
Identification, Characterization and Expression of Methuselah-Like Genes in Dastarcus helophoroides (Coleoptera: Bothrideridae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Identification and Rapid Amplification of cDNA Ends
2.3. Sequence and Phylogenetic Analysis
2.4. Expression Analysis through RT-qPCR
2.5. Statistical Analysis
3. Results
3.1. Identification and Cloning of Putative Mth-Like Genes
3.2. Phylogenetic Analysis of Mth-Like Genes
3.3. Developmental Expression Profiles
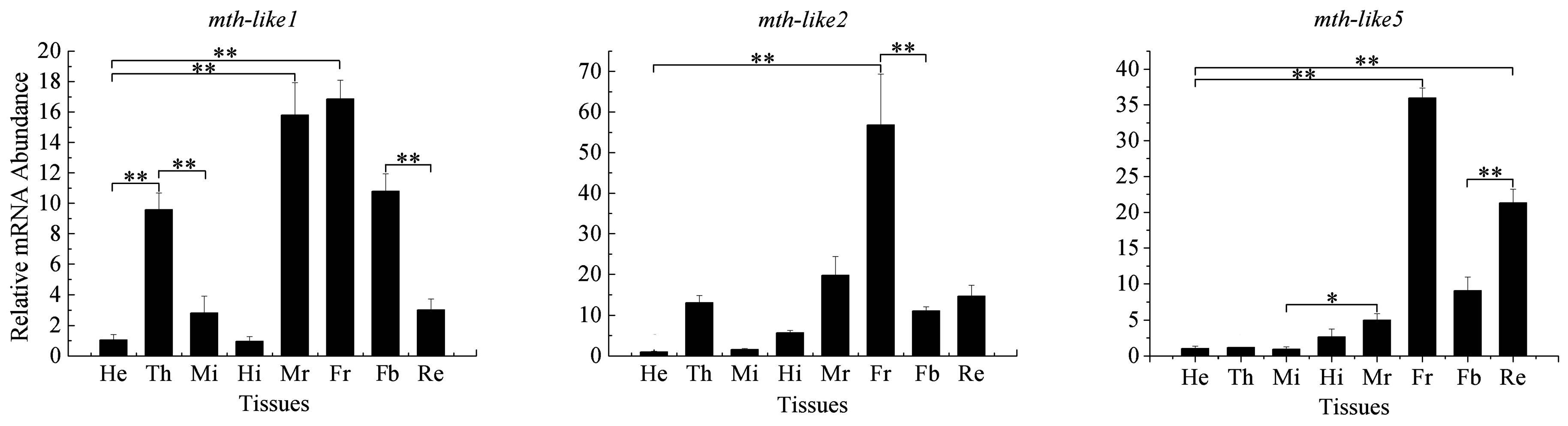
3.4. Tissue Expression Profiles
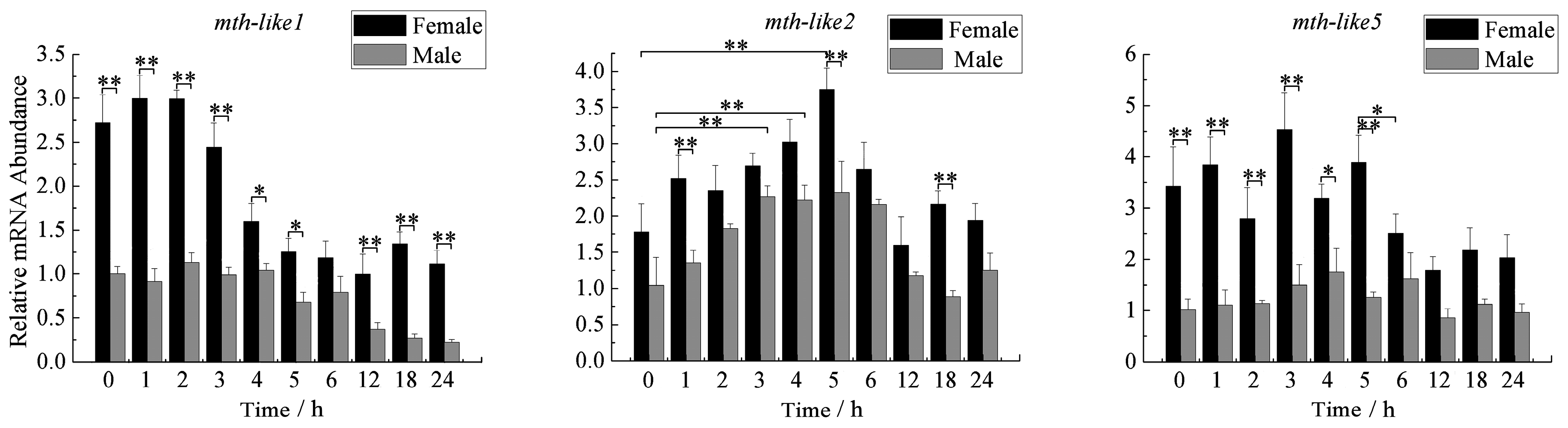
3.5. Expression Profiles during Aging Adults
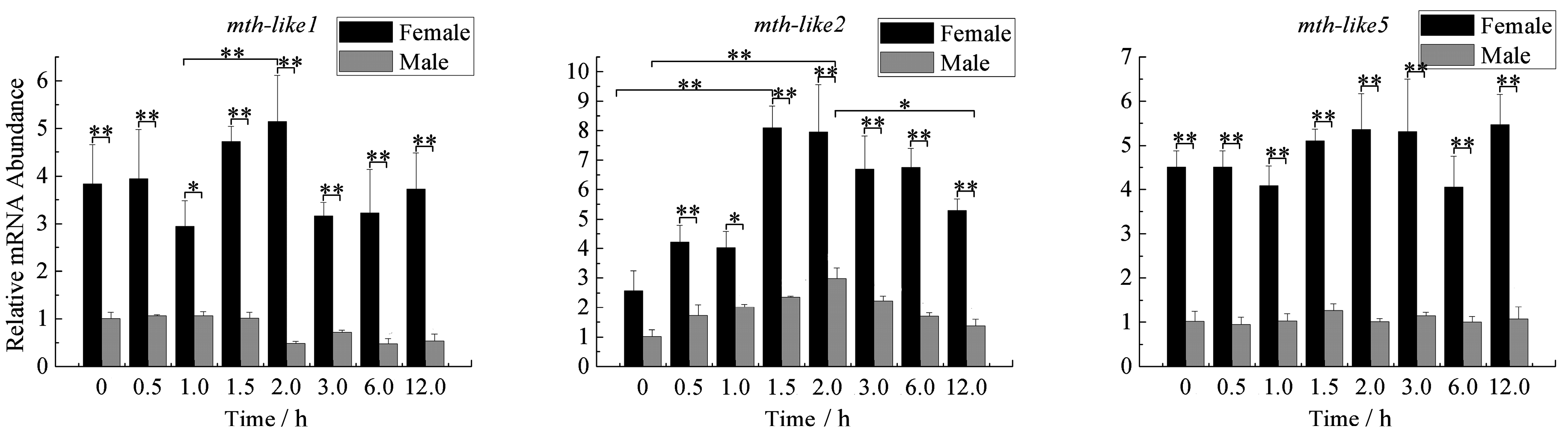
3.6. Expression Profiles under the Oxidative Stress
3.7. Expression Profiles under the Stress of High Temperature
3.8. Expression Profiles under the Stress of Starvation
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lin, Y.J.; Seroude, L.; Benzer, S. Extended life-span and stress resistance in the Drosophila mutant methuselah. Science 1998, 282, 943–946. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.V.; Hallal, D.A.; Jones, J.W.; Bronner, D.N.; Zein, R.; Caravas, J.; Husain, Z.; Friedrich, M.; Vanberkum, M.F.A. Dramatic expansion and developmental expression diversification of the methuselah gene family during recent Drosophila evolution. J. Exp. Zool. B Mol. Dev. Evol. 2012, 318, 368–387. [Google Scholar] [CrossRef] [PubMed]
- Brody, T.; Cravchik, A. Drosophila melanogaster G protein-coupled receptors. J. Cell Biol. 2000, 150, F83–F88. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Llamas, L.L.; Snow, P.M.; Benzer, S.; Bjorkman, P.J. Crystal structure of the ectodomain of Methuselah, a Drosophila G protein-coupled receptor associated with extended lifespan. Proc. Natl. Acad. Sci. USA 2001, 98, 3744–3749. [Google Scholar] [CrossRef] [PubMed]
- Harmar, A.J. Family-B G-protein-coupled receptors. Genome Biol. 2001, 2, 3013.1–3013.10. [Google Scholar] [CrossRef]
- Aigaki, T.; Seong, K.H.; Matsuo, T. Longevity determination genes in Drosophila melanogaster. Mech. Ageing Dev. 2002, 123, 1531–1541. [Google Scholar] [CrossRef]
- Grace, C.R.; Perrin, M.H.; Gulyas, J.; Digruccio, M.R.; Cantle, J.P.; Rivier, J.E.; Vale, W.W.; Riek, R. Structure of the N-terminal domain of a type B1 G protein-coupled receptor in complex with a peptide ligand. Proc. Natl. Acad. Sci. USA 2007, 104, 4858–4863. [Google Scholar] [CrossRef] [PubMed]
