Functional Characterization of Selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of USP Genes from the Salvia miltiorrhiza Genomic Database
2.2. Bioinformatics Analysis, Phylogenetic Analysis, and Multiple Sequence Alignment
2.3. Plant Growth and Stress Treatments
2.4. Promoter Sequence Analysis, RNA Isolation, and Quantitative RT-PCR Analysis
2.5. Molecular Cloning and Construction of Escherichia coli Strains Expressing Universal Stress Proteins
2.6. Salt, Heat, and Combined Treatments with Transformed E. coli Cells
2.7. Statistical Analysis
3. Results
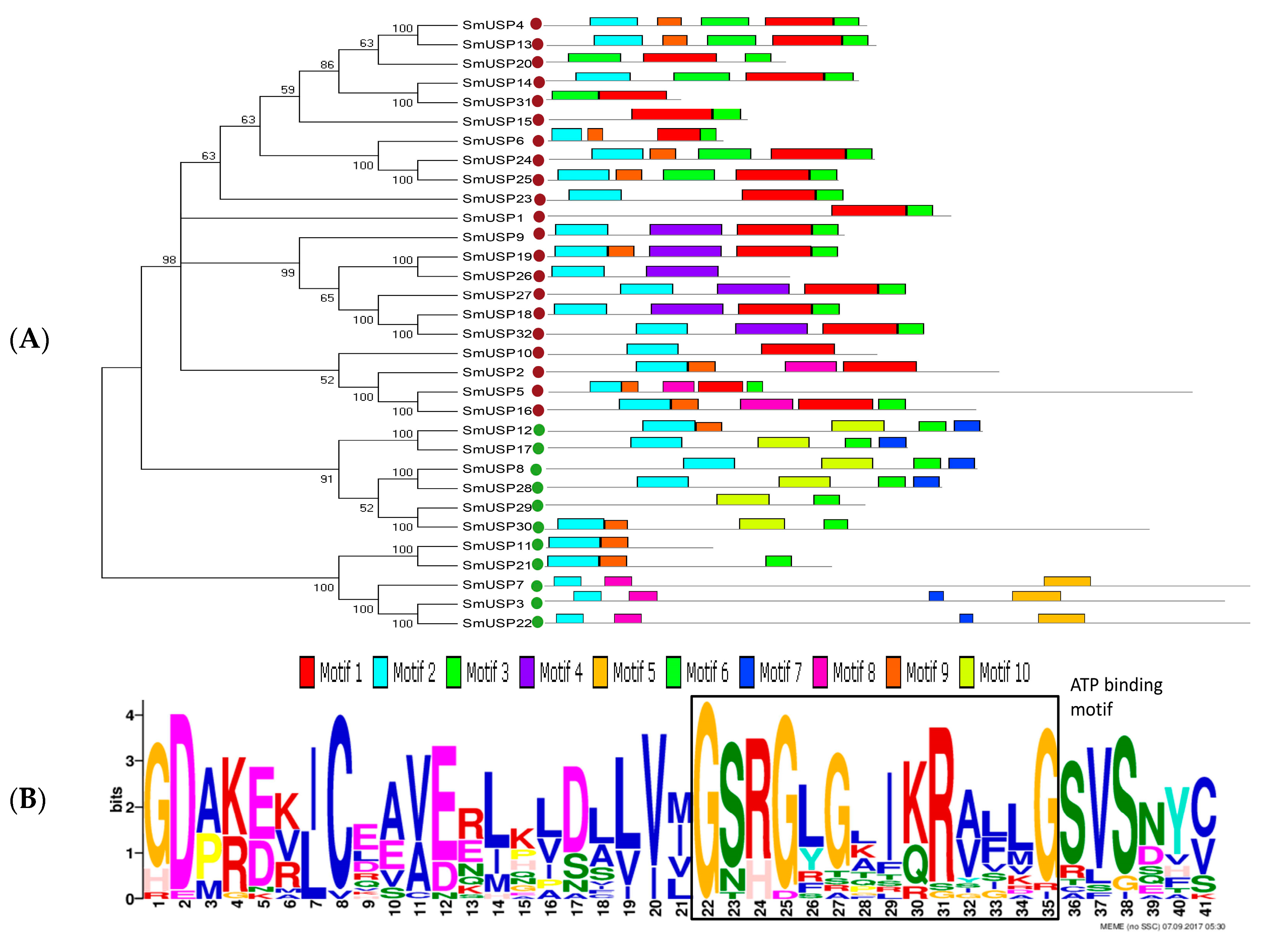
3.1. Gene Identification, Features of Sequences, and Phylogenetic Analysis
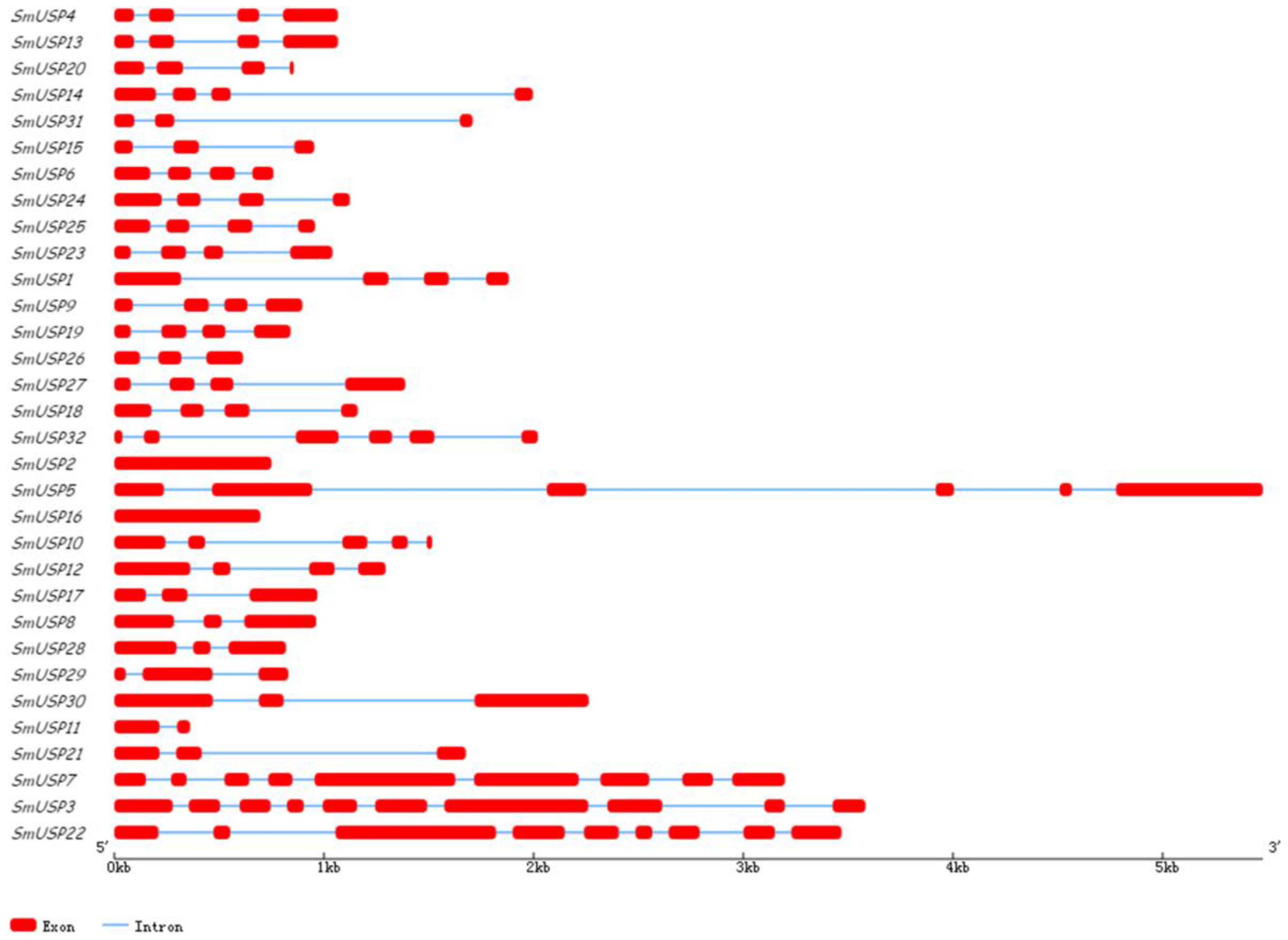
