
Endogenous Retroviral Insertions Indicate a Secondary Introduction of Domestic Sheep Lineages to the Caucasus and Central Asia between the Bronze and Iron Age

 ,
, 
{kind=link}
{kind=link}
Abstract
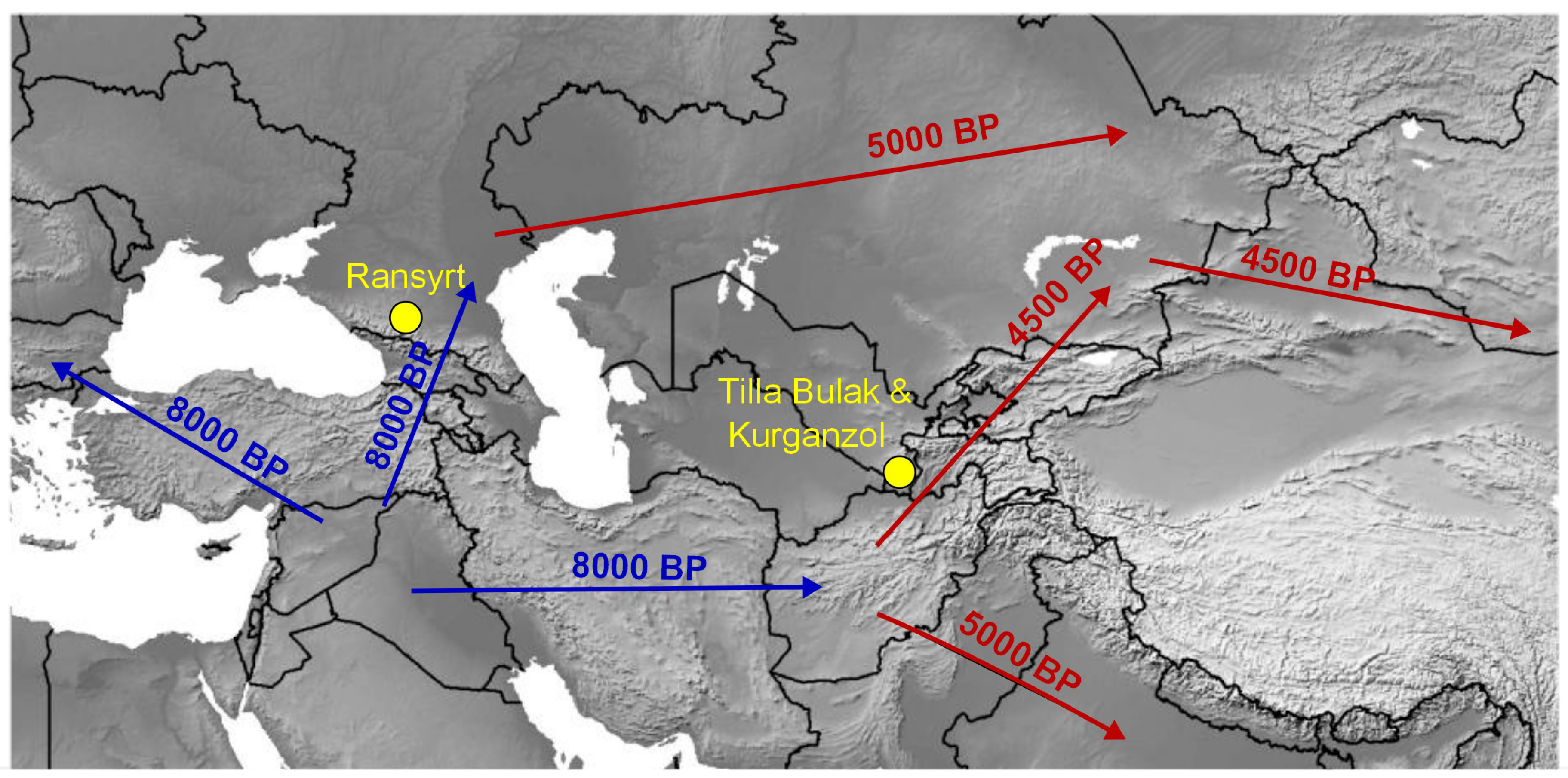
:1. Introduction
2. Materials and Methods
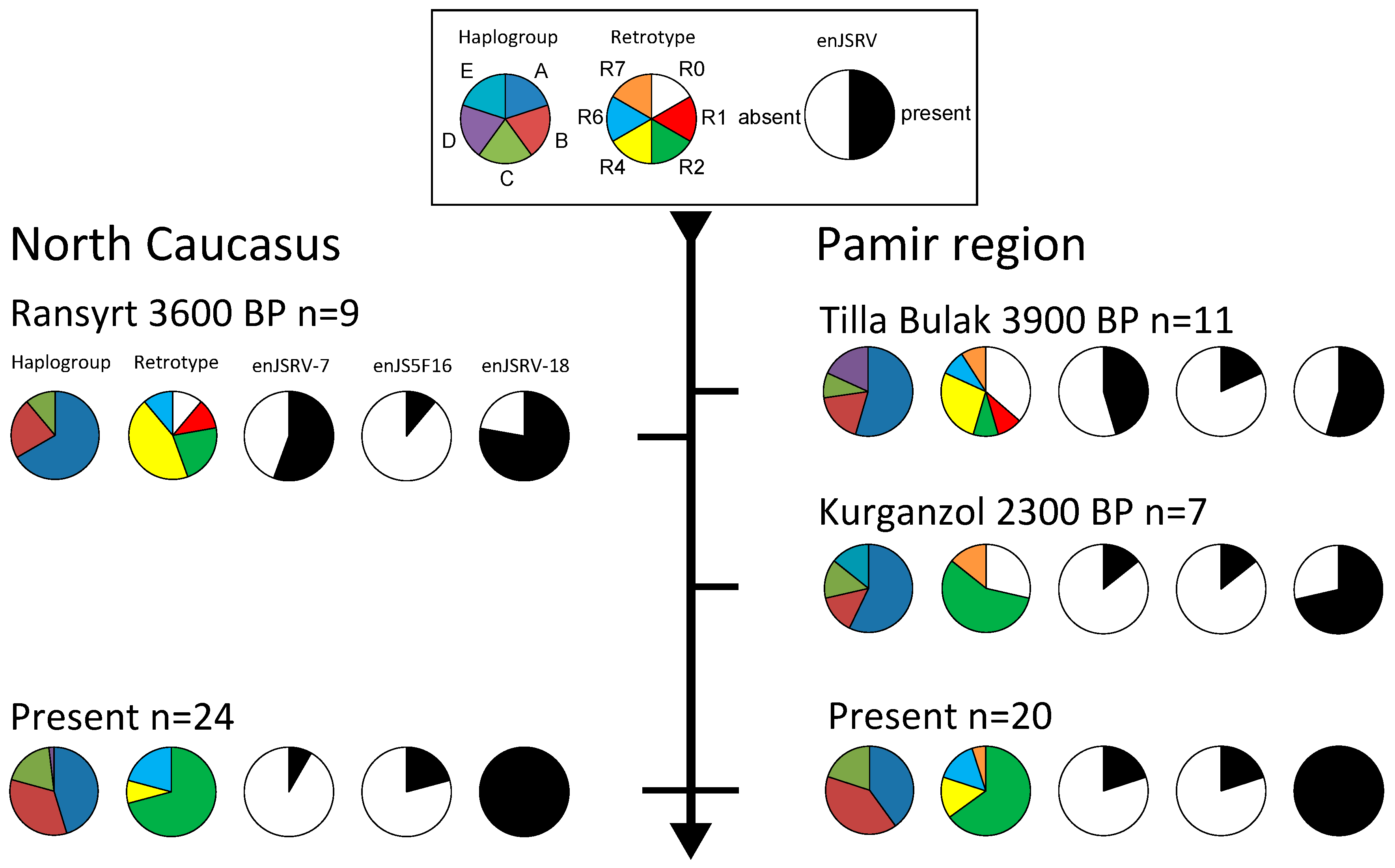
3. Results
4. Discussion
4.1. Mitochondrial Haplogroups
4.2. Retrotypes
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zeder, M.A. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proc. Natl. Acad. Sci. USA 2008, 105, 11597–11604. [Google Scholar] [CrossRef] [PubMed]
- Anthony, D.W. The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World, Reprint ed.; Princeton University Press: Princeton, NJ, USA; Woodstock, GA, USA, 2007. [Google Scholar]
- Frachetti, M.D. Multiregional Emergence of Mobile Pastoralism and Nonuniform Institutional Complexity across Eurasia. Curr. Anthropol. 2012, 53, 2–38. [Google Scholar] [CrossRef]
- Geörg, C. Paläopopulationsgenetik von Schwein und Schaf in Südosteuropa und Transkaukasien; VML: Stellerloh, Germany, 2013. [Google Scholar]
- Chataigner, C.; Badalyan, R.; Arimura, M. The Neolithic of the Caucasus. Oxford Handbooks Online 2014. [Google Scholar] [CrossRef]
- Frachetti, M.; Benecke, N. From sheep to (some) horses: 4500 years of herd structure at the pastoralist settlement of Begash (south-eastern Kazakhstan). Antiquity 2009, 83, 1023–1037. [Google Scholar] [CrossRef]
- Betts, A.; Jia, P.W.; Dodson, J. The origins of wheat in China and potential pathways for its introduction: A review. Quat. Int. 2014, 348, 158–168. [Google Scholar] [CrossRef]
- Spengler, R.N. Agriculture in the Central Asian Bronze Age. J. World Prehist. 2015, 28, 215–253. [Google Scholar] [CrossRef]
- Spengler, R.N.; Ryabogina, N.; Tarasov, P.E.; Wagner, M. The spread of agriculture into northern Central Asia: Timing, pathways, and environmental feedbacks. Holocene 2016, 26, 1527–1540. [Google Scholar] [CrossRef]
- Zhao, Z. Eastward Spread of Wheat into China—New Data and New Issues. Chin. Archaeol. 2009, 9, 1–9. [Google Scholar] [CrossRef]