- Parthier, C.; Kleinschmidt, M.; Neumann, P.; Rudolph, R.; Manhart, S.; Schlenzig, D.; Fanghanel, J.; Rahfeld, J.U.; Demuth, H.U.; Stubbs, M.T. Crystal structure of the incretin-bound extracellular domain of a G protein-coupled receptor. Proc. Natl. Acad. Sci. USA 2007, 4, 13942–13947. [Google Scholar] [CrossRef] [PubMed]
- Sohal, R.S. Role of oxidative stress and protein oxidation in the aging process. Free Radic. Biol. Med. 2002, 33, 37–44. [Google Scholar] [CrossRef]
- Mockett, R.J.; Sohal, R.S. Temperature-dependent trade-offs between longevity and fertility in the Drosophila mutant, methuselah. Exp. Gerontol. 2006, 41, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Ranjan, R.; Dawson, S.K.; Bronk, P.; Marin, L.; Seroude, L.; Lin, Y.J.; Nie, Z.; Atwood, H.L.; Benzer, S.; et al. Presynaptic regulation of neurotransmission in Drosophila by the G protein-coupled receptor methuselah. Neuron 2002, 36, 105–119. [Google Scholar] [CrossRef]
- Lee, K.P.; Simpson, S.J.; Clissold, F.J.; Brooks, R.; Ballard, J.W.O.; Taylorp, W.; Soran, N.; Raubenheimer, D. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc. Natl. Acad. Sci. USA 2007, 7, 2498–2503. [Google Scholar] [CrossRef] [PubMed]
- Bjedov, I.; Toivonen, J.M.; Kerr, F.; Slack, C.; Jacobson, J.; Foley, A.; Partridge, L. Mechanisms of life span extension by Rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010, 11, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wallenfang, M.R.; Nayak, R.; Nardo, S.D. Dynamics of the male germline stem cell population during aging of Drosophila melanogaster. Aging Cell 2006, 5, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Petrosyan, A.; Hsieh, I.H.; Saberi, K. Age-dependent stability of sensorimotor functions in the life-extended Drosophila mutant Methuselah. Behav. Genet. 2007, 37, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G protein-coupled receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Sun, P.; Wang, Y.; He, X.B.; Deng, X.Y.; Chen, X.P.; Zhang, G.Z.; Chen, X.; Zhou, N.M. The G protein-coupled receptors in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Zhu, F.; Shah, K.; Palli, S.R. Large-scale RNAi screen of G protein-coupled receptors involved in larval growth, molting and metamorphosis in the red flour beetle. BMC Genom. 2011. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, M.; Sang, M.; Liu, X.; Wu, W.; Li, B. Comparative genomic analysis and evolution of family-B G protein-coupled receptors from six model insect species. Gene 2013, 519, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.R.; Yang, Z.Q.; Poland, T.M.; Du, J.W. Parasitism and olfactory responses of Dastarcus helophoroides (Coleoptera: Bothrideridae) to different Cerambycid hosts. Biocontrol 2009, 54, 733–742. [Google Scholar] [CrossRef]
