
Parameters for Successful Parental RNAi as An Insect Pest Management Tool in Western Corn Rootworm, Diabrotica virgifera virgifera

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Identification
2.2. cDNA Preparation and dsRNA Synthesis
2.3. pRNAi Phenotypes in Embryos and Ovaries
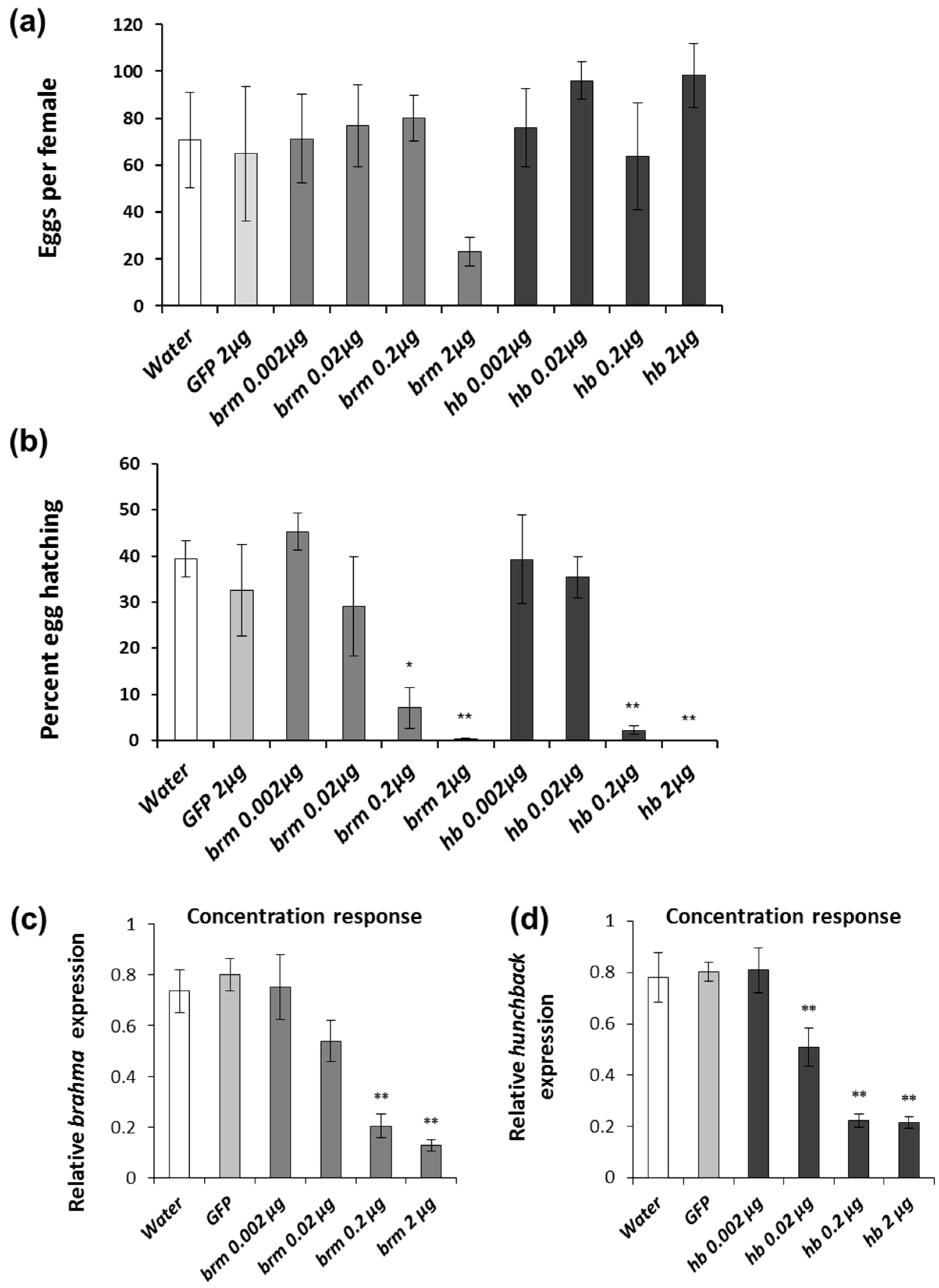
2.4. brahma and hunchback Concentration Response
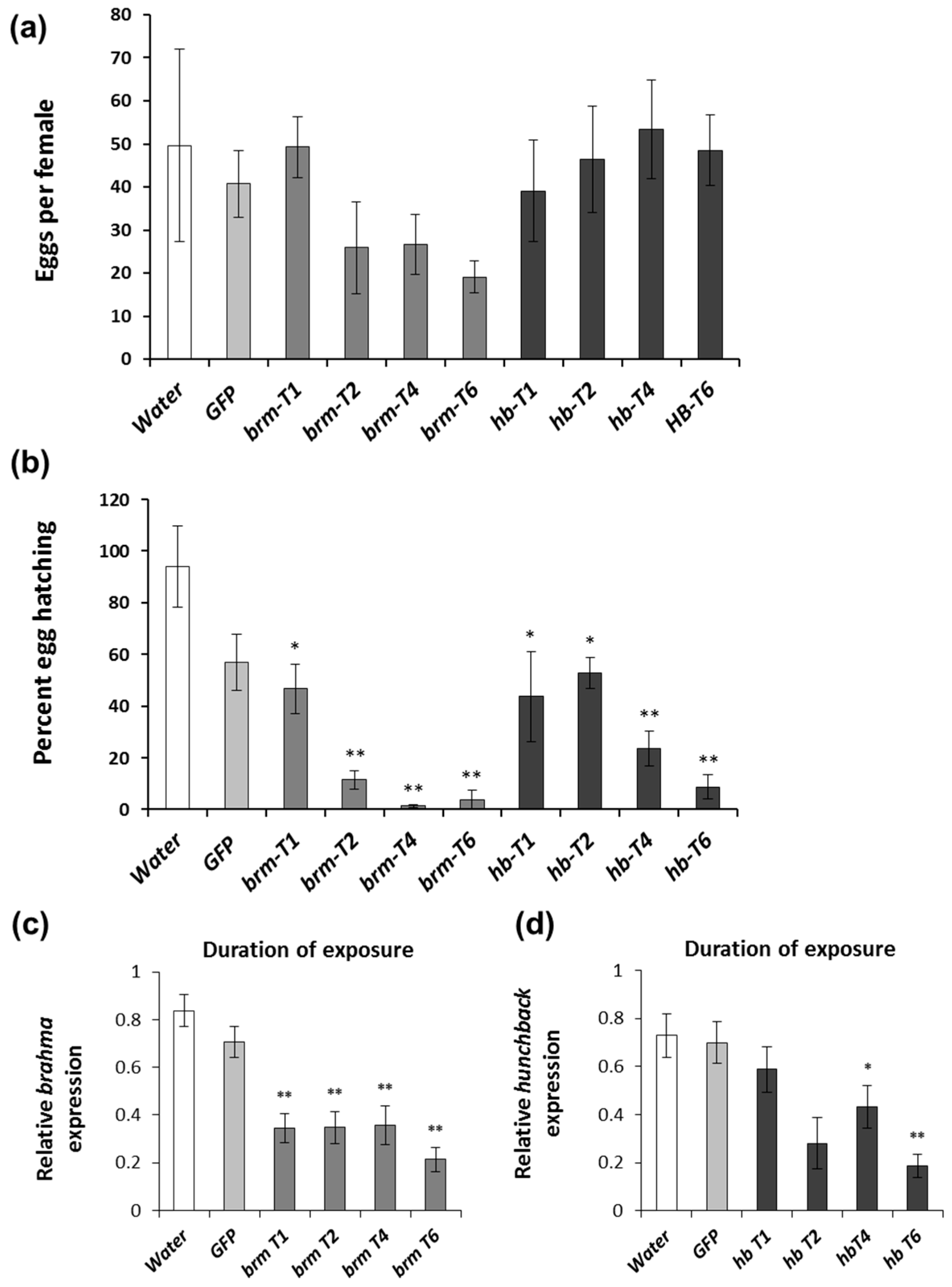
2.5. Duration of brahma and hunchback dsRNA Exposure
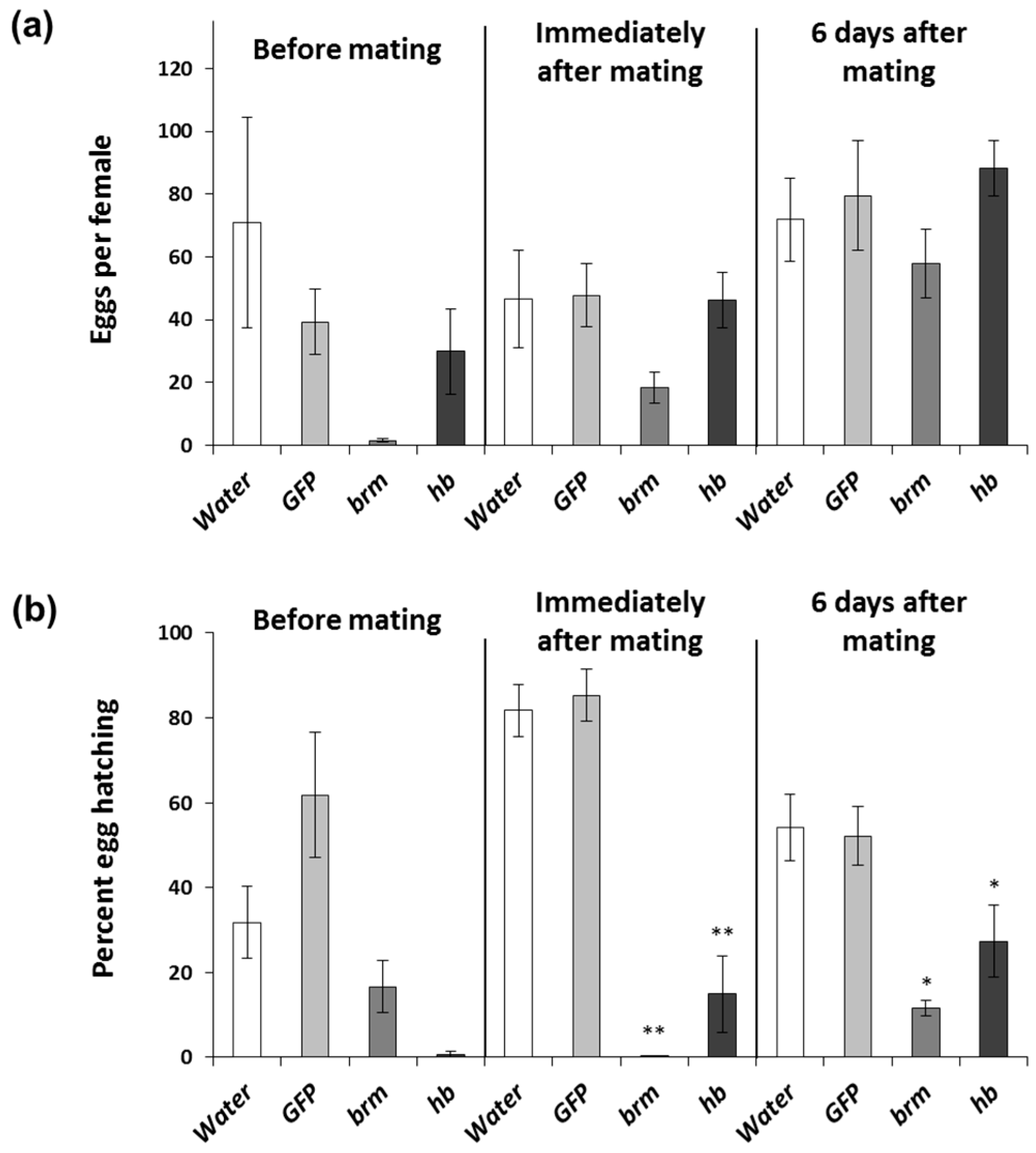
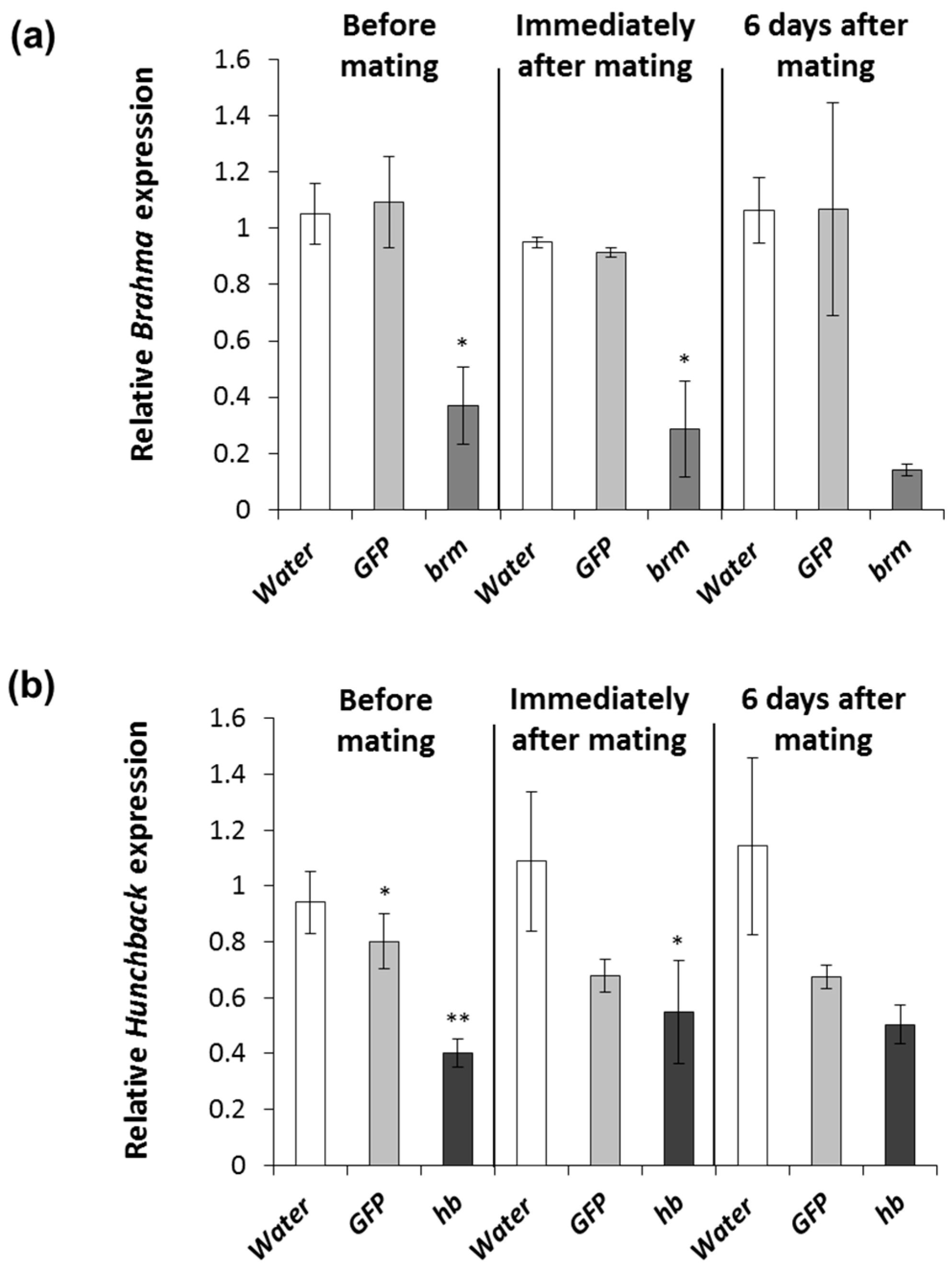
2.6. Timing of brahma and hunchback dsRNA Exposure with Respect to Mating Status
2.7. Effects of brahma and hunchback on Males
2.8. Quantitative Real-Time PCR (qRT-PCR)
2.9. Statistical Analysis
3. Results
3.1. brahma and hunchback Concentration Response
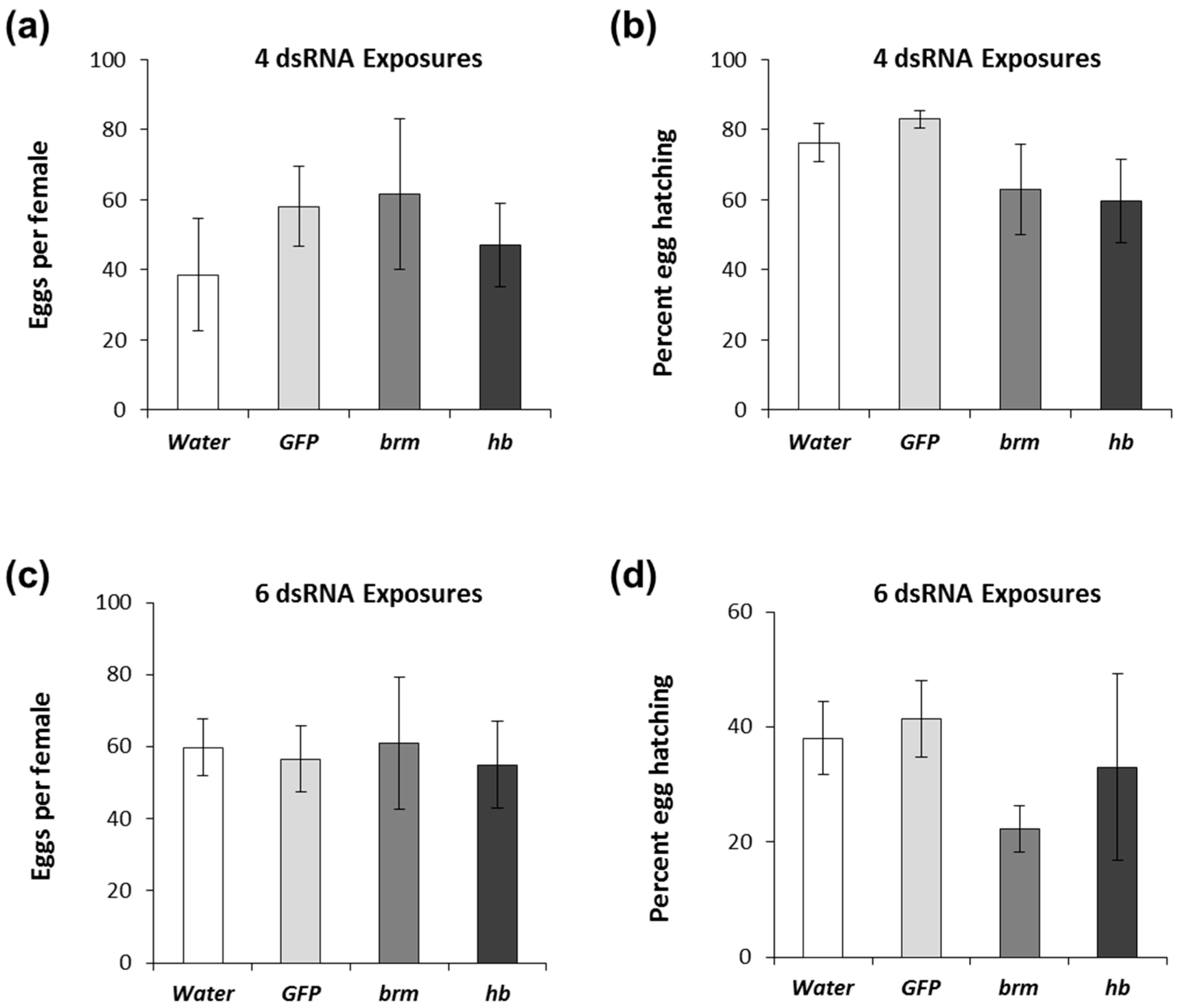
3.2. Duration of brahma and hunchback dsRNA Exposure
3.3. Timing of brahma and hunchback dsRNA Exposure with Respect to Mating Status
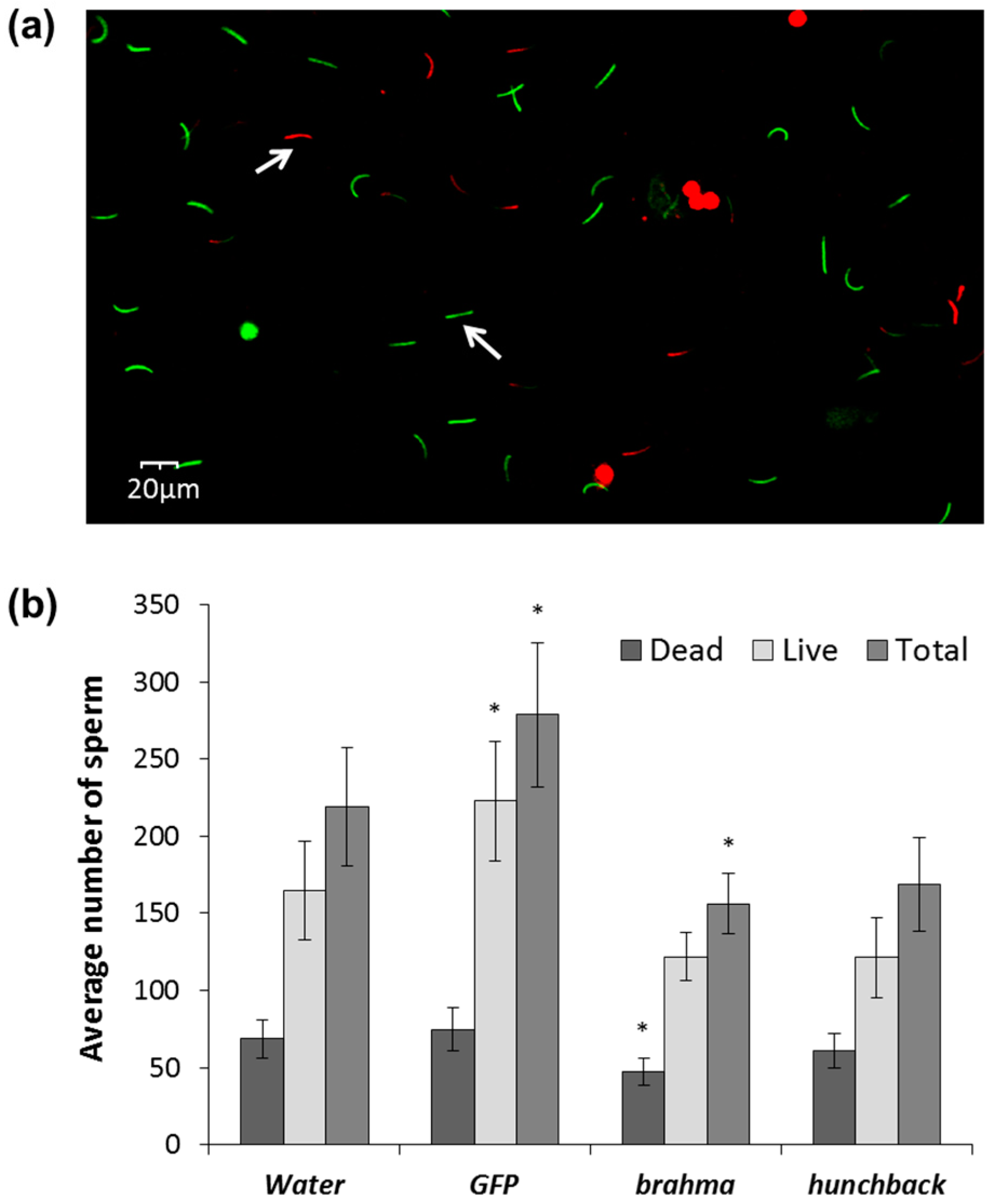
3.4. Effects of brahma and hunchback on Males
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chandler, L.D.; Coppedge, J.R.; Edwards, C.R.; Tollefson, J.J.; Wilde, G.R.; Faust, R.M. Corn rootworm areawide pest management in the midwestern USA. In Areawide Pest Management: Theory and Implementation; CABI Publishing: Wallingford, UK, 2008; pp. 191–207. [Google Scholar]
- Metcalf, R.L. Foreword. In Methods for the Study of Pest Diabrotica; Krysan, J.L., Miller, T.A, Eds.; Springer: New York, NY, USA, 1986. [Google Scholar]
- Gray, M.E.; Sappington, T.W.; Miller, N.J.; Moeser, J.; Bohn, M.O. Adaptation and invasiveness of western corn rootworm: Intensifying research on a worsening pest. Annu. Rev. Entomol. 2009, 54, 303–321. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.L.; Levine, E.; Isard, S.A.; Mabry, T.R. Movement, dispersal and behavior of western corn rootworm adults in rotated maize and soybean fields. In Western Corn Rootworm: Ecology and Management; Vidal, S., Kuhlmann, U., Edwards, C.R., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 121–144. [Google Scholar]