- Cai, D.-W.; Han, L.; Zhang, X.-L.; Zhou, H.; Zhu, H. DNA analysis of archaeological sheep remains from China. J. Archaeol. Sci. 2007, 34, 1347–1355. [Google Scholar] [CrossRef]
- Cai, D.; Tang, Z.; Yu, H.; Han, L.; Ren, X.; Zhao, X.; Zhu, H.; Zhou, H. Early history of Chinese domestic sheep indicated by ancient DNA analysis of Bronze Age individuals. J. Archaeol. Sci. 2011, 38, 896–902. [Google Scholar] [CrossRef]
- O’Sullivan, N.J.; Teasdale, M.D.; Mattiangeli, V.; Maixner, F.; Pinhasi, R.; Bradley, D.G.; Zink, A. A whole mitochondria analysis of the Tyrolean Iceman’s leather provides insights into the animal sources of Copper Age clothing. Sci. Rep. 2016, 6, 31279. [Google Scholar] [CrossRef] [PubMed]
- Schröder, O.; Wagner, M.; Wutke, S.; Zhang, Y.; Ma, Y.; Xu, D.; Goslar, T.; Neef, R.; Tarasov, P.E.; Ludwig, A. Ancient DNA identification of domestic animals used for leather objects in Central Asia during the Bronze Age. Holocene 2016, 26, 1722–1729. [Google Scholar] [CrossRef]
- Hiendleder, S.; Mainz, K.; Plante, Y.; Lewalski, H. Analysis of mitochondrial DNA indicates that domestic sheep are derived from two different ancestral maternal sources: No evidence for contributions from urial and argali sheep. J. Hered. 1998, 89, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep Mitochondrial DNA Variation in European, Caucasian, and Central Asian Areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, Y.H.; Chen, H.; Guan, W.J. Genetic and phylogenetic studies of Chinese native sheep breeds (Ovis aries) based on mtDNA D-loop sequences. Small Rumin. Res. 2007, 72, 232–236. [Google Scholar] [CrossRef]
- Meadows, J.R.S.; Hiendleder, S.; Kijas, J.W. Haplogroup relationships between domestic and wild sheep resolved using a mitogenome panel. Heredity 2011, 106, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.; Yu, Q.; Zhang, N.; Kong, D.; Zhao, Y. Mitochondrial DNA diversity and the origin of Chinese indigenous sheep. Trop. Anim. Health Prod. 2013, 45, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.-H.; Peng, W.-F.; Yang, J.; Zhao, Y.-X.; Li, W.-R.; Liu, M.-J.; Ma, Y.-H.; Zhao, Q.-J.; Yang, G.