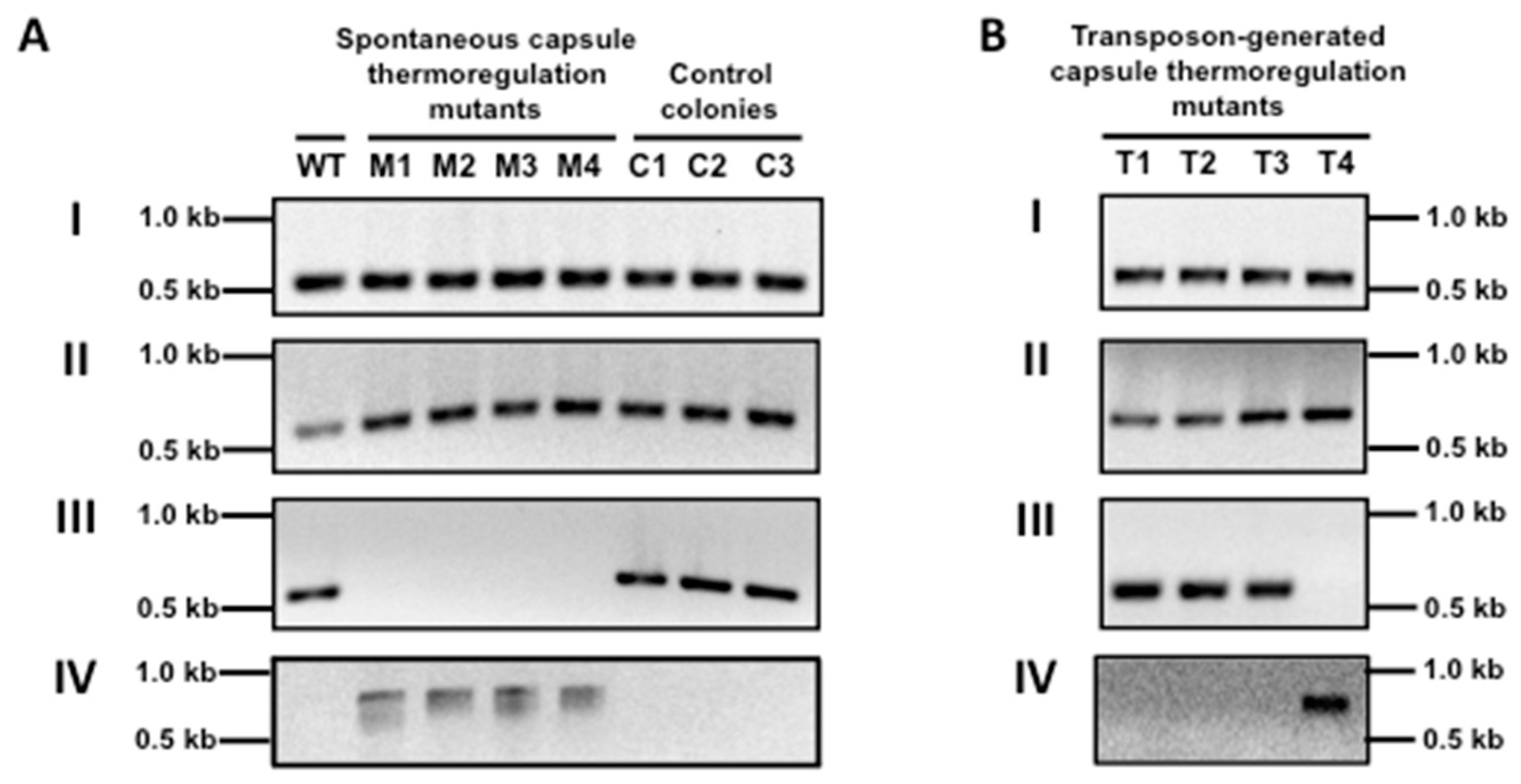
2.6. PCR to Determine the Curing of Prophages
2.7. Phage Nomenclature
3. Results
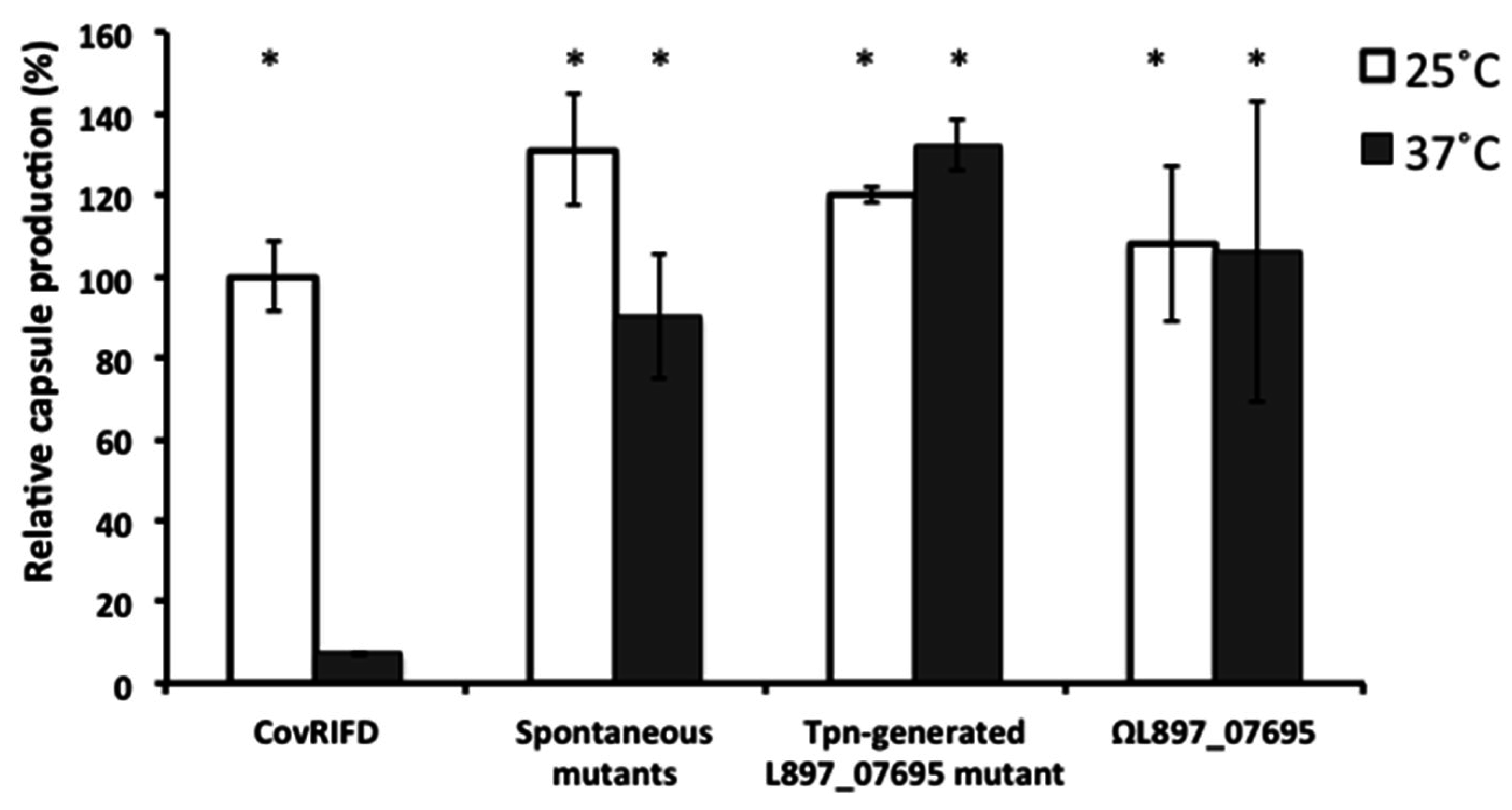
3.1. Generation of Capsule Thermoregulation Mutants by in Vitro Passaging
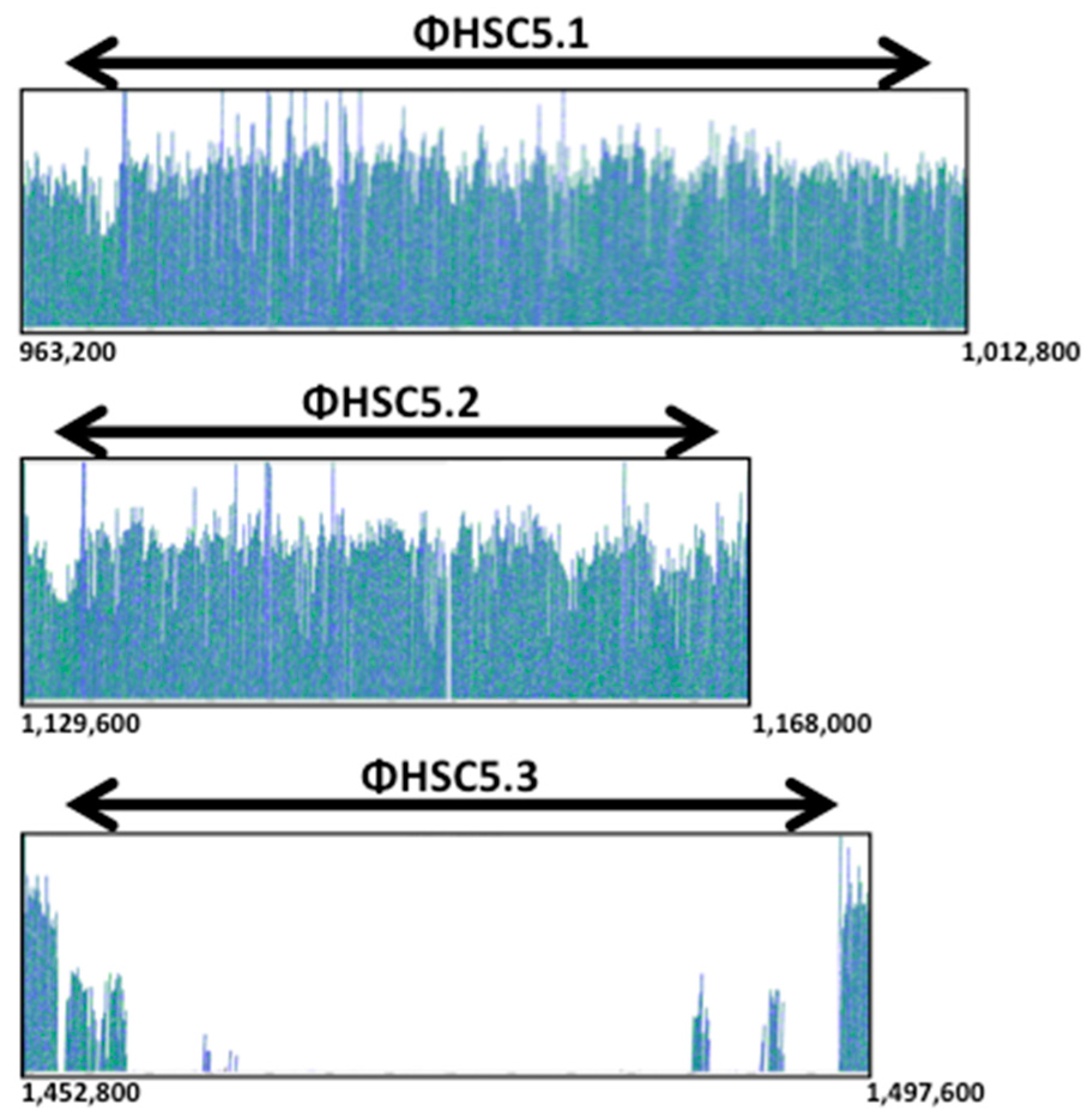
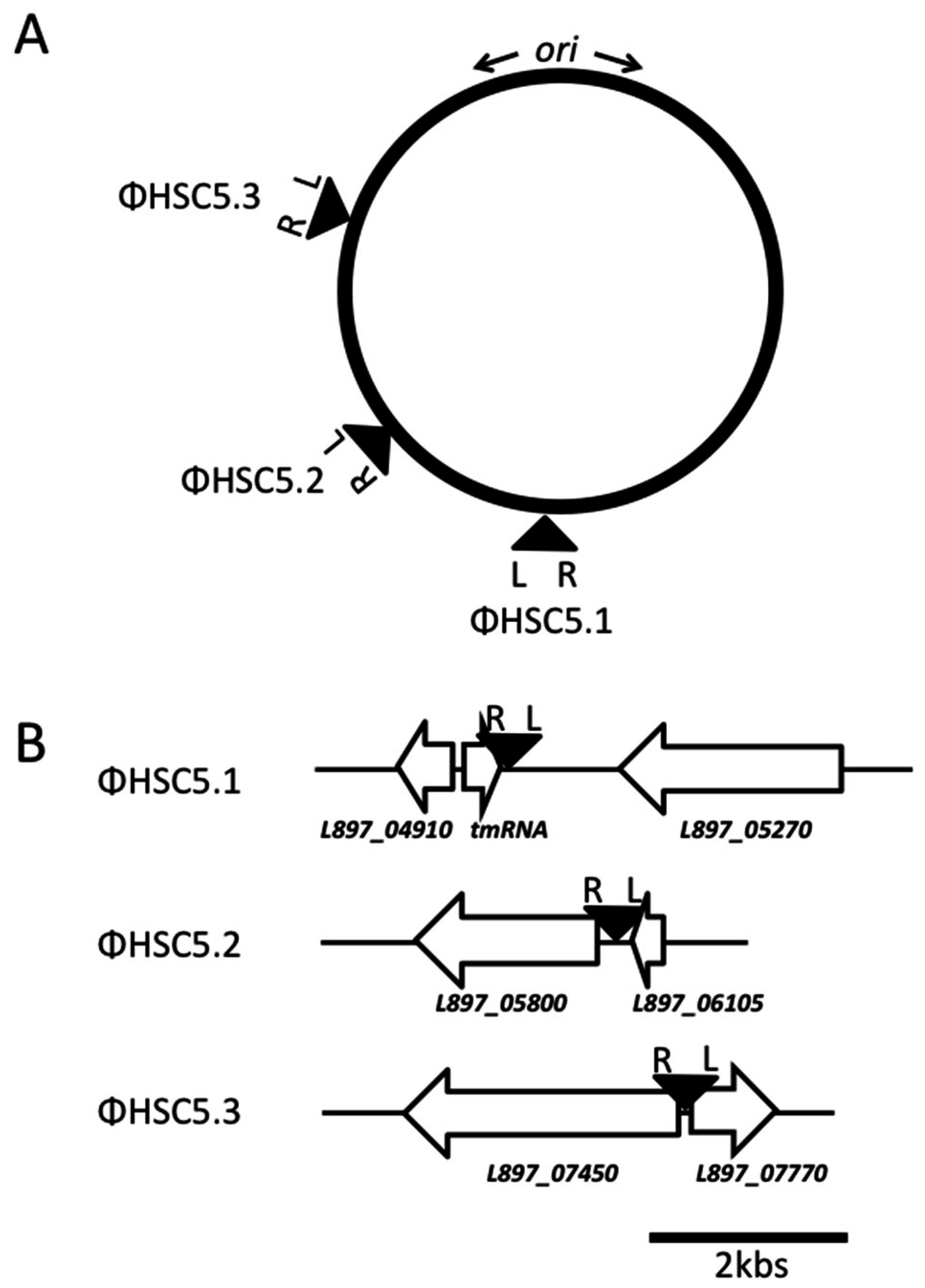
3.2. A Prophage Was Cured in Spontaneous Capsule Thermoregulation-Negative Mutants
3.3. The Prophage ΦHSC5.3 Influences Capsule Thermoregulation
3.4. The Role of CvfA in Capsule Thermoregulation is Independent of the Curing of ΦHSC5.3
3.5. Disruption of the L897_07695 Gene in ΦHSC5.3 Confers the Same Capsule Thermoregulation-Negative Phenotype as That of the ΦHSC5.3-Cured Strains
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CFU | Colony Forming Unit |
| IGR | Intergenic Region |
| IFD | In-Frame Deletion |
| OD | Optical Density |
| PBS | Phosphate-Buffered Saline |
| PCR | Polymerase Chain Reaction |
| WT | Wild Type |
References
- Kass, E.H.; Seastone, C.V. The role of the mucoid polysaccharide (hyaluronic acid) in the virulence of group a hemolytic streptococci. J. Exp. Med. 1944, 79, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Dinkla, K.; Sastalla, I.; Godehardt, A.W.; Janze, N.; Chhatwal, G.S.; Rohde, M.; Medina, E. Upregulation of capsule enables Streptococcus pyogenes to evade immune recognition by antigen-specific antibodies directed to the G-related α2-macroglobulin-binding protein GRAB located on the bacterial surface. Microbes Infect. 