
Error-Free Bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by DNA Polymerase of Pseudomonas aeruginosa Phage PaP1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Construction, Expression, and Purification of Gp90 Mutants
- E60A:
- sense, 5′-GTAGTAGCCGCCCACGGCGGTAACATTCTGGCGTTCTAC-3′;antisense, 5′-GCC GTGGGCGGCTACTACGACACAGTGATGACTGTAGCT-3′.
- D137A:
- sense, 5′-ATTAACTTCGCCCTTATGTCGATGAAGCTTGTGGAAGATATG-3′;antisense, 5′-CGACATAAGGGCGAAGTTAATCATGTTGTGAGCCACTACGCG-3′.
- D234A:
- sense, 5′-TGTATCTATGCCGTAAAGGCGAACACCGCTGTATGGCACTGG-3′;antisense, 5′-CGCCTTTACGGCATAGATACAGTAGTAAAGCATATCGGCTGC-3′.
2.3. Examination of Exonuclease Activities of Gp90 Mutants
2.4. Primer Extension by Gp90 Mutants Using All Four dNTPs
2.5. Steady-State Kinetics Analysis of Single-Base Incorporation and Next-Base Extension
2.6. Pre-Steady-State Kinetic Analysis
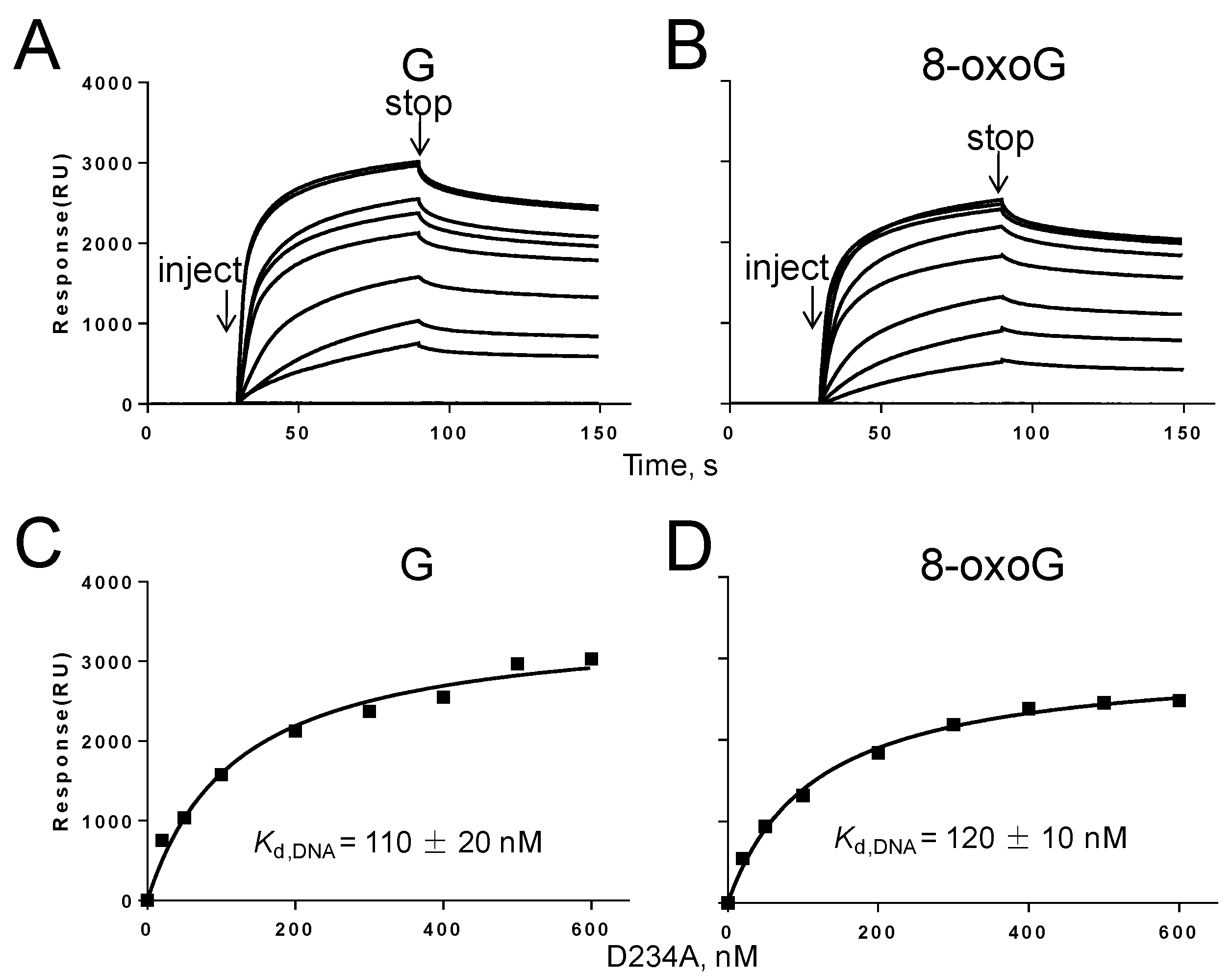
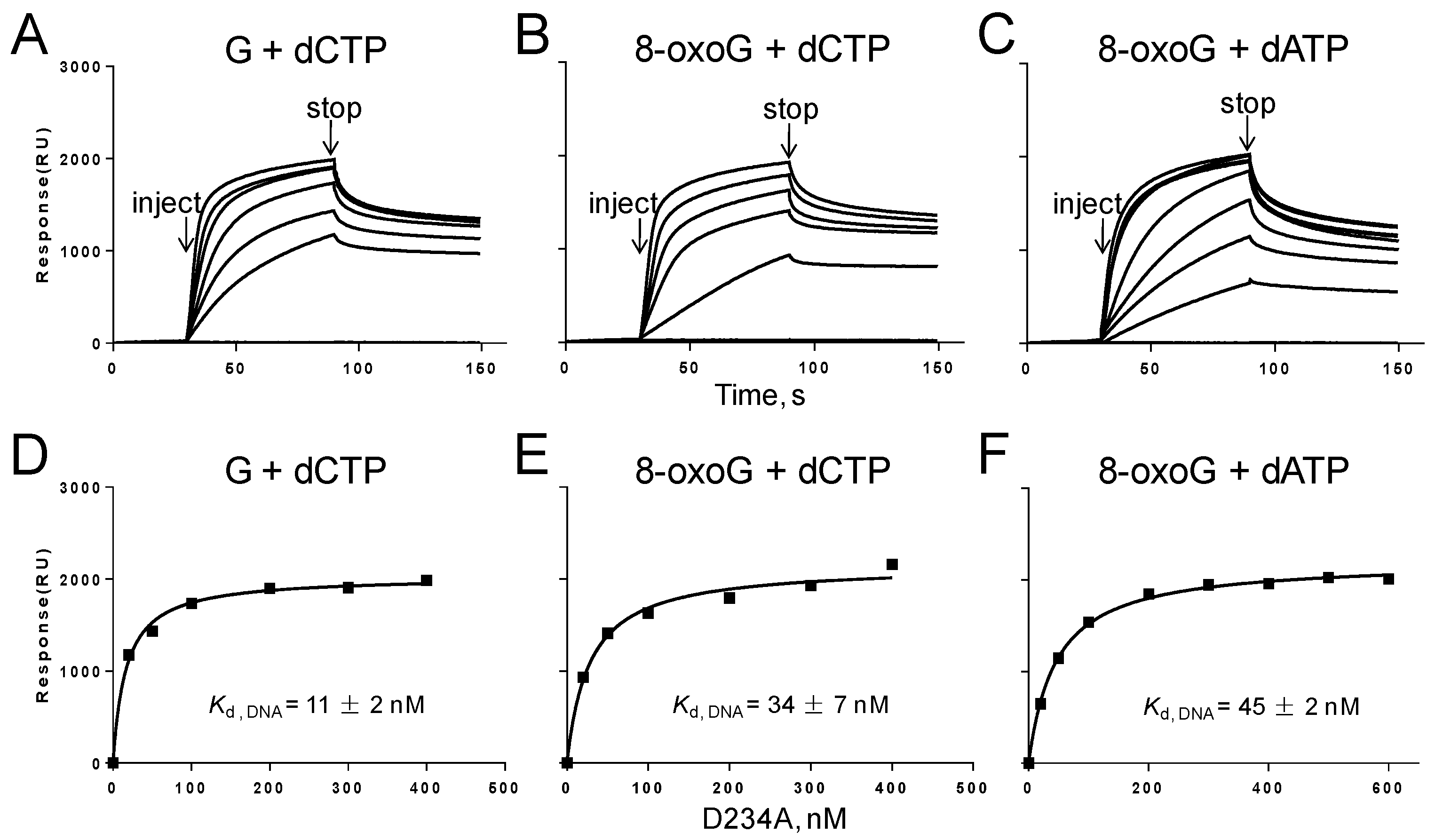
2.7. Biophysical Binding of Gp90 D234A to DNA Containing G or 8-oxoG
3. Results
3.1. Examination of Exonuclease and Polymerase Activities of Gp90 Mutants
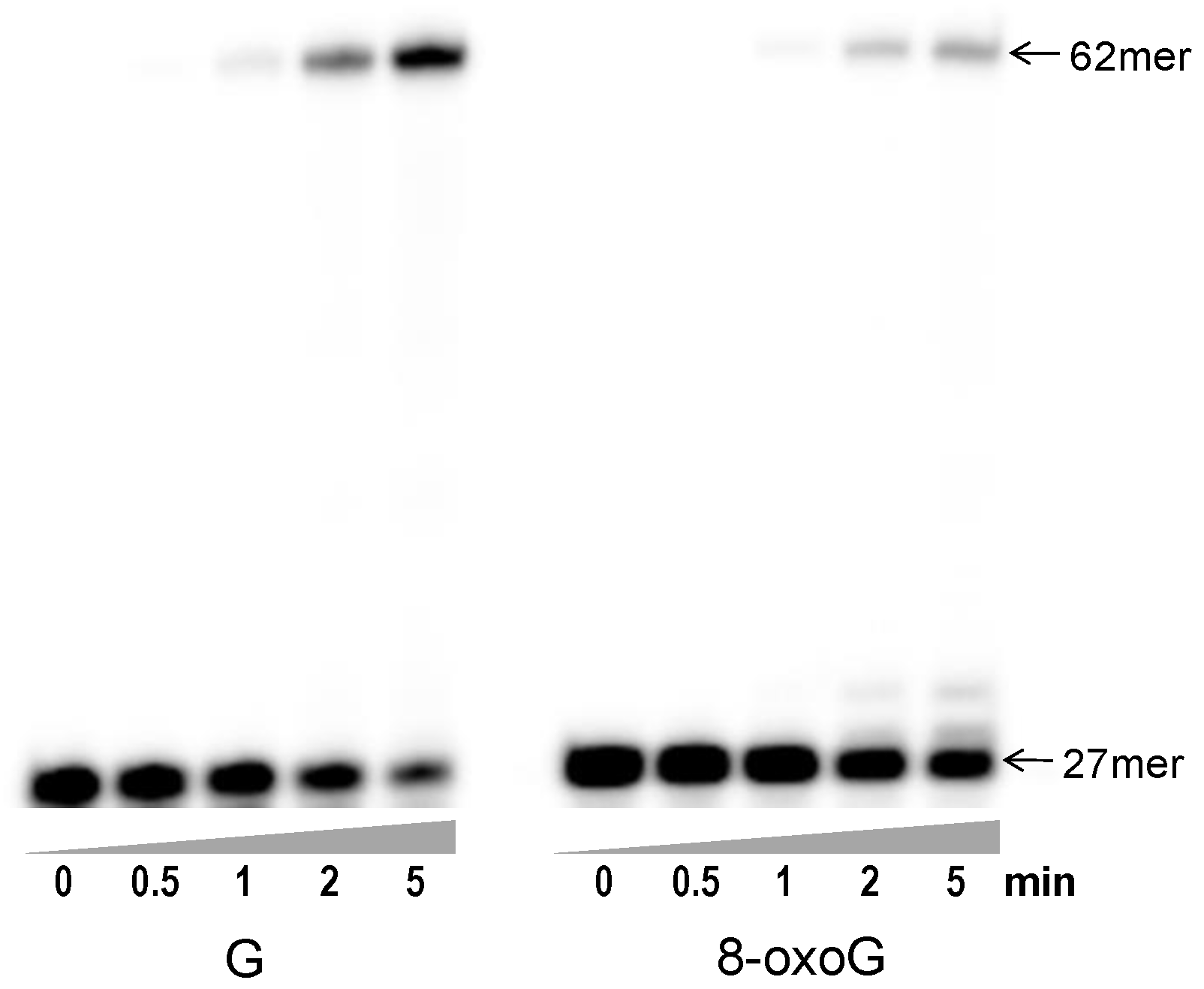
3.2. Primer Extension beyond 8-oxoG by Gp90 D234A Using All Four dNTPs
3.3. Steady-State Kinetic Analysis of Single-Base Incorporation Opposite G or 8-oxoG by Gp90 D234A
3.4. Steady-State Kinetic Analysis of Next-Base Extension beyond G or 8-oxoG by Gp90 D234A
3.5. Pre-Steady-State Kinetic Analysis of Single dNTP Incorporation by Gp90 D234A
3.6. Binding of Gp90 D234A to the PrimerTemplate Containing 8-oxoG
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hogg, M.; Rudnicki, J.; Midkiff, J.; Reha-Krantz, L.; Doublie, S.; Wallace, S.S. Kinetics of mismatch formation opposite lesions by the replicative DNA polymerase from bacteriophage RB69. Biochemistry 2010, 49, 2317–2325. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.; Nagy, L.D.; Zhang, Q.; Su, Y.; Muller, L.; Guengerich, F.P.; Egli, M. Kinetics, structure, and mechanism of 8-oxo-7,8-dihydro-2′-deoxyguanosine bypass by human DNA polymerase eta. J. Biol. Chem. 2014, 289, 16867–16882. [Google Scholar] [CrossRef] [PubMed]