- Ma, B.L.; Meloche, F.; Wei, L. Agronomic assessment of Bt trait and seed or soil-applied insecticides on the control of corn rootworm and yield. Field Crops Res. 2009, 111, 189–196. [Google Scholar] [CrossRef]
- Marra, M.C.; Piggott, N.E.; Goodwin, B.K. The impact of corn rootworm protected biotechnology traits in the United States. AgBioForum 2012, 15, 217–230. [Google Scholar]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Ramaseshadri, P.; Anderson, J.; Bachman, P.; Clinton, W.; Flannagan, R.; Ilagan, O.; Lawrence, C.; Levine, S.; Moar, W.; et al. Characterizing the mechanism of action of double-stranded RNA activity against western corn rootworm (Diabrotica virgifera virgifera LeConte). PLoS ONE 2012, 7, e47534. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, M.; Siegfried, B.D. Validation of RNA interference in western corn rootworm Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae) adults. Pest Manag. Sci. 2012, 68, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Negri, I.; Oliveira, W.; Brown, C.; Asiimwe, P.; Sammons, B.; Horak, M.; Jiang, C.; Carson, D. Transportable data from non-target arthropod field studies for the environmental risk assessment of genetically modified maize expressing an insecticidal double-stranded RNA. Transgenic Res. 2016, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Petrick, J.S.; Frierdich, G.E.; Carleton, S.M.; Kessenich, C.R.; Silvanovich, A.; Zhang, Y.; Koch, M.S. Corn rootworm-active RNA DvSnf7: Repeat dose oral toxicology assessment in support of human and mammalian safety. Regula. Toxicol. Pharm. 2016. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.L.; Tan, J.; Mueller, G.M.; Bachman, P.M.; Jensen, P.D.; Uffman, J.P. Independent action between DvSnf7 RNA and Cry3Bb1 protein in southern corn rootworm, Diabrotica undecimpunctata howardi and Colorado potato beetle, Leptinotarsa decemlineata. PLoS ONE 2015, 10, e0118622. [Google Scholar] [CrossRef] [PubMed]