- Wang, X.M.; Ren, G.D.; Ma, F. Classification position of Dastarcus helophoroides and its applied prospects. Acta Agric. Bor-Occid. Sin. 1996, 5, 75–78. [Google Scholar]
- Zhang, Y.N.; Yang, Z.Q. Studies on the natural enemies and biocontrol of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Plant Prot. 2006, 32, 9–14. [Google Scholar]
- Li, M.L.; Wang, P.X.; Ma, F.; Yang, Z.Q. Study on the Parasitic Efficiency of Dastarcus helophoroides on Anoplophora glabripennis. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-XBNY200706030.htm (accessed on 15 October 2016). (In Chinese)
- Wang, H.D.; Li, F.F.; He, C.; Cui, J.; Song, W.; Li, M.L. Molecular cloning and sequence analysis of novel cytochrome P450 cDNA fragments from Dastarcus helophoroides. J. Insect Sci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Li, M.L.; Yang, Z.Q. A study on biological feature of Dastarcus longulus. J. Northwest Sci.-Tech. Univ. Agric. For. (Natl. Sci. Ed.) 2003, 31, 62–66. (In Chinese) [Google Scholar]
- Zhang, W.; Song, W.; Zhang, Z.Q.; Wang, H.D.; Yang, M.M.; Guo, R.J.; Li, M.L. Transcriptome Analysis of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Illumina HiSeq Sequencing. PLoS ONE 2014, 9, e100673. [Google Scholar] [CrossRef] [PubMed]
- BLAST, BlastX. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastx&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 15 October 2016).
- National Center of Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 15 October 2016).
- ORFfinder. Available online: https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 15 October 2016).
- Qiao, L.; Wu, J.X.; Qin, D.Z.; Lu, Z.C.; Pan, Z.L.; Chen, H.; Li, G.W. Gene Expression profiles of heat shock proteins 70 and 90 from Empoasca onukii (Hemiptera: Cicadellidae) in response to temperature stress. J. Insect Sci. 2015, 15, 49–49. [Google Scholar] [CrossRef] [PubMed]
- ExPASy, Compute pI/Mw Tool. Available online: http://web.expasy.org/compute_pi/ (accessed on 15 October 2016).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis or relative gene expression data using real–time quantitative PCR and the 2–ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001. [Google Scholar] [CrossRef]
