
Contribution of the RgfD Quorum Sensing Peptide to rgf Regulation and Host Cell Association in Group B Streptococcus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, and rgf Sequence Analysis
2.2. rgfD Mutagenesis and Complementation
2.3. Association Assays
2.4. RNA Extraction, Preparation, and Quantitation
2.5. Statistical Analysis
3. Results
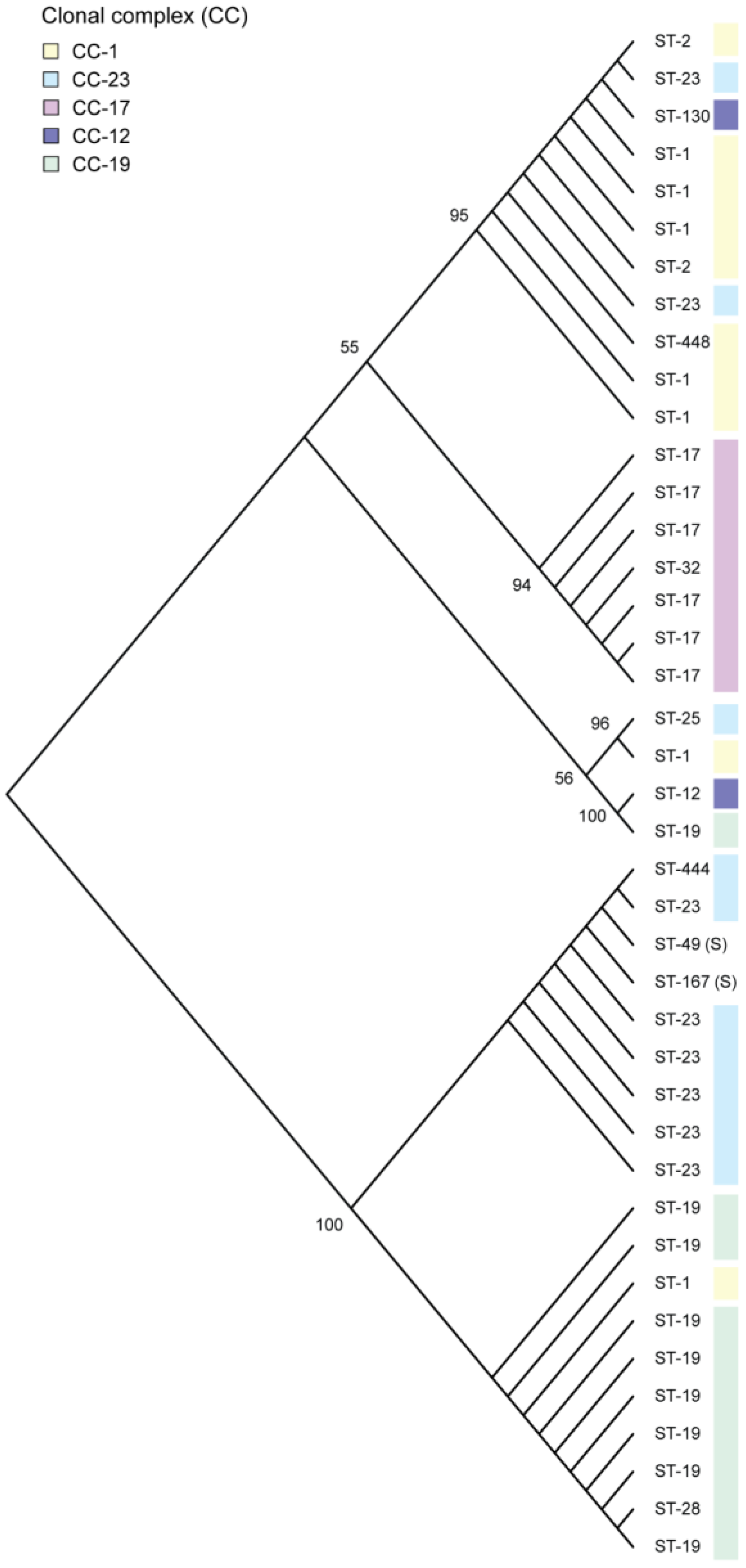
3.1. Allelic Variation in rgf among Diverse GBS Lineages
3.2. Association between rgf Variation and Biofilm Production
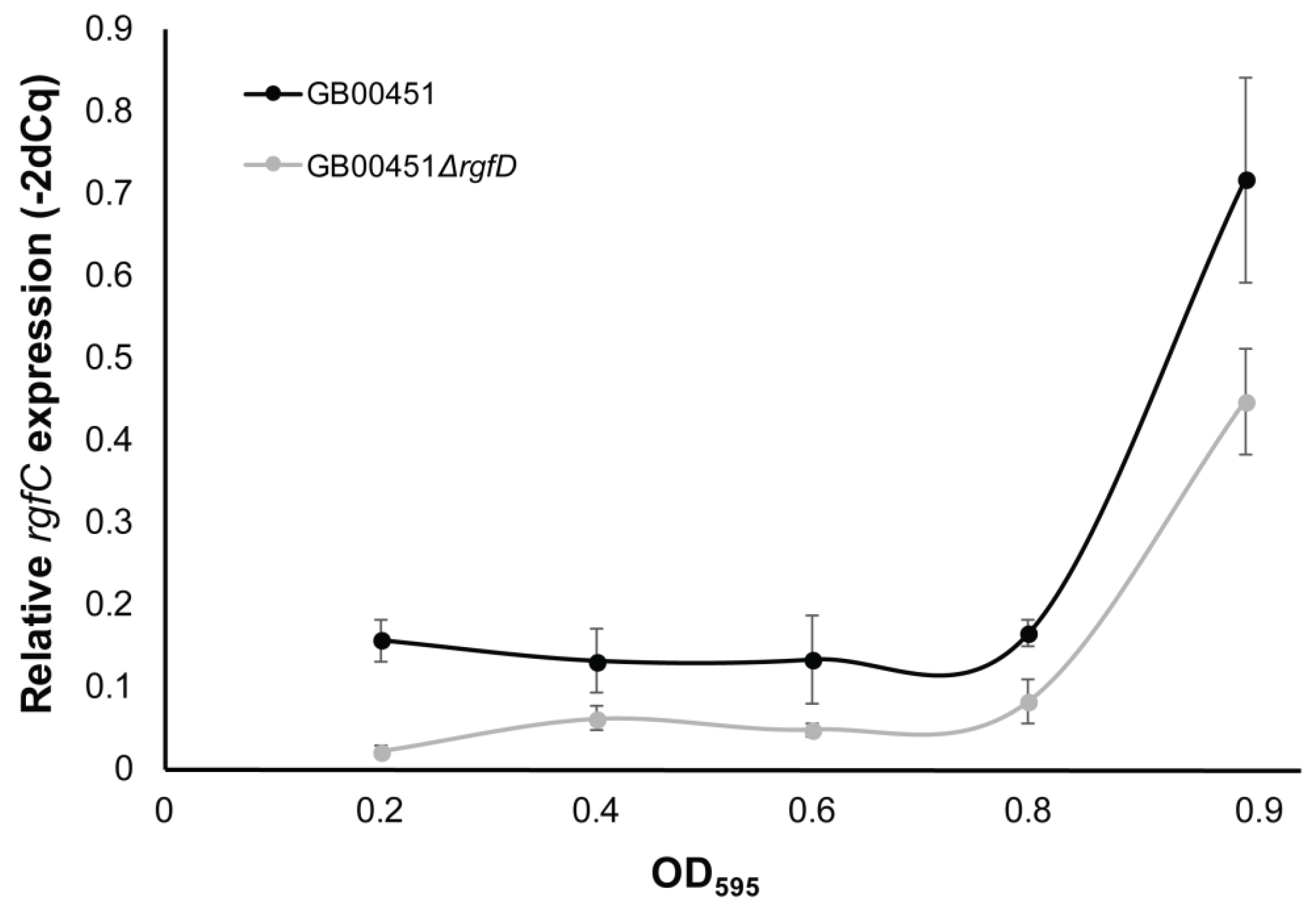
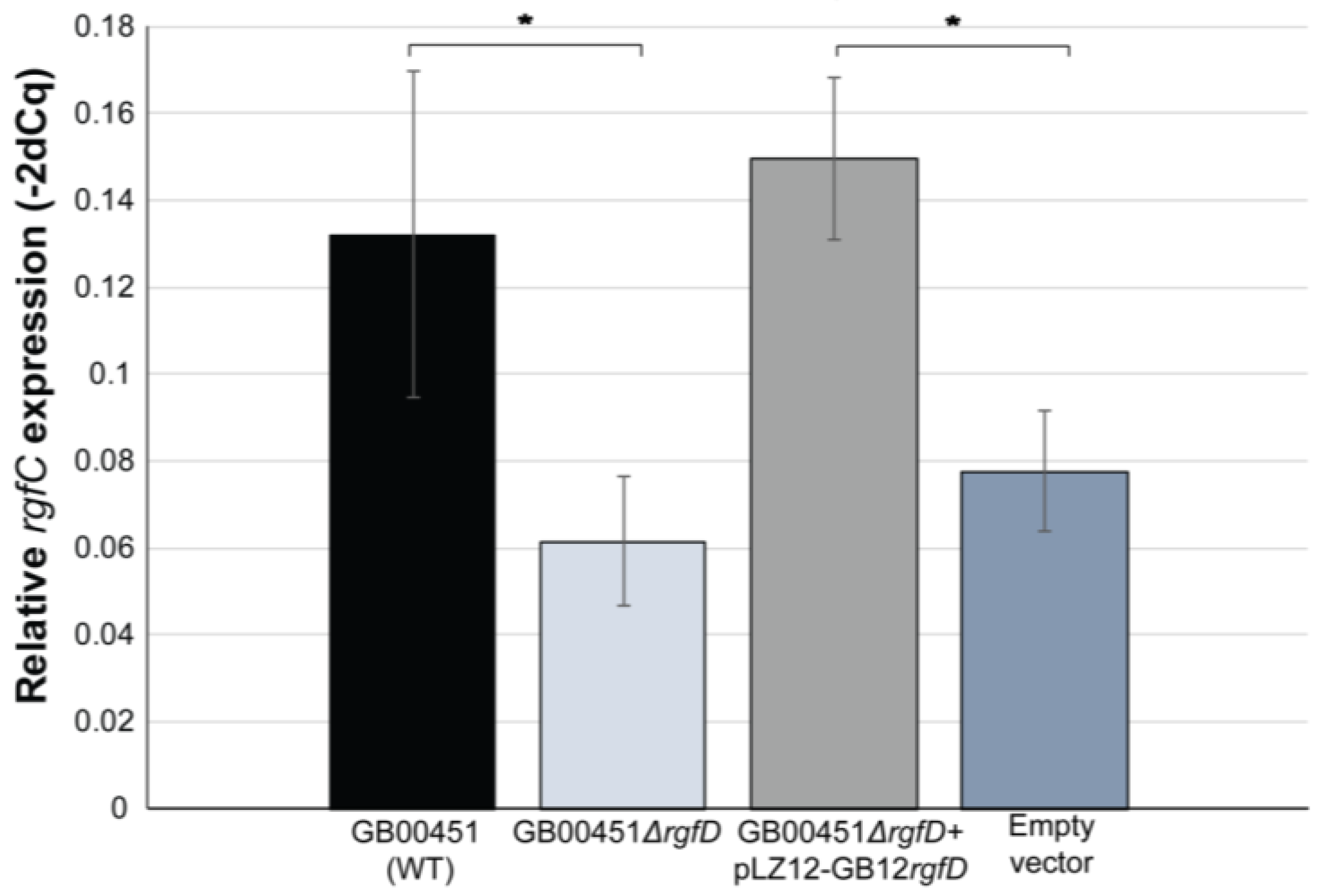
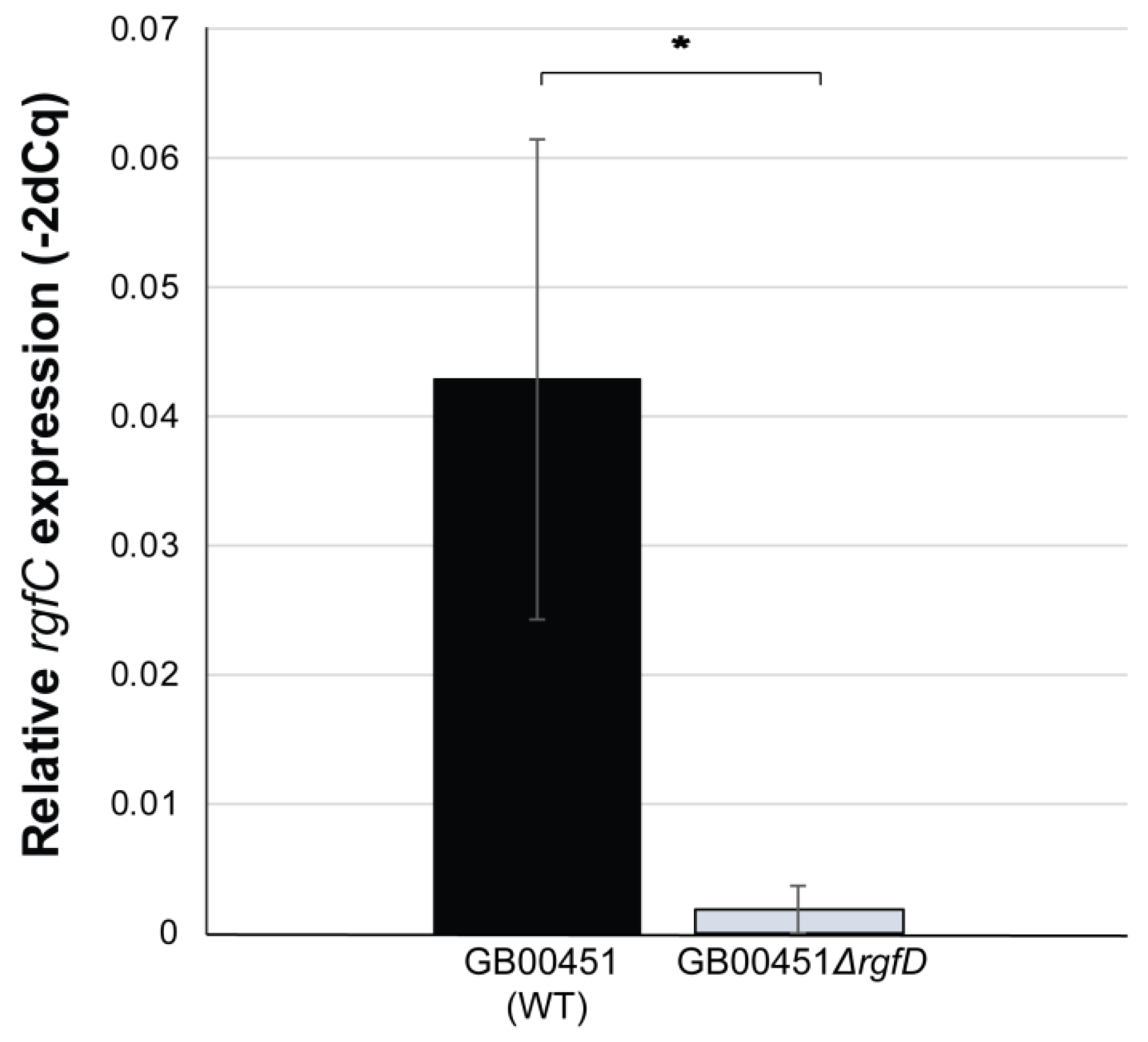
3.3. rgfD-Dependent Expression of the rgf Operon
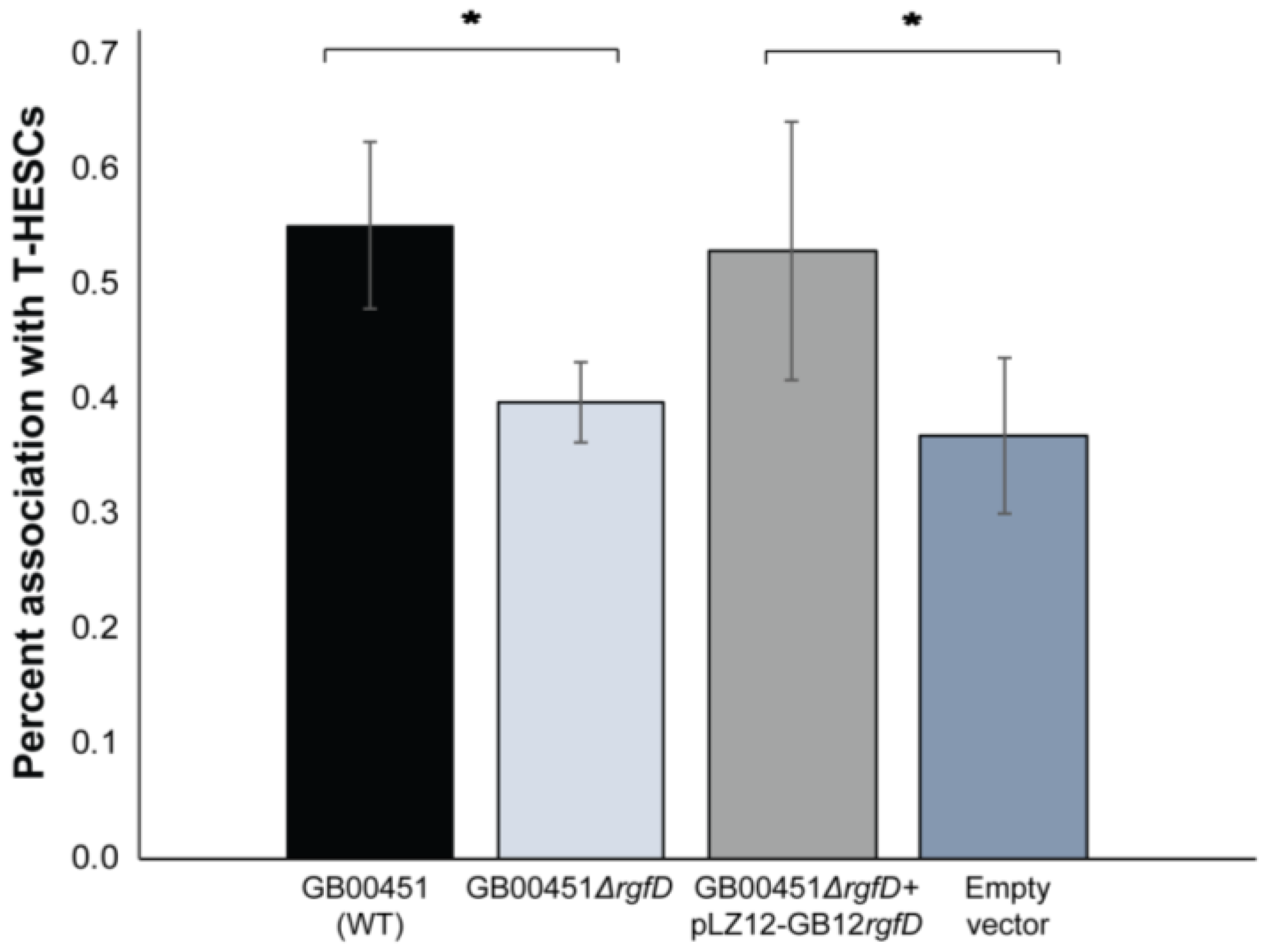
3.4. Role of rgfD in Association with Host Cells and Biofilm Production
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
Abbreviations
| GBS | Group B Streptococcus |
| qPCR | quantitative polymerase chain reaction |
| EOD | early onset disease |
| MLST | multilocus sequence type |
| STS | signal transduction system |
| TCS | two component system |
| ST | sequence type |
| CC | clonal complex |
| THB | Todd-Hewitt broth |
| THG | Todd-Hewitt broth plus 1% glucose |
| THA | Todd-Hewitt agar |
| PBS | phosphate-buffered saline |
| T-HESC | Telomerase-immortalized human endometrial stromal cells |
| MOI | multiplicity of infection |
| SNP | single nucleotide polymorphism |
References
- Schuchat, A.; Wenger, J.D. Epidemiology of group B streptococcal disease. Risk factors, prevention strategies, and vaccine development. Epidemiol. Rev. 1994, 16, 374–402. [Google Scholar] [PubMed]
- Phares, C.R.; Lynfield, R.; Farley, M.M.; Mohle-boetani, J.; Harrison, L.H.; Petit, S.; Craig, A.S.; Schaffner, W.; Gershman, K.; Stefonek, K.R.; et al. Epidemiology of invasive group B streptococcal disease in the United States, 1999–2005. J. Am. Med. Assoc. 2008, 299, 2056–2065. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.