-L.; Wang, F.; et al. Mitogenomic meta-analysis identifies two phases of migration in the history of eastern Eurasian sheep. Mol. Biol. Evol. 2015, 32, 2515–2533. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Pichler, R.; Babar, M.E.; Khan, W.A.; Ullah, Z.; Shehzad, S.; Periasamy, K. Mitochondrial DNA D-Loop diversity and evolutionary relationship of wild Punjab Urial sheep (Ovis vignei punjabiensis) with closely related taxa. Small Rumin. Res. 2016. [Google Scholar] [CrossRef]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the History of Sheep Domestication Using Retrovirus Integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, S. Spätbronzezeitliche Fundplätze im kaukasischen Hochgebirge. e-Forschungsberichte DAI 2014 2014, 2, 82–88. [Google Scholar]
- Kaniuth, K. The Late Bronze Age Settlement of Tilla Bulak (Southern Uzbekistan). A Summary of Four Year’s Work. In South Asian Archaeology and Art 2012. Vol. I. Man and Environment in Prehistoric South Asia: New Perspectives; Lefèvre, V., Didier, A., Mutin, B., Eds.; Indicopleustoi; Brepols: Turnhout, Belgium, 2016; Volume 12, pp. 117–127. [Google Scholar]
- Sverchkov, L. The Kurganzol Fortress (on the History of Central Asia in the Hellenistic Era). Anc. Civ. Scythia Siberia 2008, 14, 123–191. [Google Scholar] [CrossRef]
- Cooper, A.; Poinar, H.N. Ancient DNA: Do it right or not at all. Science 2000, 289, 1139. [Google Scholar] [CrossRef] [PubMed]
- Dabney, J.; Knapp, M.; Glocke, I.; Gansauge, M.-T.; Weihmann, A.; Nickel, B.; Valdiosera, C.; García, N.; Pääbo, S.; Arsuaga, J.-L.; et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. USA 2013, 110, 15758–15763. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.-S.; Fan, L.; Shi, N.-N.; Ning, T.; Yao, Y.-G.; Murphy, R.W.; Wang, W.-Z.; Zhang, Y.-P. DomeTree: A canonical toolkit for mitochondrial DNA analyses in domesticated animals. Mol. Ecol. Resour. 2015, 15, 1238–1242. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, F.; Caporale, M.; Varela, M.; Biek, R.; Chessa, B.; Alberti, A.; Golder, M.; Mura, M.; Zhang, Y.; Yu, L.; et al. A Paradigm for Virus–Host Coevolution: Sequential Counter-Adaptations between Endogenous and Exogenous Retroviruses. PLoS Pathog. 2007, 3, e170. [Google Scholar] [CrossRef] [PubMed]