- Taggart, D.J.; Fredrickson, S.W.; Gadkari, V.V.; Suo, Z. Mutagenic potential of 8-oxo-7,8-dihydro-2′-deoxyguanosine bypass catalyzed by human Y-family DNA polymerases. Chem. Res. Toxicol. 2014, 27, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Sasaki, S.; Taniguchi, Y. Effects of 8-halo-7-deaza-2′-deoxyguanosine triphosphate on DNA synthesis by DNA polymerases and cell proliferation. Bioorg. Med. Chem. 2016, 24, 3856–3861. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.; Wang, M.; Blaha, G.; Wang, J.; Konigsberg, W.H. Substitution of Ala for Tyr567 in RB69 DNA polymerase allows dAMP to be inserted opposite 7,8-dihydro-8-oxoguanine. Biochemistry 2010, 49, 4116–4125. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Schorr, S.; Carell, T. Crystal structure analysis of DNA lesion repair and tolerance mechanisms. Curr. Opin. Struct. Biol. 2009, 19, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Vooradi, V.; Romano, L.J. Effect of N-2-Acetylaminofluorene and 2-Aminofluorene Adducts on DNA Binding and Synthesis by Yeast DNA Polymerase η. Biochemistry 2009, 48, 4209–4216. [Google Scholar] [CrossRef] [PubMed]
- Swanson, A.L.; Wang, J.; Wang, Y. In Vitro Replication Studies of Carboxymethylated DNA Lesions with Saccharomyces cerevisiae Polymerase η. Biochemistry 2011, 50, 7666–7673. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xue, Q.; Tang, Y.; Cao, J.; Guengerich, F.P.; Zhang, H. Mechanisms of mutagenesis: DNA replication in the presence of DNA damage. Mutat. Res. Rev. Mutat. Res. 2016, 768, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Freudenthal, B.D.; Beard, W.A.; Wilson, S.H. DNA polymerase minor groove interactions modulate mutagenic bypass of a templating 8-oxoguanine lesion. Nucleic Acids Res. 2013, 41, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Patro, J.N.; Urban, M.; Kuchta, R.D. Interaction of Human DNA Polymerase α and DNA Polymerase I from Bacillus stearothermophilus with Hypoxanthine and 8-oxoguanine Nucleotides. Biochemistry 2009, 48, 8271–8278. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, B.A.; Suo, Z. Kinetic basis for the differing response to an oxidative lesion by a replicative and a lesion bypass DNA polymerase from Sulfolobus solfataricus. Biochemistry 2012, 51, 3485–3496. [Google Scholar] [CrossRef] [PubMed]
- Eoff, R.L.; Irimia, A.; Angel, K.C.; Egli, M.; Guengerich, F.P. Hydrogen bonding of 7,8-dihydro-8-oxodeoxyguanosine with a charged residue in the little finger domain determines miscoding events in Sulfolobus solfataricus DNA polymerase Dpo4. J. Biol. Chem. 2007, 282, 19831–19843. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Song, I.; Jo, A.; Shin, J.H.; Cho, H.; Eoff, R.L.; Guengerich, F.P.; Choi, J.Y. Biochemical analysis of six genetic variants of error-prone human DNA polymerase iota involved in translesion DNA synthesis. Chem. Res. Toxicol. 2014, 27, 1837–1852. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Duym, W.W.; Fowler, J.D.; Suo, Z. Single-turnover kinetic analysis of the mutagenic potential of 8-oxo-7,8-dihydro-2′-deoxyguanosine during gap-filling synthesis catalyzed by human DNA polymerases lambda and beta. J. Mol. Biol. 2007, 367, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- Irimia, A.; Eoff, R.L.; Guengerich, F.P.; Egli, M. Structural and functional elucidation of the mechanism promoting error-prone synthesis by human DNA polymerase kappa opposite the 7,8-dihydro-8-oxo-2′-deoxyguanosine adduct. J. Biol. Chem. 2009, 284, 22467–22480. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Gu, S.; Liang, N.; Xiong, M.; Xue, Q.; Lu, S.; Hu, F.; Zhang, H. Pseudomonas aeruginosa phage PaP1 DNA polymerase is an A-family DNA polymerase demonstrating ssDNA and dsDNA 3′–5′ exonuclease activity. Virus Genes 2016, 52, 538–551. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Le, S.; Tan, Y.; Zhu, J.; Li, M.; Rao, X.; Zou, L.; Li, S.; Wang, J.; Jin, X. Genomic and proteomic analyses of the terminally redundant genome of the Pseudomonas aeruginosa phage PaP1: Establishment of genus PaP1-like phages. PLoS ONE 2013, 8, e62933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guengerich, F.P. Effect of N 2 -Guanyl Modifications on Early Steps in Catalysis of Polymerization by Sulfolobus solfataricus P2 DNA Polymerase Dpo4 T239W. J. Mol. Biol. 2010, 395, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Washington, M.T.; Wolfle, W.T.; Spratt, T.E.; Prakash, L.; Prakash, S. Yeast DNA polymerase eta makes functional contacts with the DNA minor groove only at the incoming nucleoside triphosphate. Proc. Natl. Acad. Sci. USA 2003, 100, 5113–5118. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Le, S.; Tan, Y.; Li, M.; Liu, C.; Zhang, K.; Huang, J.; Chen, H.; Rao, X.; Zhu, J. Unlocking the mystery of the hard-to-sequence phage genome: PaP1 methylome and bacterial immunity. BMC Genom. 2014, 15, 803. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yong, T.; Seung-Joo, L.; Wei, Z.; Jia, C.; Richardson, C.C. Helicase-DNA polymerase interaction is critical to initiate leading-strand DNA synthesis. Proc. Natl. Acad. Sci. USA 2011, 108, 9372–9377. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Rong, W.; Liu, B.; Xue, Q.; Zhong, M.; Hao, Z.; Zhang, H. Kinetic analysis of bypass of abasic site by the catalytic core of yeast DNA polymerase eta. Mutat. Res. 2015, 779, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Zhong, M.; Liu, B.; Tang, Y.; Wei, Z.; Guengerich, F.P.; Zhang, H. Kinetic analysis of bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by the catalytic core of yeast DNA polymerase η. Biochimie 2016, 121, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Eoff, R.L.; Kozekov, I.D.; Rizzo, C.J.; Egli, M.; Guengerich, F.P. Versatility of Y-family Sulfolobus solfataricus DNA Polymerase Dpo4 in Translesion Synthesis Past Bulky N2-Alkylguanine Adducts. J. Biol. Chem. 2008, 284, 3563–3576. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Eoff, R.L.; Kozekov, I.D.; Rizzo, C.J.; Egli, M.; Guengerich, F.P. Structure-Function Relationships in Miscoding by Sulfolobus solfataricus DNA Polymerase Dpo4: Guanine N2,N2-dimethyl substitution produces inactive and miscoding polymerase complexes. J. Biol. Chem. 2009, 284, 17687–17699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bren, U.; Kozekov, I.D.; Rizzo, C.J.; Stec, D.F.; Guengerich, F.P. Steric and Electrostatic Effects at the C2 Atom Substituent Influence Replication and Miscoding of the DNA Deamination Product Deoxyxanthosine and Analogs by DNA Polymerases. J. Mol. Biol. 2009, 392, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, Y.; Lee, S.-J.; Wei, Z.; Cao, J.; Richardson, C.C. Binding Affinities among DNA Helicase-Primase, DNA Polymerase, and Replication Intermediates in the Replisome of Bacteriophage T7. J. Biol. Chem. 2016, 291, 1472–1480. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Xue, Q.; Gu, S.; Wang, W.; Chen, J.; Li, Y.; Wang, C.; Zhang, H. Kinetic analysis of bypass of O(6)-methylguanine by the catalytic core of yeast DNA polymerase eta. Arch. Biochem. Biophys. 2016, 596, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Carlson, K.D.; Washington, M.T. Mechanism of efficient and accurate nucleotide incorporation opposite 7,8-dihydro-8-oxoguanine by Saccharomyces cerevisiae DNA polymerase eta. Mol. Cell. Biol. 2005, 25, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Katafuchi, A.; Nohmi, T. DNA polymerases involved in the incorporation of oxidized nucleotides into DNA: Their efficiency and template base preference. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 703, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Taggart, D.J.; Dayeh, D.M.; Fredrickson, S.W.; Suo, Z. N-terminal domains of human DNA polymerase lambda promote primer realignment during translesion DNA synthesis. DNA Repair 2014, 22, 41–52. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 27-mer | 5′-GCTACAGAGTTATGGTGACGATACGTC-3′ |
| 28C-mer | 5′-GCTACAGAGTTATGGTGACGATACGTCC-3′ |
| 28A-mer | 5′-GCTACAGAGTTATGGTGACGATACGTCA-3′ |
| 30-mer | 5′-TTTGCTACAGAGTTATGGTGACGATACGTCdd-3′ |