-S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.M.; Harding, R.M. Multilocus sequence typing system for group B Streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Manning, S.D.; Springman, A.C.; Lehotzky, E.; Lewis, M.A.; Whittam, T.S.; Davies, H.D. Multilocus sequence types associated with neonatal group B streptococcal sepsis and meningitis in canada. J. Clin. Microbiol. 2009, 47, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Brochet, M.; Couve, E.; Zouine, M.; Vallaeys, T.; Rusniok, C.; Lamy, M.C.; Buchrieser, C.; Trieu-Cuot, P.; Kunst, F.; Poyart, C.; et al. Genomic diversity and evolution within the species Streptococcus agalactiae. Microbes Infect. 2006, 8, 1227–1243. [Google Scholar] [CrossRef] [PubMed]
- Bisharat, N.; Crook, D.W.; Leigh, J.; Harding, R.M.; Ward, P.N.; Coffey, T.J.; Maiden, M.C.; Peto, T.; Jones, N. Hyperinvasive neonatal group B Streptococcus has arisen from a bovine ancestor. J. Clin. Microbiol. 2004, 42, 2161–2167. [Google Scholar] [CrossRef] [PubMed]
- Springman, A.C.; Lacher, D.W.; Waymire, E.A.; Wengert, S.L.; Singh, P.; Zadoks, R.N.; Davies, H.D.; Manning, S.D. Pilus distribution among lineages of group B Streptococcus: An evolutionary and clinical perspective. BMC Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Springman, A.C.; Lacher, D.W.; Wu, G.; Milton, N.; Whittam, T.S.; Davies, H.D.; Manning, S.D. Selection, recombination, and virulence gene diversity among group B streptococcal genotypes. J. Bacteriol. 2009, 191, 5419–5427. [Google Scholar] [CrossRef] [PubMed]
- Doran, K.S.; Nizet, V. Molecular pathogenesis of neonatal group B streptococcal infection: No longer in its infancy. Mol. Microbiol. 2004, 54, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-H.; Stins, M.F.; Kim, K.S. Bacterial penetration across the blood-brain barrier during the development of neonatal meningitis. Microbes Infect. 2000, 2, 1237–1244. [Google Scholar] [CrossRef]