2007, 9, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Wessels, M.R.; Goldberg, J.B.; Moses, A.E.; DiCesare, T.J. Effects on virulence of mutations in a locus essential for hyaluronic acid capsule expression in group a streptococci. Infect. Immun. 1994, 62, 433–441. [Google Scholar] [PubMed]
- Schrager, H.M.; Alberti, S.; Cywes, C.; Dougherty, G.J.; Wessels, M.R. Hyaluronic acid capsule modulates M protein-mediated adherence and acts as a ligand for attachment of group a streptococcus to CD44 on human keratinocytes. J. Clin. Investig. 1998, 101, 1708–1716. [Google Scholar] [CrossRef] [PubMed]
- Schrager, H.M.; Rheinwald, J.G.; Wessels, M.R. Hyaluronic acid capsule and the role of streptococcal entry into keratinocytes in invasive skin infection. J. Clin. Investig. 1996, 98, 1954–1958. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Kawabata, S.; Terao, Y.; Fujitaka, H.; Okuno, Y.; Hamada, S. The Streptococcus pyogenes capsule is required for adhesion of bacteria to virus-infected alveolar epithelial cells and lethal bacterial-viral superinfection. Infect. Immun. 2004, 72, 6068–6075. [Google Scholar] [CrossRef] [PubMed]
- Levin, J.C.; Wessels, M.R. Identification of csrR/csrS, a genetic locus that regulates hyaluronic acid capsule synthesis in group a streptococcus. Mol. Microbiol. 1998, 30, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Federle, M.J.; Scott, J.R. Identification of binding sites for the group a streptococcal global regulator covr. Mol. Microbiol. 