
Functional Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in the Human RORγT Gene Promoter Region


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RORγT Promoter Constructs and Transfection
2.3. Site-Directed Mutagenesis
2.4. Electrophoretic Mobility Shift Assays
2.5. Material and Sample Preparation
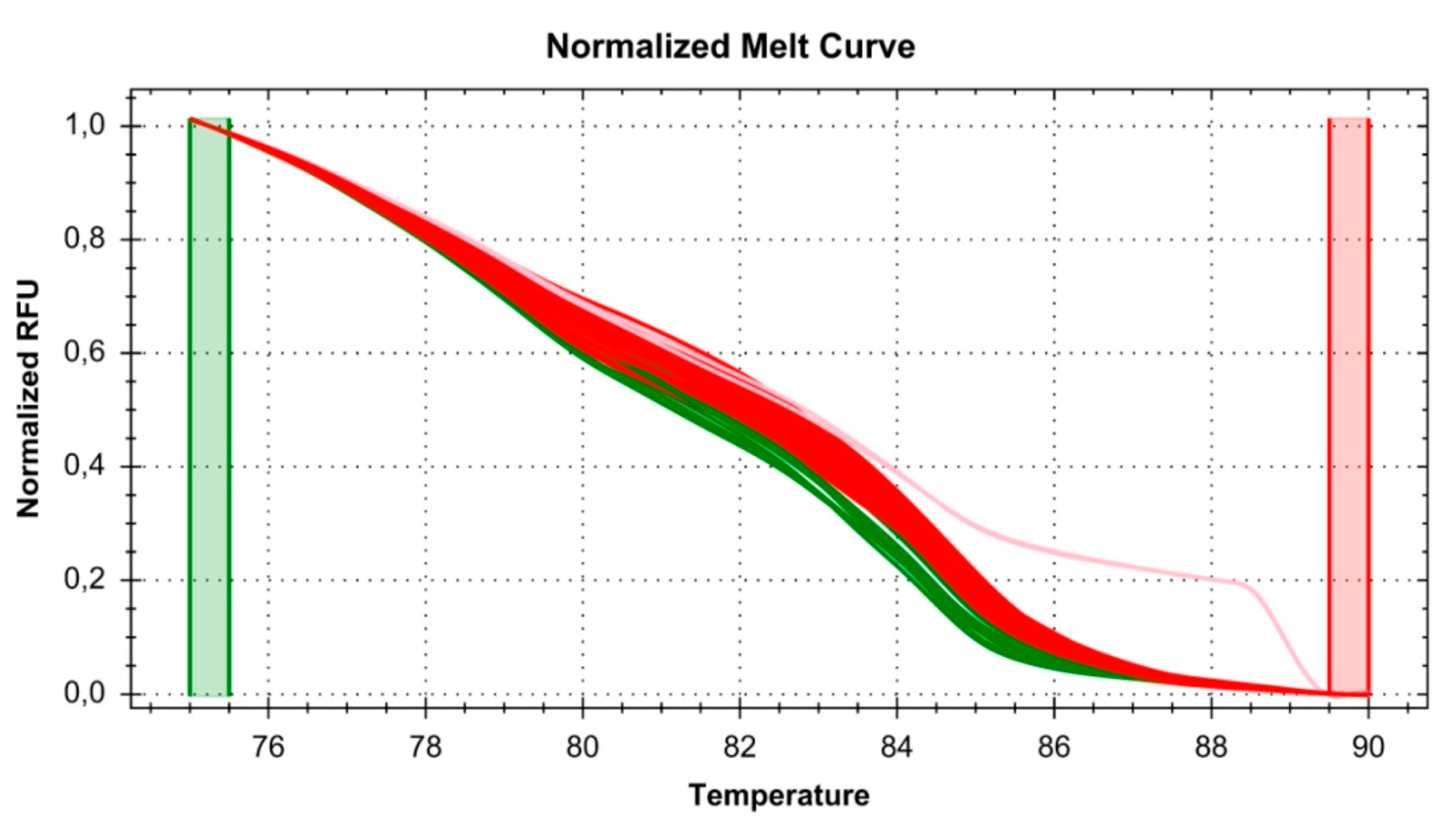
2.6. High Resolution Melt (HRM) Conditions and Analysis
2.7. Detection of Genetic Variation
2.8. Computational Analysis and Statistics
3. Results
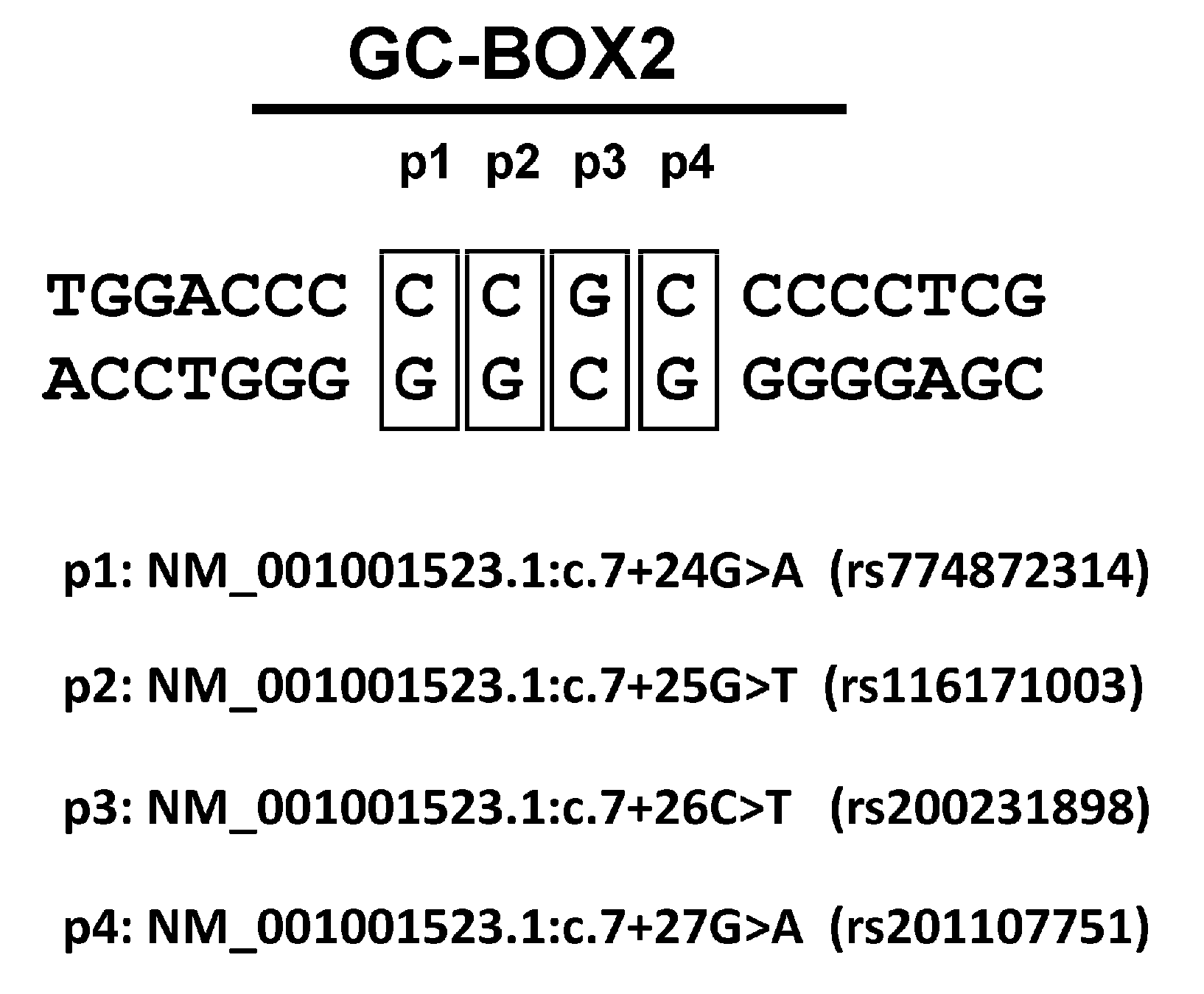
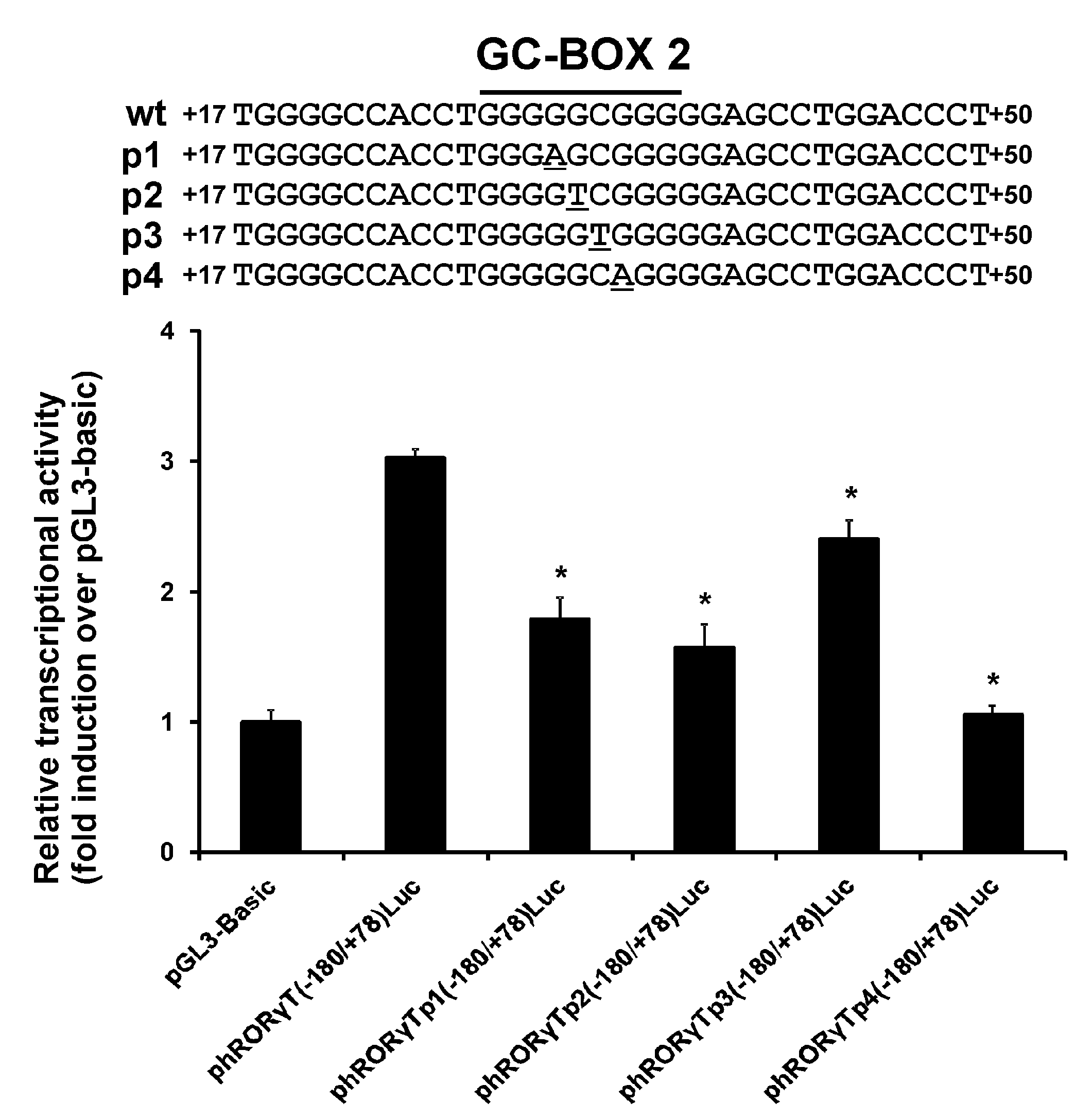
3.1. Effect of the rs774872314, rs116171003, rs200231898 and rs201107751 SNPs on the Activity of the RORγT Promoter
3.2. Protein Binding to the Sequences Corresponding to the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms
3.3. Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in a Polish Population
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Medvedev, A.; Chistokhina, A.; Hirose, T.; Jetten, A.M. Genomic structure and chromosomal mapping of the nuclear orphan receptor RORγ (RORC) gene. Genomics 1997, 46, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Villey, I.; de Chasseval, R.; de Villartay, J.P. RORγT, a thymus-specific isoform of the orphan nuclear receptor RORγ/TOR, is up-regulated by signaling through the pre-T cell receptor and binds to the tea promoter. Eur. J. Immunol. 1999, 29, 4072–4080. [Google Scholar] [CrossRef]