- Schröder, O.; Lieckfeldt, D.; Lutz, W.; Rudloff, C.; Frölich, K.; Ludwig, A. Limited hybridization between domestic sheep and the European mouflon in Western Germany. Eur. J. Wildl. Res. 2016, 62, 307–314. [Google Scholar] [CrossRef]
- Bowles, D.; Carson, A.; Isaac, P. Genetic Distinctiveness of the Herdwick Sheep Breed and Two Other Locally Adapted Hill Breeds of the UK. PLoS ONE 2014, 9, e87823. [Google Scholar] [CrossRef] [PubMed]
- Rannamäe, E.; Lõugas, L.; Speller, C.F.; Valk, H.; Maldre, L.; Wilczyński, J.; Mikhailov, A.; Saarma, U. Three Thousand Years of Continuity in the Maternal Lineages of Ancient Sheep (Ovis aries) in Estonia. PLoS ONE 2016, 11, e0163676. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeder, O.; Benecke, N.; Frölich, K.; Peng, Z.; Kaniuth, K.; Sverchkov, L.; Reinhold, S.; Belinskiy, A.; Ludwig, A. Endogenous Retroviral Insertions Indicate a Secondary Introduction of Domestic Sheep Lineages to the Caucasus and Central Asia between the Bronze and Iron Age. Genes 2017, 8, 165. https://doi.org/10.3390/genes8060165
Schroeder O, Benecke N, Frölich K, Peng Z, Kaniuth K, Sverchkov L, Reinhold S, Belinskiy A, Ludwig A. Endogenous Retroviral Insertions Indicate a Secondary Introduction of Domestic Sheep Lineages to the Caucasus and Central Asia between the Bronze and Iron Age. Genes. 2017; 8(6):165. https://doi.org/10.3390/genes8060165
Chicago/Turabian StyleSchroeder, Oskar, Norbert Benecke, Kai Frölich, Zuogang Peng, Kai Kaniuth, Leonid Sverchkov, Sabine Reinhold, Andrey Belinskiy, and Arne Ludwig. 2017. "Endogenous Retroviral Insertions Indicate a Secondary Introduction of Domestic Sheep Lineages to the Caucasus and Central Asia between the Bronze and Iron Age" Genes 8, no. 6: 165. https://doi.org/10.3390/genes8060165
APA StyleSchroeder, O., Benecke, N., Frölich, K., Peng, Z., Kaniuth, K., Sverchkov, L., Reinhold, S., Belinskiy, A., & Ludwig, A. (2017). Endogenous Retroviral Insertions Indicate a Secondary Introduction of Domestic Sheep Lineages to the Caucasus and Central Asia between the Bronze and Iron Age. Genes, 8(6), 165. https://doi.org/10.3390/genes8060165