- Dai, L.; Wang, C.; Zhang, X.; Yu, J.; Zhang, R.; Chen, H. Two CYP4 genes of the Chinese white pine beetle, Dendroctonus armandi (Curculionidae: Scolytinae), and their transcript levels under different development stages and treatments. Insect Mol. Biol. 2014, 23, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Paaby, A.B.; Schmidt, P.S. Functional significance of allelic variation at methuselah, an aging gene in Drosophila. PLoS ONE 2008, 3, e1987. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Zhang, Y.; Yun, X.P.; Wang, Y.Y.; Sang, M.; Liu, X.; Hu, X.X.; Li, B. Methuselah-like genes affect development, stress resistance, lifespan and reproduction in Tribolium castaneum. Insect Mol. Biol. 2014, 23, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, A.; Jones, J.W.; Friedrich, M. Methuselah/Methuselah-like G protein-coupled receptors constitute an ancient metazoan gene family. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, L.E.; Ghildyal, P.; Fischer, K.E.; Hu, H.X.; Ja, W.W.; Eaton, B.A.; Wu, Y.M.; Austad, S.N.; Ranjan, R. Modulation of methuselah expression targeted to Drosophila insulin-producing cells extends life and enhances oxidative stress resistance. Aging Cell 2013, 12, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Hwangbo, D.S.; Gershman, B.; Tu, M.P.; Palmer, M.; Tatar, M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature 2004, 429, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK extends life span and limits growth by antagonizing cellular and organism-wide responses to insulin signaling. Cell 2005, 121, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.; Partridge, L. Insulin/IGF-like signalling, the central nervous system and aging. Biochem. J. 2009, 418, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hay, N. Interplay between FOXO, TOR, and Akt. Biochim. Biophys. Acta 2011, 1813, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Apfeld, J.; Kenyon, C. Regulation of lifespan by sensory perception in Caenorhabditis elegans. Nature 1999, 402, 804–809. [Google Scholar] [PubMed]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, H.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Bluher, M.; Kahn, B.B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | Oligo Sequence (5′ to 3′) |
|---|---|
| Specific primer for 5′ RACE | |
| Mth-like1 5′GSP1 | TCACCAATAGATGGAAGTCCCTGAGAGC |
| Mth-like1 5′GSP2 | CGCCGATTGGACCGTAGAAGAATGTAAG |