- Winzer, K.; Williams, P. Quorum sensing and the regulation of virulence gene expression in pathogenic bacteria. Int. J. Med. Microbiol. 2001, 291, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Mereghetti, L.; Sitkiewicz, I.; Green, N.M.; Musser, J.M. Remodeling of the Streptococcus agalactiae transcriptome in response to growth temperature. PLoS ONE 2008, 3, e2785. [Google Scholar] [CrossRef] [PubMed]
- Mereghetti, L.; Sitkiewicz, I.; Green, N.M.; Musser, J.M. Extensive adaptive changes occur in the transcriptome of Streptococcus agalactiae (group B Streptococcus) in response to incubation with human blood. PLoS ONE 2008, 3, e3143. [Google Scholar] [CrossRef] [PubMed]
- Sitkiewicz, I.; Green, N.M.; Guo, N.; Bongiovanni, A.M.; Witkin, S.S.; Musser, J.M. Transcriptome adaptation of group B Streptococcus to growth in human amniotic fluid. PLoS ONE 2009, 4, e6114. [Google Scholar] [CrossRef] [PubMed]
- Hoch, J.A. Two-component and phosphorelay signal transduction. Curr. Opin. Microbiol. 2000, 3, 165–170. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Quadri, L.E.N.; Kuipers, O.P.; de Vos, W.M. Quorum sensing by peptide pheromones and two-component signal-transduction systems in Gram-positive bacteria. Mol. Microbiol. 1997, 24, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, L.E.; Koonin, E.V.; Zhulin, I.B. One-component systems dominate signal transduction in prokaryotes. Trends Microbiol. 2005, 13, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [PubMed]
- Lamy, M.-C.; Zouine, M.; Fert, J.; Vergassola, M.; Couve, E.; Pellegrini, E.; Glaser, P.; Kunst, F.; Msadek, T.; Trieu-Cuot, P.; et al. CovS/CovR of group B Streptococcus: A two-component global regulatory system involved in virulence. Mol. Microbiol. 2004, 54, 1250–1268. [Google Scholar] [CrossRef] [PubMed]
- Poyart, C.; Pellegrini, E.; Marceau, M.; Baptista, M.; Jaubert, F.; Lamy, M.-C.; Trieu-Cuot, P. Attenuated virulence of Streptococcus agalactiae deficient in d-alanyl-lipoteichoic acid is due to an increased susceptibility to defensins and phagocytic cells. Mol. Microbiol. 