- Ratajewski, M.; Walczak-Drzewiecka, A.; Salkowska, A.; Dastych, J. Upstream stimulating factors regulate the expression of RORγT in human lymphocytes. J. Immunol. 2012, 189, 3034–3042. [Google Scholar] [CrossRef] [PubMed]
- Jetten, A.M. Retinoid-related orphan receptors (RORs): Critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl. Recept. Signal. 2009, 7, e003. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.M.; Ramachandran, G.; Basu, S.; Rollins, S.; Mann, D.; Cross, A.S. The IL-23/Th17 axis is involved in the adaptive immune response to Bacillus anthracis in humans. Eur. J. Immunol. 2014, 44, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ibrahim, A.S.; Xu, X.; Farber, J.M.; Avanesian, V.; Baquir, B.; Fu, Y.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. Th1-Th17 cells mediate protective adaptive immunity against Staphylococcus aureus and Candida albicans infection in mice. PLoSPathog. 2009, 5, e1000703. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Na, L.; Fidel, P.L.; Schwarzenberger, P. Requirement of interleukin-17a for systemic anti-Candida albicans host defense in mice. J. Infect. Dis. 2004, 190, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Kebir, H.; Kreymborg, K.; Ifergan, I.; Dodelet-Devillers, A.; Cayrol, R.; Bernard, M.; Giuliani, F.; Arbour, N.; Becher, B.; Prat, A. Human Th17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation. Nat. Med. 2007, 13, 1173–1175. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Hashimoto, M.; Yoshitomi, H.; Tanaka, S.; Nomura, T.; Yamaguchi, T.; Iwakura, Y.; Sakaguchi, N.; Sakaguchi, S. T cell self-reactivity forms a cytokine milieu for spontaneous development of IL-17+Th cells that cause autoimmune arthritis. J. Exp. Med. 2007, 204, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, X.; Liu, Z.; Yue, Q.; Liu, H. Expression of Th17 cytokines in skin lesions of patients with psoriasis. J. HuazhongUniv. Sci. Technol. Med. Sci. 2007, 27, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Ye, P.; Liu, C. The role of the IL-23/IL-17 axis in the pathogenesis of graves’ disease. Endocr. J. 2013, 60, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Schmidl, C.; Hansmann, L.; Andreesen, R.; Edinger, M.; Hoffmann, P.; Rehli, M. Epigenetic reprogramming of the RORC locus during in vitro expansion is a distinctive feature of human memory but not naive Treg. Eur. J. Immunol. 2011, 41, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Ratajewski, M.; Walczak-Drzewiecka, A.; Gorzkiewicz, M.; Salkowska, A.; Dastych, J. Expression of human gene coding RORγT receptor depends on the Sp2 transcription factor. J. Leukoc. Biol. 2016, 100, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2015, 43. [Google Scholar] [CrossRef]