- Fishilevich, E.; Vélez, A.M.; Storer, N.P.; Li, H.; Bowling, A.J.; Rangasamy, M.; Worden, S.E.; Narva, K.E.; Siegfried, B.D. RNAi as a pest management tool for the western corn rootworm, Diabrotica virgifera virgifera. Pest Manag. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Bucher, G.; Scholten, J.; Klingler, M. Parental RNAi in Tribolium (Coleoptera). Curr. Biol. 2002, 12, R85–R86. [Google Scholar] [CrossRef]
- Shukla, J.N.; Palli, S.R. Production of all female progeny: Evidence for the presence of the male sex determination factor on the Y chromosome. J. Exp. Biol. 2014, 217, 1653–1655. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, C.; Vélez, A.M.; Rangasamy, M.; Wang, H.; Fishilevich, E.; Frey, M.L.F.; Carneiro, N.; Premchand, G.; Narva, K.E.; Siegfried, B.D. Parental RNA interference of genes involved in embryonic development of the western corn rootworm, Diabrotica virgifera virgifera LeConte. Insect Biochem. Mol. Biol. 2015, 63, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Konopova, B.; Akam, M. The hox genes ultrabithorax and abdominal-a specify three different types of abdominal appendage in the springtail Orchesella cincta (Collembola). Evodevo 2014. [Google Scholar] [CrossRef] [PubMed]
- Mito, T.; Okamoto, H.; Shinahara, W.; Shinmyo, Y.; Miyawaki, K.; Ohuchi, H.; Noji, S. Kruppel acts as a gap gene regulating expression of hunchback and even-skipped in the intermediate germ cricket Gryllus bimaculatus. Dev. Biol. 2006, 294, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Z.; Kaufman, T.C. Hunchback is required for suppression of abdominal identity, and for proper germband growth and segmentation in the intermediate germband insect Oncopeltus fasciatus. Development 2004, 131, 1515–1527. [Google Scholar] [CrossRef] [PubMed]
- Paim, R.M.; Araujo, R.N.; Lehane, M.J.; Gontijo, N.F.; Pereira, M.H. Long-term effects and parental RNAi in the blood feeder Rhodnius prolixus (Hemiptera: Reduviidae). Insect Biochem. Mol. Biol. 2013, 43, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.D.; Wouters, R.H.; Mugford, S.T.; Hogenhout, S.A. Persistence and