2002, 43, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Federle, M.J.; McIver, K.S.; Scott, J.R. A response regulator that represses transcription of several virulence operons in the group a streptococcus. J. Bacteriol. 1999, 181, 3649–3657. [Google Scholar] [PubMed]
- Kang, S.O.; Wright, J.O.; Tesorero, R.A.; Lee, H.; Beall, B.; Cho, K.H. Thermoregulation of capsule production by Streptococcus pyogenes. PLoS ONE 2012, 7, e37367. [Google Scholar] [CrossRef] [PubMed]
- Hanski, E.; Caparon, M. Protein F, a fibronectin-binding protein, is an adhesin of the group a streptococcus Streptococcus pyogenes. Proc. Natl. Acad. Sci. USA 1992, 89, 6172–6176. [Google Scholar] [CrossRef] [PubMed]
- Hanski, E.; Horwitz, P.A.; Caparon, M.G. Expression of protein F, the fibronectin-binding protein of Streptococcus pyogenes JRS4, in heterologous streptococcal and enterococcal strains promotes their adherence to respiratory epithelial cells. Infect. Immun. 1992, 60, 5119–5125. [Google Scholar] [PubMed]
- Okada, N.; Geist, R.T.; Caparon, M.G. Positive transcriptional control of mry regulates virulence in the group A streptococcus. Mol. Microbiol. 1993, 7, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Caparon, M.G.; Scott, J.R. Genetic manipulation of pathogenic streptococci. Meth. Enzymol. 1991, 204, 556–586. [Google Scholar] [PubMed]