2003, 49, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Quach, D.; van Sorge, N.M.; Kristian, S.a.; Bryan, J.D.; Shelver, D.W.; Doran, K.S. The ciar response regulator in group B Streptococcus promotes intracellular survival and resistance to innate immune defenses. J. Bacteriol. 2009, 191, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Spellerberg, B.; Rozdzinski, E.; Martin, S.; Weber-Heynemann, J.; Lütticken, R. rgf encodes a novel two-component signal transduction system of Streptococcus agalactiae. Infect. Immun. 2002, 70, 2434–2440. [Google Scholar] [CrossRef] [PubMed]
- Al Safadi, R.; Mereghetti, L.; Salloum, M.; Lartigue, M.-F.; Virlogeux-Payant, I.; Quentin, R.; Rosenau, A. Two-component system RgfA/C activates the fbsB gene encoding major fibrinogen-binding protein in highly virulent CC-17 clone group B Streptococcus. PLoS ONE 2011, 6, e14658. [Google Scholar] [CrossRef] [PubMed]
- Traber, K.E.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- Novick, R.P.; Projan, S.J.; Kornblum, J.; Ross, H.F.; Ji, G.; Kreiswirth, B.; Vandenesch, F.; Moghazeh, S. The agr p2 operon: An autocatalytic sensory transduction system in Staphylococcus aureus. Mol. Gen. Genet. 1995, 248, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Todd, D.A.; Velázquez, J.V.; Cech, N.B.; Sonenshein, A.L. Cody-mediated regulation of the Staphylococcus aureus agr system integrates nutritional and population density signals. J. Bacteriol. 2014, 196, 1184–1196. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.-T.; Cheung, A.L. Molecular interactions between two global regulators, sar and agr, in Staphylococcus aureus. J. Biol. Chem. 1998, 273, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.S.; Traber, K.E.; Corrigan, R.; Benson, S.A.; Musser, J.M.; Novick, R.P. The agr radiation: An early event in the evolution of staphylococci. J. Bacteriol. 2005, 187, 5585–5594. [Google Scholar] [CrossRef] [PubMed]
- Gendrin, C.; Lembo, A.; Whidbey, C.; Burnside, K.; Berry, J.; Ngo, L.; Banerjee, A.; Xue, L.; Arrington, J.; Doran, K.S.; et al. The sensor histidine kinase RgfC affects group B streptococcal virulence factor expression independent of its response regulator RgfA. Infect. Immun. 2015, 83, 1078–1088. [Google Scholar] [CrossRef] [PubMed]