transgenerational effect of plant-mediated RNAi in aphids. J. Exp. Bot. 2015, 66, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.J.; Liu, C.Y.; Zeng, F.R. Hunchback is required for abdominal identity supression and germband growth in the parthenogenetic embryogenesis of the pea aphis, Acyrthosiphon pisum. Arch. Insect Biochem. 2013, 84, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, N.; Tojo, K.; Hatakeyama, M. A survey of the effectiveness of non-cell autonomous RNAi throughout development in the sawfly, Athalia rosae (Hymenoptera). J. Insect Physiol. 2013, 59, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Piulachs, M.D.; Pagone, V.; Belles, X. Key roles of the broad-complex gene in insect embryogenesis. Insect Biochem. Mol. 2010, 40, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Nakao, H. Anterior and posterior centers jointly regulate Bombyx embryo body segmentation. Dev. Biol. 2012, 371, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Fishilevich, E.; Velez, A.M.; Khajuria, C.; Frey, M.L.; Hamm, R.L.; Wang, H.; Schulenberg, G.A.; Bowling, A.J.; Pence, H.E.; Gandra, P.; et al. Use of chromatin remodeling atpases as RNAi targets for parental control of western corn rootworm (Diabrotica virgifera virgifera) and neotropical brown stink bug (Euschistus heros). Insect Biochem. Mol. Biol. 2016, 71, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Miyata, K.; Ramaseshadri, P.; Zhang, Y.J.; Segers, G.; Bolognesi, R.; Tomoyasu, Y. Establishing an in vivo assay system to identify components involved in environmental RNA interference in the western corn rootworm. PLoS ONE 2014, 9, e101661. [Google Scholar] [CrossRef] [PubMed]
- Branson, T.F.; Krysan, J.L. Feeding and oviposition behavior and life cycle strategies of Diabrotica: An evolutionary view with implications for pest management (Coleoptera, Chrysomelidae). Environ. Entomol. 1981, 10, 826–831. [Google Scholar] [CrossRef]