| Mth-like2 5′GSP1 | CAACAGTTGTGGTGTGCCATCTCTAATAAA |
| Mth-like2 5′GSP2 | CAAGAAGGACAGCCATACCCCAAGCATA |
| Mth-like5 5′GSP1 | TTGGAGAACGACGACAGGAAGAACAGC |
| Mth-like5 5′GSP2 | ACGACCAAGCGTATGCCGAATAGTAGC |
| Specific primer for 3′ RACE | |
| Mth-like1 3′GSP1 | TCTTCTCATCGGCGACATTCTCCTCTGC |
| Mth-like1 3′GSP2 | ATTCTTCTACGGTCCAATCGGCGTTCTT |
| Mth-like2 3′GSP1 | AGTTGGTATCATCCGGAAGTTGGCAGAG |
| Mth-like2 3′GSP2 | CTTTATTAGAGATGGCACACCACAACTG |
| Mth-like5 3′GSP1 | GTGGCTGTTCTTCCTGTCGTCGTTCTC |
| Mth-like5 3′GSP2 | GCTGGGGTCCGTCCAAAGGAGGTGATA |
| Specific primer for qPCR | |
| Mth-like1 S | GCCAACCACAAGTCCTATCAGCT |
| Mth-like1 A | CACCAATAGATGGAAGTCCCTGA |
| Mth-like2 S | TTTGGTTAGGATTCAGCGGACT |
| Mth-like2 A | GGACAGCCATACCCCAAGCA |
| Mth-like5 S | ACTATTCGGCATACGCTTGGTC |
| Mth-like5 A | CAGTGATCCGATTTCTTCCCG |
| Reference primer for qPCR | |
| α-tubulin S | TCGGTGGTGGTACTGGGTCT |
| α-tubulin A | ACGGCTGTTGAAACTTGAGGA |
| EF-1α S | TCCTTCAAATATGCCTGGGTACT |
| EF-1α A | AAATCTCTGTGTGTCCAGGGGCAT |
| Gene Name | Length (bp) | ORF (bp) | Protein Length (aa) | Isoelectric Point | Molecular Weight (kDa) | Accession Number |
|---|---|---|---|---|---|---|
| mth-like1 | 2600 | 1701 | 566 | 8.67 | 63.8 | KM588897 |
| mth-like2 | 1875 | 1353 | 450 | 8.06 | 51.8 | KU363815 |
| mth-like5 | 2312 | 1428 | 475 | 6.01 | 54.4 | KU363816 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wang, H.; Hao, C.; Zhang, W.; Yang, M.; Chang, Y.; Li, M. Identification, Characterization and Expression of Methuselah-Like Genes in Dastarcus helophoroides (Coleoptera: Bothrideridae). Genes 2016, 7, 91. https://doi.org/10.3390/genes7100091
Zhang Z, Wang H, Hao C, Zhang W, Yang M, Chang Y, Li M. Identification, Characterization and Expression of Methuselah-Like Genes in Dastarcus helophoroides (Coleoptera: Bothrideridae). Genes. 2016; 7(10):91. https://doi.org/10.3390/genes7100091
Chicago/Turabian StyleZhang, Zhengqing, Huapeng Wang, Chunfeng Hao, Wei Zhang, Miaomiao Yang, Yong Chang, and Menglou Li. 2016. "Identification, Characterization and Expression of Methuselah-Like Genes in Dastarcus helophoroides (Coleoptera: Bothrideridae)" Genes 7, no. 10: 91. https://doi.org/10.3390/genes7100091
APA StyleZhang, Z., Wang, H., Hao, C., Zhang, W., Yang, M., Chang, Y., & Li, M. (2016). Identification, Characterization and Expression of Methuselah-Like Genes in Dastarcus helophoroides (Coleoptera: Bothrideridae). Genes, 7(10), 91. https://doi.org/10.3390/genes7100091