- Strapagiel, D.; Sobalska-Kwapis, M.; Slomka, M.; Marciniak, B. Biobank Lodz—DNA based biobank at the University of Lodz, Poland. Open J. Bioresour. 2016, 3, e6. [Google Scholar] [CrossRef]
- Strapagiel, D.; Majewska, M.; Słomka, M.; Janik, K.; Sobalska, M.; Bartosz, G. Method for Determining sex, Involves Utilizing Melting Profile Analysis Technique, and Obtaining Specific Fragments of DNA by PCR-Based DNA from Samples of Human Biological Material; Biobank Lab: Lodz, Poland, 2016. [Google Scholar]
- Slomka, M.; Sobalska-Kwapis, M.; Korycka-Machala, M.; Bartosz, G.; Dziadek, J.; Strapagiel, D. Genetic variation of the ABC transporter gene ABCC1 (multidrug resistance protein 1-MRP1) in the Polish population. BMC Genet. 2015, 16, 114. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. Matinspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 2005, 21, 2933–2942. [Google Scholar] [CrossRef] [PubMed]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Sudmant, P.H.; Rausch, T.; Gardner, E.J.; Handsaker, R.E.; Abyzov, A.; Huddleston, J.; Zhang, Y.; Ye, K.; Jun, G.; Hsi-Yang Fritz, M.; et al. An integrated map of structural variation in 2,504 human genomes. Nature 2015, 526, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Exome Variant Server, NHLBI GO Exome Sequencing Project (ESP), Seattle, WA, USA. Available online: http://evs.gs.washington.edu/EVS/ (accessed on 20 March 2017).