- Glaser, P.; Rusniok, C.; Buchrieser, C.; Chevalier, F.; Frangeul, L.; Msadek, T.; Zouine, M.; Couvé, E.; Lalioui, L.; Poyart, C.; et al. Genome sequence of Streptococcus agalactiae, a pathogen causing invasive neonatal disease. Mol. Microbiol. 2002, 45, 1499–1513. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Manning, S.D.; Schaeffer, K.E.; Springman, A.C.; Lehotzky, E.; Lewis, M.A.; Ouellette, L.M.; Wu, G.; Moorer, G.M.; Whittam, T.S.; Davies, H.D. Genetic diversity and antimicrobial resistance in group B Streptococcus colonizing young, nonpregnant women. Clin. Infect. Dis. 2008, 47, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.E.; Laut, C.; Gaddy, J.A.; Zadoks, R.N.; Davies, H.D.; Manning, S.D. Association between genotypic diversity and biofilm production in group B Streptococcus. BMC Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Biswas, I.; Gruss, A.; Ehrlich, S.D.; Maguin, E. High-efficiency gene inactivation and replacement system for gram-positive bacteria. J. Bacteriol. 1993, 175, 3628–3635. [Google Scholar] [CrossRef] [PubMed]
- Schubert, A.; Zakikhany, K.; Schreiner, M.; Frank, R.; Spellerberg, B.; Eikmanns, B.J.; Reinscheid, D.J. A fibrinogen receptor from group B Streptococcus interacts with fibrinogen by repetitive units with novel ligand binding sites. Mol. Microbiol. 2002, 46, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Hanson, B.R.; Lowe, B.A.; Neely, M.N. Membrane topology and DNA-binding ability of the streptococcal cpsa protein. J. Bacteriol. 2011, 193, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Korir, M.L.; Knupp, D.; LeMerise, K.; Boldenow, E.; Loch-Caruso, R.; Aronoff, D.M.; Manning, S.D. Association and virulence gene expression vary among serotype III group B Streptococcus isolates following exposure to decidual and lung epithelial cells. Infect. Immun. 2014, 82, 4587–4595. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative Ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Faralla, C.; Metruccio, M.M.; de Chiara, M.; Mu, R.; Patras, K.A.; Muzzi, A.; Grandi, G.; Margarit, I.; Doran, K.S.; Janulczyk, R. Analysis of two-component systems in group B Streptococcus shows that RgfAC and the novel FspSR modulate virulence and bacterial fitness. mBio 2014. [Google Scholar] [CrossRef] [PubMed]
- Cafiso, V.; Bertuccio, T.; Santagati, M.; Demelio, V.; Spina, D.; Nicoletti, G.; Stefani, S. agr-genotyping and transcriptional analysis of biofilm-producing Staphylococcus aureus. FEMS Immunol. Med.Microbiol. 2007, 51, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Lerat, E.; Moran, N.A. The evolutionary history of quorum-sensing systems in bacteria. Mol. Biol. Evol. 2004, 21, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, S.; Maeda, T.; Hashimoto, N.; Imaizumi, K.; Kaidoh, T.; Hayakawa, Y. Variation of the agr locus in Staphylococcus aureus isolates from cows with mastitis. Vet. Microbiol. 2001, 79, 267–274. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.C.; Novick, R.P. Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proc. Natl. Acad. Sci. USA 1995, 92, 12055–12059. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.E.; Tang, J.L.; Feng, J.X.; Pan, M.Q.; Wilson, T.J.G.; Slater, H.; Dow, J.M.; Williams, P.; Daniels, M.J. A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol. Microbiol. 1997, 24, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Surette, M.G.; Bassler, B.L. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1998, 95, 7046–7050. [Google Scholar] [CrossRef] [PubMed]
- Vuong, C.; Saenz, H.L.; Götz, F.; Otto, M. Impact of the agr quorum-sensing system on adherence to polystyrene in Staphylococcus aureus. J. Infect. Dis. 2000, 182, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Nicolas, P.; Bessières, P.; Bolotin, A.; Monnet, V.; Gardan, R. A genome-wide survey of short coding sequences in streptococci. Microbiology 2007, 153, 3631–3644. [Google Scholar] [CrossRef] [PubMed]