- Banks, D.J.; Beres, S.B.; Musser, J.M. The fundamental contribution of phages to gas evolution, genome diversification and strain emergence. Trends Microbiol. 2002, 10, 515–521. [Google Scholar] [CrossRef]
- Canchaya, C.; Proux, C.; Fournous, G.; Bruttin, A.; Brussow, H. Prophage genomics. Microbiol. Mol. Biol. Rev. 2003, 67, 238–276. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.O.; Caparon, M.G.; Cho, K.H. Virulence gene regulation by Cvfa, a putative RNase: The Cvfa-enolase complex in streptococcus pyogenes links nutritional stress, growth-phase control, and virulence gene expression. Infect. Immun. 2010, 78, 2754–2767. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H.; Canchaya, C.; Hardt, W.D. Phages and the evolution of bacterial pathogens: From genomic rearrangements to lysogenic conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed]
- Spanier, J.G.; Cleary, P.P. Bacteriophage control of antiphagocytic determinants in group a streptococci. J. Exp. Med. 1980, 152, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H.; Desiere, F. Comparative phage genomics and the evolution of siphoviridae: Insights from dairy phages. Mol. Microbiol. 2001, 39, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Edwards, R.A.; Taylor, W.W.; Low, D.E.; McGeer, A.; Kotb, M. Mosaic prophages with horizontally acquired genes account for the emergence and diversification of the globally disseminated M1T1 clone of Streptococcus pyogenes. J. Bacteriol. 2005, 187, 3311–3318. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.M. Preferential orientation of natural lambdoid prophages and bacterial chromosome organization. Theor. Popul. Biol. 2002, 61, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Canchaya, C.; Fournous, G.; Brussow, H. The impact of prophages on bacterial chromosomes. Mol. Microbiol. 2004, 53, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Ikebe, T.; Ato, M.; Matsumura, T.; Hasegawa, H.; Sata, T.; Kobayashi, K.; Watanabe, H. Highly frequent mutations in negative regulators of multiple virulence genes in group a streptococcal toxic shock syndrome isolates. PLoS Pathog. 2010, 6, e1000832. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Genotype/Description | Reference/Source |
|---|---|---|
| Escherichia coli | ||
| TOP10 | mcrA Δ(mrr-hsdRMS-mcrBC) φ80lacZΔM15 ΔlacX74 nupG recA1 araD139 Δ(ara-leu)7697 galE15 galK16 rpsL(StrR) endA1 | Invitrogen |
| Streptococcus pyogenes | ||
| HSC5 | Wild-type, M14 serotype | [12] |
| CovRIFD | HSC5 strain with in-frame deleted covR. This strain has a deletion from the leucine codon (L39) to valine codon (V183) in the gene of CovR (228 amino acid long). This is the parental strain for most mutagenesis in this study. | [10] |
| ΩL897_07695 | Strain of CovRIFD with the disruption of the prophage gene L897_07695 | This study |
| CovRIFD:TnCvfA1 & CovRIFD:TnCvfA2 | Strain of CovRIFD with a transposon insertion into cvfA | [10] |