- Hoppe, K.L.; Francone, O.L. Binding and functional effects of transcription factors Sp1 and Sp3 on the proximal human lecithin: Cholesterol acyltransferase promoter. J. Lipid Res. 1998, 39, 969–977. [Google Scholar] [PubMed]
- Moorefield, K.S.; Fry, S.J.; Horowitz, J.M. Sp2 DNA binding activity and trans-activation are negatively regulated in mammalian cells. J. Biol. Chem. 2004, 279, 13911–13924. [Google Scholar] [CrossRef] [PubMed]
- Courey, A.J.; Holtzman, D.A.; Jackson, S.P.; Tjian, R. Synergistic activation by the glutamine-rich domains of human transcription factor Sp1. Cell 1989, 59, 827–836. [Google Scholar] [CrossRef]
- Pascal, E.; Tjian, R. Different activation domains of Sp1 govern formation of multimers and mediate transcriptional synergism. Genes Dev. 1991, 5, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Emili, A.; Greenblatt, J.; Ingles, C.J. Species-specific interaction of the glutamine-rich activation domains of Sp1 with the TATA box-binding protein. Mol. Cell. Biol. 1994, 14, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Torigoe, T.; Izumi, H.; Yoshida, Y.; Ishiguchi, H.; Okamoto, T.; Itoh, H.; Kohno, K. Low pH enhances Sp1 DNA binding activity and interaction with TBP. Nucleic Acids Res. 2003, 31, 4523–4530. [Google Scholar] [CrossRef] [PubMed]
- Hoey, T.; Weinzierl, R.O.; Gill, G.; Chen, J.L.; Dynlacht, B.D.; Tjian, R. Molecular cloning and functional analysis of drosophila TAF110 reveal properties expected of coactivators. Cell 1993, 72, 247–260. [Google Scholar] [CrossRef]
- Crome, S.Q.; Wang, A.Y.; Kang, C.Y.; Levings, M.K. The role of retinoic acid-related orphan receptor variant 2 and IL-17 in the development and function of human CD4+ T cells. Eur. J. Immunol. 2009, 39, 1480–1493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Meng, G.; Strober, W. Interactions among the transcription factors Runx1, RORγT and Foxp3 regulate the differentiation of interleukin 17-producing T cells. Nat. Immunol. 2008, 9, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.O.; Pappu, B.P.; Nurieva, R.; Akimzhanov, A.; Kang, H.S.; Chung, Y.; Ma, L.; Shah, B.; Panopoulos, A.D.; Schluns, K.S.; et al. T helper 17 lineage differentiation is programmed by orphan nuclear receptors RORα and RORγ. Immunity 2008, 28, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Onishi, R.M.; Gaffen, S.L. Interleukin-17 and its target genes: Mechanisms of interleukin-17 function in disease. Immunology 2010, 129, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Kabata, Y.; Shimada, M.K.; Hayakawa, Y.; Minoshima, S.; Chakraborty, R.; Gojobori, T.; Imanishi, T. Distribution and effects of nonsense polymorphisms in human genes. PLoS ONE 2008, 3, e3393. [Google Scholar] [CrossRef] [PubMed]
- Dunna, N.R.; Vuree, S.; Kagita, S.; Surekha, D.; Digumarti, R.; Rajappa, S.; Satti, V. Association of GSTP1 gene (I105V) polymorphism with acute Leukaemia. J. Genet. 2012, 91, e60–e63. [Google Scholar] [CrossRef] [PubMed]