- Fleuchot, B.; Gitton, C.; Guillot, A.; Vidic, J.; Nicolas, P.; Besset, C.; Fontaine, L.; Hols, P.; Leblond-Bourget, N.; Monnet, V.; et al. Rgg proteins associated with internalized small hydrophobic peptides: A new quorum-sensing mechanism in streptococci. Mol. Microbiol. 2011, 80, 1102–1119. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.; Guillot, A.; Wessner, F.; Algaron, F.; Besset, C.; Courtin, P.; Gardan, R.; Monnet, V. Control of the transcription of a short gene encoding a cyclic peptide in Streptococcus thermophilus: A new quorum-sensing system? J. Bacteriol. 2007, 189, 8844–8854. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pascual, D.; Gaudu, P.; Fleuchot, B.; Besset, C.; Rosinski-Chupin, I.; Guillot, A.; Monnet, V.; Gardan, R. RovS and its associated signaling peptide form a cell-to-cell communication system required for Streptococcus agalactiae pathogenesis. mBio 2015. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Accession Number | Strain | Accession Number |
|---|---|---|---|
| rgf reference sequence | AF390107.1 | GB00557 | GCA_000290235.1 |
| GB00002 | GCA_000289475.1 | GB00614 | GCA_000290335.1 |
| GB00012 | GCA_000288135.1 | GB00651 | GCA_000290375.1 |
| GB00013 | GCA_000288095.1 | GB00654 | GCA_000290395.1 |
| GB00020 | GCA_000288235.1 | GB00663 | GCA_000290435.1 |
| GB00082 | GCA_000288215.1 | GB00679 | GCA_000290475.1 |
| GB00083 | GCA_000288255.1 | GB00865 | GCA_000290495.1 |
| GB00092 | GCA_000290055.1 | GB00867 | GCA_000289595.1 |
| GB00097 | GCA_000289495.1 | GB00874 | GCA_000289615.1 |
| GB00111 | GCA_000290075.1 | GB00884 | GCA_000289635.1 |
| GB00112 | GCA_000291585.1 | GB00887 | GCA_000289655.1 |
| GB00115 | GCA_000290095.1 | GB00891 | GCA_000290215.1 |
| GB00190 | GCA_000290135.1 | GB00904 | GCA_000288375.1 |
| GB00206 | GCA_000289535.1 | GB00923 | GCA_000288475.1 |
| GB00226 | GCA_000288195.1 | GB00929 | GCA_000288515.1 |
| GB00241 | GCA_000288175.1 | GB00932 | GCA_000288535.1 |