| 62-mer | 3′-CGATGTCTCAATACCACTGCTATGCAGG*CTATCTCGCCTAATGATATGATGTAAT CTTAAGT-5′ |
| Template Base | dNTP | Km,dNTP μM | kcat, ×10−3min−1 | kcat/Km, μM−1min−1 | Misincorporation Frequency |
|---|---|---|---|---|---|
| G | C | (1.3 ± 0.1) × 10−3 | 840 ± 10 | 650 | |
| A | 12 ± 1 | 750 ± 50 | 0.06 | 9.3 × 10−5 | |
| G | 7.7 ± 0.5 | 800 ± 10 | 0.10 | 1.5 × 10−4 | |
| T | 4.2 ± 0.5 | 430 ± 8 | 0.10 | 1.5 × 10−4 | |
| 8-oxoG | C | 2.9 ± 0.1 | 850 ± 10 | 0.85 | |
| A | 300 ± 36 | 190 ± 8 | 6.1 × 10−4 | 7.2 × 10−4 | |
| G | 73 ± 9 | 0.15 ± 0.01 | 2.1 × 10−6 | 2.5 × 10−4 | |
| T | 120 ± 9 | 0.25 ± 0.01 | 8.3 × 10−6 | 1.0 × 10−5 |
| Template Base | Primer X | Km,dGTP μM | kcat, ×10−2min−1 | kcat/Km, μM−1min−1 | Efficiency relative to G:C |
|---|---|---|---|---|---|
| G | C | 0.05 ± 0.01 | 49 ± 1 | 9.8 | 1 |
| A | 0.04 ± 0.01 | 5 ± 0.2 | 1.3 | 8-fold less | |
| T | 0.05 ± 0.01 | 1 ± 0.1 | 0.2 | 49-fold less | |
| 8-oxoG | C | 20 ± 2 | 40 ± 1 | 2.0 × 10−2 | 490-fold less |
| A | 20 ± 2 | 46 ± 1 | 2.3 × 10−2 | 490-fold less |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.; Xue, Q.; Liu, Q.; Xiong, M.; Wang, W.; Zhang, H. Error-Free Bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by DNA Polymerase of Pseudomonas aeruginosa Phage PaP1. Genes 2017, 8, 18. https://doi.org/10.3390/genes8010018
Gu S, Xue Q, Liu Q, Xiong M, Wang W, Zhang H. Error-Free Bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by DNA Polymerase of Pseudomonas aeruginosa Phage PaP1. Genes. 2017; 8(1):18. https://doi.org/10.3390/genes8010018
Chicago/Turabian StyleGu, Shiling, Qizhen Xue, Qin Liu, Mei Xiong, Wanneng Wang, and Huidong Zhang. 2017. "Error-Free Bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by DNA Polymerase of Pseudomonas aeruginosa Phage PaP1" Genes 8, no. 1: 18. https://doi.org/10.3390/genes8010018
APA StyleGu, S., Xue, Q., Liu, Q., Xiong, M., Wang, W., & Zhang, H. (2017). Error-Free Bypass of 7,8-dihydro-8-oxo-2′-deoxyguanosine by DNA Polymerase of Pseudomonas aeruginosa Phage PaP1. Genes, 8(1), 18. https://doi.org/10.3390/genes8010018