- Meinke, L.J.; Sappington, T.W.; Onstad, D.W.; Guillemaud, T.; Miller, N.J.; Komáromi, J.; Levay, N.; Furlan, L.; Kiss, J.; Toth, F. Western corn rootworm (Diabrotica virgifera virgifera LeConte) population dynamics. Agric. Forest Entomol. 2009, 11, 29–46. [Google Scholar] [CrossRef]
- Spencer, J.L.; Hibbard, B.E.; Moeser, J.; Onstad, D.W. Behaviour and ecology of the western corn rootworm (Diabrotica virgifera virgifera LeConte). Agric. Forest Entomol. 2009, 11, 9–27. [Google Scholar] [CrossRef]
- Krysan, J.L. Introduction: Biology, distribution, and identification of pest Diabrotica. In Methods for the Study of Pest Diabrotica; Krysan, J.L., Miller, T.A., Eds.; Springer: New York, NY, USA, 1986; pp. 1–23. [Google Scholar]
- Oyediran, I.O.; Hibbard, B.E.; Clark, T.L. Prairie grasses as hosts of the western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2004, 33, 740–747. [Google Scholar] [CrossRef]
- Mabry, T.R.; Spencer, J.L.; Levine, E.; Isard, S.A. Western corn rootworm (Coleoptera: Chrysomelidae) behavior is affected by alternating diets of corn and soybean. Environ. Entomol. 2004, 33, 860–871. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Mullin, C.A. Effects of alternative host plants on longevity, oviposition, and emergence of western and northern corn rootworms (Coleoptera, Chrysomelidae). Environ. Entomol. 1990, 19, 474–480. [Google Scholar] [CrossRef]
- Shaw, J.T.; Paullus, J.H.; Luckmann, W.H. Corn rootworm (Coleoptera Chrysomelidae) oviposition in soybeans. J. Econ. Entomol. 1978, 71, 189–191. [Google Scholar] [CrossRef]
- Campbell, L.A.; Meinke, L.J. Seasonality and adult habitat use by four Diabrotica species at prairie-corn interfaces. Environ. Entomol. 2006, 35, 922–936. [Google Scholar] [CrossRef]
- Moeser, J.; Vidal, S. Nutritional resources used by the invasive maize pest Diabrotica virgifera virgifera in its new south-east-european distribution range. Entomol. Exp. Appl. 2005, 114, 55–63. [Google Scholar] [CrossRef]
- Branson, T.F. The contribution of prehatch and posthatch development to protandry in the Chrysomelid, Diabrotica virgifera virgifera. Entomol. Exp. Appl. 1987, 43, 205–208. [Google Scholar] [CrossRef]