- Pastinen, T.; Ge, B.; Hudson, T.J. Influence of human genome polymorphism on gene expression. Hum. Mol. Genet. 2006, 15, R9–R16. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.S.; Chew, G.Y.; Simpson, N.; Priyadarshi, A.; Wong, M.; Grimbacher, B.; Fulcher, D.A.; Tangye, S.G.; Cook, M.C. Deficiency of Th17 cells in hyper IgE syndrome due to mutations in stat3. J. Exp. Med. 2008, 205, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Eyerich, K.; Foerster, S.; Rombold, S.; Seidl, H.P.; Behrendt, H.; Hofmann, H.; Ring, J.; Traidl-Hoffmann, C. Patients with chronic mucocutaneous candidiasis exhibit reduced production of Th17-associated cytokines IL-17 and IL-22. J. Investig. Dermatol. 2008, 128, 2640–2645. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Meng, S.; Song, R.H.; Qin, Q.; Wang, X.; Yao, Q.; Jiang, Y.; Jiang, W.; Shi, L.; Xu, J.; et al. Polymorphism of IL37 gene as a protective factor for autoimmune thyroid disease. J. Mol. Endocrinol. 2015, 55, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, B.; Tang, K.; Lee, E.J.; Chong, S.S.; Lee, C.G. A functional polymorphism within the MRP1 gene locus identified through its genomic signature of positive selection. Hum. Mol. Genet. 2005, 14, 2075–2087. [Google Scholar] [CrossRef] [PubMed]
- Umamaheswaran, G.; Kumar, D.K.; Adithan, C. Distribution of genetic polymorphisms of genes encoding drug metabolizing enzymes & drug transporters—A review with Indian perspective. Indian J. Med. Res. 2014, 139, 27–65. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Genotype | N | % | MAF |
|---|---|---|---|---|
| NM_001001523.1:c.7+25G>T (rs116171003) | GG | 4878 | 96.575 | (T) 0.017 |
| GT | 171 | 3.385 | ||
| TT | 2 | 0.040 | ||
| NM_001001523.1:c.7+34C>G (rs111882199) | CC | 4993 | 98.852 | (G) 0.006 |
| CG | 57 | 1.128 | ||
| GG | 1 | 0.020 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratajewski, M.; Słomka, M.; Karaś, K.; Sobalska-Kwapis, M.; Korycka-Machała, M.; Sałkowska, A.; Dziadek, J.; Strapagiel, D.; Dastych, J. Functional Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in the Human RORγT Gene Promoter Region. Genes 2017, 8, 126. https://doi.org/10.3390/genes8040126
Ratajewski M, Słomka M, Karaś K, Sobalska-Kwapis M, Korycka-Machała M, Sałkowska A, Dziadek J, Strapagiel D, Dastych J. Functional Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in the Human RORγT Gene Promoter Region. Genes. 2017; 8(4):126. https://doi.org/10.3390/genes8040126
Chicago/Turabian StyleRatajewski, Marcin, Marcin Słomka, Kaja Karaś, Marta Sobalska-Kwapis, Małgorzata Korycka-Machała, Anna Sałkowska, Jarosław Dziadek, Dominik Strapagiel, and Jarosław Dastych. 2017. "Functional Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in the Human RORγT Gene Promoter Region" Genes 8, no. 4: 126. https://doi.org/10.3390/genes8040126
APA StyleRatajewski, M., Słomka, M., Karaś, K., Sobalska-Kwapis, M., Korycka-Machała, M., Sałkowska, A., Dziadek, J., Strapagiel, D., & Dastych, J. (2017). Functional Analysis of the rs774872314, rs116171003, rs200231898 and rs201107751 Polymorphisms in the Human RORγT Gene Promoter Region. Genes, 8(4), 126. https://doi.org/10.3390/genes8040126