| GB00245 | GCA_000288335.1 | GB00959 | GCA_000288615.1 |
| GB00279 | GCA_000288355.1 | GB00984 | GCA_000288655.1 |
| GB00300 | GCA_000289575.1 | GB00986 | GCA_000289715.1 |
| GB00555 | GCA_000290235.1 | GB00992 | GCA_000289735.1 |
| Primer/Gene | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| Mutagenesis | ||
| rgfD_del 1 & 2 | CCGCGGATCCCCACTTTTACTCATGGGTGACTT | CCCATCCACTAAACTTAAACAGCATTCCAAACTTTGTAAGGAGTC |
| rgfD_del 3 & 4 | TGTTTAAGTTTAGTGGATGGGTTTTATTCAACAGGCACGTTTAG | GGGGGTACCAAAACTTCTTCAATCCTTCTGCT |
| rgfD_del 5 & 6 | TCATACTCGTCGTGCTCTGG | CAACTCTATGTGACCTTAATGACG |
| plz12:rgfD | CGCGGATCCAGGAGGACAGCTATGCGAAGTTTGGAATGCATGAG | AAAACTGCAGTTCTCTCTAAACGTGCCTGTTG |
| qPCR Detection | ||
| gyrA | CGGGACACGTACAGGCTACT | CGATACGAGAAGCTCCCACA |
| rgfC | GCGAAGTAGTGAAGTTTCGCCCAT | CCGGTCTAAACTGGCTATTGCTCC |
| rgfB | GCAAGTACCATGAAGGGGTAGCG | TCAGCTACCAGAGCACGACGAGT |
| fbsB | GCGATTGTGAATAGAATGAGTG | ACAGAAGCGGCGATTTCATT |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parker, R.E.; Knupp, D.; Al Safadi, R.; Rosenau, A.; Manning, S.D. Contribution of the RgfD Quorum Sensing Peptide to rgf Regulation and Host Cell Association in Group B Streptococcus. Genes 2017, 8, 23. https://doi.org/10.3390/genes8010023
Parker RE, Knupp D, Al Safadi R, Rosenau A, Manning SD. Contribution of the RgfD Quorum Sensing Peptide to rgf Regulation and Host Cell Association in Group B Streptococcus. Genes. 2017; 8(1):23. https://doi.org/10.3390/genes8010023
Chicago/Turabian StyleParker, Robert E., David Knupp, Rim Al Safadi, Agnѐs Rosenau, and Shannon D. Manning. 2017. "Contribution of the RgfD Quorum Sensing Peptide to rgf Regulation and Host Cell Association in Group B Streptococcus" Genes 8, no. 1: 23. https://doi.org/10.3390/genes8010023
APA StyleParker, R. E., Knupp, D., Al Safadi, R., Rosenau, A., & Manning, S. D. (2017). Contribution of the RgfD Quorum Sensing Peptide to rgf Regulation and Host Cell Association in Group B Streptococcus. Genes, 8(1), 23. https://doi.org/10.3390/genes8010023