- Guss, P.L. The sex pheromone of the western corn rootworm (Diabrotica virgifera). Environ. Entomol. 1976, 5, 219–223. [Google Scholar] [CrossRef]
- Hammack, L. Calling behavior in female western corn rootworm beetles (Coleoptera, Chrysomelidae). Ann. Entomol. Soc. Am. 1995, 88, 562–569. [Google Scholar] [CrossRef]
- Ball, H.J. On the biology and egg-laying habits of the western corn rootworm. J. Econ. Entomol. 1957, 50, 126–128. [Google Scholar] [CrossRef]
- Hill, R.E. Mating, oviposition patterns, fecundity and longevity of the western corn rootworm. J. Econ. Entomol. 1975, 68, 311–315. [Google Scholar] [CrossRef]
- Branson, T.F.; Guss, P.L.; Jackson, J.J. Mating frequency of the western corn rootworm. Ann. Entomol. Soc. Am. 1977, 70, 506–508. [Google Scholar] [CrossRef]
- Quiring, D.T.; Timmins, P.R. Influence of reproductive ecology on feasibility of mass trapping Diabrotica virgifera virgifera (Coleoptera, Chrysomelidae). J. Appl. Ecol. 1990, 27, 965–982. [Google Scholar] [CrossRef]
- Coats, S.A.; Tollefson, J.J.; Mutchmor, J.A. Study of migratory flight in the western corn rootworm (Coleoptera, Chrysomelidae). Environ. Entomol. 1986, 15, 620–625. [Google Scholar] [CrossRef]
- Naranjo, S.E. Movement of corn rootworm beetles, Diabrotica spp. (Coleoptera, Chrysomelidae), at cornfield boundaries in relation to sex, reproductive status, and crop phenology. Environ. Entomol. 1991, 20, 230–240. [Google Scholar] [CrossRef]
- Stebbing, J.A.; Meinke, L.J.; Naranjo, S.E.; Siegfried, B.D.; Wright, R.J.; Chandler, L.D. Flight behavior of methyl-parathion-resistant and -susceptible western corn rootworm (Coleoptera: Chrysomelidae) populations from Nebraska. J. Econ. Entomol. 2005, 98, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Darnell, S.J.; Meinke, L.T.; Young, L.J.; Gotway, C.A. Geostatistical investigation of the small-scale spatial variation of western corn rootworm (Coleoptera: Chrysomelidae) adults. Environ. Entomol. 1999, 28, 266–274. [Google Scholar] [CrossRef]