| CovRIFD:TnL897_07695 | Strain of CovRIFD with a transposon insertion into L897_07695 | [10] |
| CovRIFD:TnHasAupIGR | Strain of CovRIFD with a transposon insertion into the intergenic region immediate upstream of hasA | [10] |
| Name | Prophage | Forward | Reverse |
|---|---|---|---|
| A | ΦHSC5.1 | 5ΦHSC5.1 AGAAACAGGCGATGCCATAC | 3ΦHSC5.1 CATTCATTTCATCCGACAGC |
| B | ΦHSC5.2 | 5ΦHSC5.2 GTCCACCGCTAAATCGAGAC | 3ΦHSC5.2 ATTTCTCCACCGATTTCACG |
| C | ΦHSC5.3 | 5ΦHSC5.3 CAATTATGGGAGCGGCTATG | 3ΦHSC5.3 ATCAAATTCTGTCGCCCAAG |
| D | ΦHSC5.3 | 5ΦHSC5.3del TTGCCCTTCCGCATAAATAG | 3ΦHSC5.3del ATCAATGGTCCTCAAGGCAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, L.; Kim, J.-H.; Cho, K.H. Presence of a Prophage Determines Temperature-Dependent Capsule Production in Streptococcus pyogenes. Genes 2016, 7, 74. https://doi.org/10.3390/genes7100074
Brown L, Kim J-H, Cho KH. Presence of a Prophage Determines Temperature-Dependent Capsule Production in Streptococcus pyogenes. Genes. 2016; 7(10):74. https://doi.org/10.3390/genes7100074
Chicago/Turabian StyleBrown, Leslie, Jeong-Ho Kim, and Kyu Hong Cho. 2016. "Presence of a Prophage Determines Temperature-Dependent Capsule Production in Streptococcus pyogenes" Genes 7, no. 10: 74. https://doi.org/10.3390/genes7100074
APA StyleBrown, L., Kim, J. -H., & Cho, K. H. (2016). Presence of a Prophage Determines Temperature-Dependent Capsule Production in Streptococcus pyogenes. Genes, 7(10), 74. https://doi.org/10.3390/genes7100074