- Pierce, C.M.F.; Gray, M.E. Western corn rootworm, Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae), oviposition: A variant′s response to maize phenology. Environ. Entomol. 2006, 35, 423–434. [Google Scholar] [CrossRef]
- Cagan, L.; Rosca, I. Seasonal dispersal of the western corn rootworm (Diabrotica virgifera virgifera) adults in Bt and non-Bt maize fields. Plant Prot. Sci. 2012, 48, S36–S42. [Google Scholar]
- Steffey, K.L.; Tollefson, J.J. Spatial dispersion patterns of northern and western corn rootworm adults in Iowa cornfields. Environ. Entomol. 1982, 11, 283–286. [Google Scholar] [CrossRef]
- Eyun, S.I.; Wang, H.C.; Pauchet, Y.; Ffrench-Constant, R.H.; Benson, A.K.; Valencia-Jimenez, A.; Moriyama, E.N.; Siegfried, B.D. Molecular evolution of glycoside hydrolase genes in the western corn rootworm (Diabrotica virgifera virgifera). PLoS ONE 2014, 9, e94052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branson, T.F.; Jackson, J.J. An improved diet for adult Diabrotica virgifera vrgifera (Coleoptera, Chrysomelidae). J. Kansas Entomol. Soc. 1988, 61, 353–355. [Google Scholar]
- Campbell, L.A.; Meinke, L.J. Fitness of Diabrotica barberi, Diabrotica longicornis, and their hybrids (Coleoptera: Chrysomelidae). Ann. Entomol. Soc. Am. 2010, 103, 925–935. [Google Scholar] [CrossRef]
- Jackson, J.J. Rearing and handling of Diabrotica virgifera and Diabrotica undecimpunctata howardi. In Methods for the Study of Pest Diabrotica; Krysan, J.L., Miller, T.A., Eds.; Springer: New York, NY, USA, 1986; pp. 25–47. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Dahlgren, L.; Siegfried, B.D.; Ellis, M.D. Effect of in-hive miticides on drone honey bee survival and sperm viability. J. Apic. Res. 2013, 52, 88–95. [Google Scholar] [CrossRef]
- Rodrigues, T.B.; Khajuria, C.; Wang, H.; Matz, N.; Cunha Cardoso, D.; Valicente, F.H.; Zhou, X.; Siegfried, B. Validation of reference housekeeping genes for gene expression studies in western corn rootworm (Diabrotica virgifera virgifera). PLoS ONE 2014, 9, e109825. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- JMP®, version 11 pro; SAS Institute Inc.: Cary, NC, USA, 1989–2007.
- Wynant, N.; Verlinden, H.; Breugelmans, B.; Simonet, G.; Vanden Broeck, J. Tissue-dependence and sensitivity of the systemic rna interference response in the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2012, 42, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Vélez, A.M.; Khajuria, C.; Wang, H.; Narva, K.E.; Siegfried, B.D. Knockdown of RNA interference pathway genes in western corn rootworms (Diabrotica virgifera virgifera LeConte) demonstrates a possible mechanism of resistance to lethal dsRNA. PLoS ONE 2016, 11, e0157520. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bowling, A.J.; Gandra, P.; Rangasamy, M.; Pence, H.E.; McEwan, R.; Khajuria, C.; Siegfried, B.; Narva, K.E. Systemic RNAi in western corn rootworm, Diabrotica virgifera virgifera leconte, does not involve transitive pathways. Insect Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Betel, D.; Miller, M.L.; Sander, C.; Leslie, C.S.; Marks, D.S. Transfection of small RNAs globally perturbs gene regulation by endogenous microRNAs. Nat. Biotechnol. 2009, 27, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Brizuela, B.J.; Elfring, L.; Ballard, J.; Tamkun, J.W.; Kennison, J.A. Genetic analysis of the brahma gene of Drosophila melanogaster and polytene chromosome subdivisions 72ab. Genetics 1994, 137, 803–813. [Google Scholar] [PubMed]
- Deuring, R.; Fanti, L.; Armstrong, J.A.; Sarte, M.; Papoulas, O.; Prestel, M.; Daubresse, G.; Verardo, M.; Moseley, S.L.; Berloco, M.; et al. The ISWI chromatin-remodeling protein is required for gene expression and the maintenance of higher order chromatin structure in vivo. Mol. Cell 2000, 5, 355–365. [Google Scholar] [CrossRef]
- McDaniel, I.E.; Lee, J.M.; Berger, M.S.; Hanagami, C.K.; Armstrong, J.A. Investigations of CHD1 function in transcription and development of Drosophila melanogaster. Genetics 2008, 178, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Kehle, J.; Beuchle, D.; Treuheit, S.; Christen, B.; Kennison, J.A.; Bienz, M.; Muller, J. dMi-2, a hunchback-interacting protein that functions in polycomb repression. Science 1998, 282, 1897–1900. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xuan, T.; Xin, T.; An, H.; Wang, J.; Zhao, G.; Li, M. Evidence for chromatin-remodeling complex pbap-controlled maintenance of the Drosophila ovarian germline stem cells. PLoS ONE 2014, 9, e103473. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, P.T.; Krupke, C.H. Dispersal and mating behavior of Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae) in Bt cornfields. Environ. Entomol. 2009, 38, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Elliott, N.C.; Gustin, R.D.; Hanson, S.L. Influence of adult diet on the reproductive biology and survival of the western corn rootworm, Diabrotica virgifera virgifera. Entomol. Exp. Appl. 1990, 56, 15–21. [Google Scholar] [CrossRef]
- Isard, S.A.; Spencer, J.L.; Mabry, T.R.; Levine, E. Influence of atmospheric conditions on high-elevation flight of western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2004, 33, 650–656. [Google Scholar] [CrossRef]
- Hibbard, B.E.; Higdon, M.L.; Duran, D.P.; Schweikert, Y.M.; Ellersieck, M.R. Role of egg density on establishment and plant-to-plant movement by western corn rootworm larvae (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2004, 97, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.R.; Sutter, G.R.; Branson, T.F. Influence of corn planting date on the survival and on some reproductive parameters of Diabrotica virgifera virgifera (Coleoptera, Chrysomelidae). Environ. Entomol. 1991, 20, 185–189. [Google Scholar] [CrossRef]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vélez, A.M.; Fishilevich, E.; Matz, N.; Storer, N.P.; Narva, K.E.; Siegfried, B.D. Parameters for Successful Parental RNAi as An Insect Pest Management Tool in Western Corn Rootworm, Diabrotica virgifera virgifera. Genes 2017, 8, 7. https://doi.org/10.3390/genes8010007
Vélez AM, Fishilevich E, Matz N, Storer NP, Narva KE, Siegfried BD. Parameters for Successful Parental RNAi as An Insect Pest Management Tool in Western Corn Rootworm, Diabrotica virgifera virgifera. Genes. 2017; 8(1):7. https://doi.org/10.3390/genes8010007
Chicago/Turabian StyleVélez, Ana M., Elane Fishilevich, Natalie Matz, Nicholas P. Storer, Kenneth E. Narva, and Blair D. Siegfried. 2017. "Parameters for Successful Parental RNAi as An Insect Pest Management Tool in Western Corn Rootworm, Diabrotica virgifera virgifera" Genes 8, no. 1: 7. https://doi.org/10.3390/genes8010007
APA StyleVélez, A. M., Fishilevich, E., Matz, N., Storer, N. P., Narva, K. E., & Siegfried, B. D. (2017). Parameters for Successful Parental RNAi as An Insect Pest Management Tool in Western Corn Rootworm, Diabrotica virgifera virgifera. Genes, 8(1), 7. https://doi.org/10.3390/genes8010007