3.2. Conserved Motifs, Multiple Sequence Alignments, Genetic Structures, and Analysis of Subcellular Locations
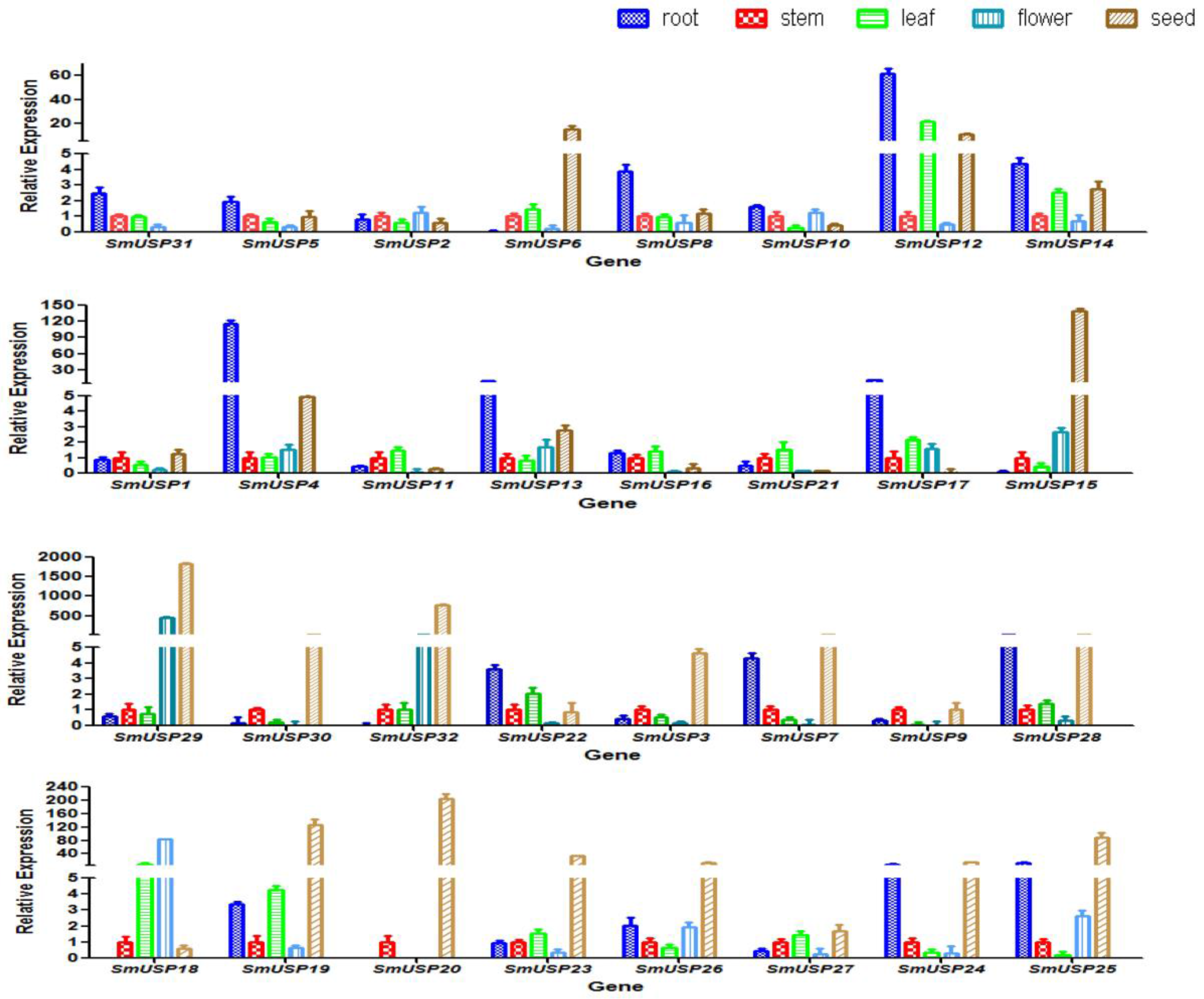
3.3. Patterns of Expression for SmUSP Genes in Various Tissues/Organs
3.4. Analysis of Promoter Sequences and Differential Expression of SmUSP Genes in Response to Salt and Heat Stresses and Their Combination
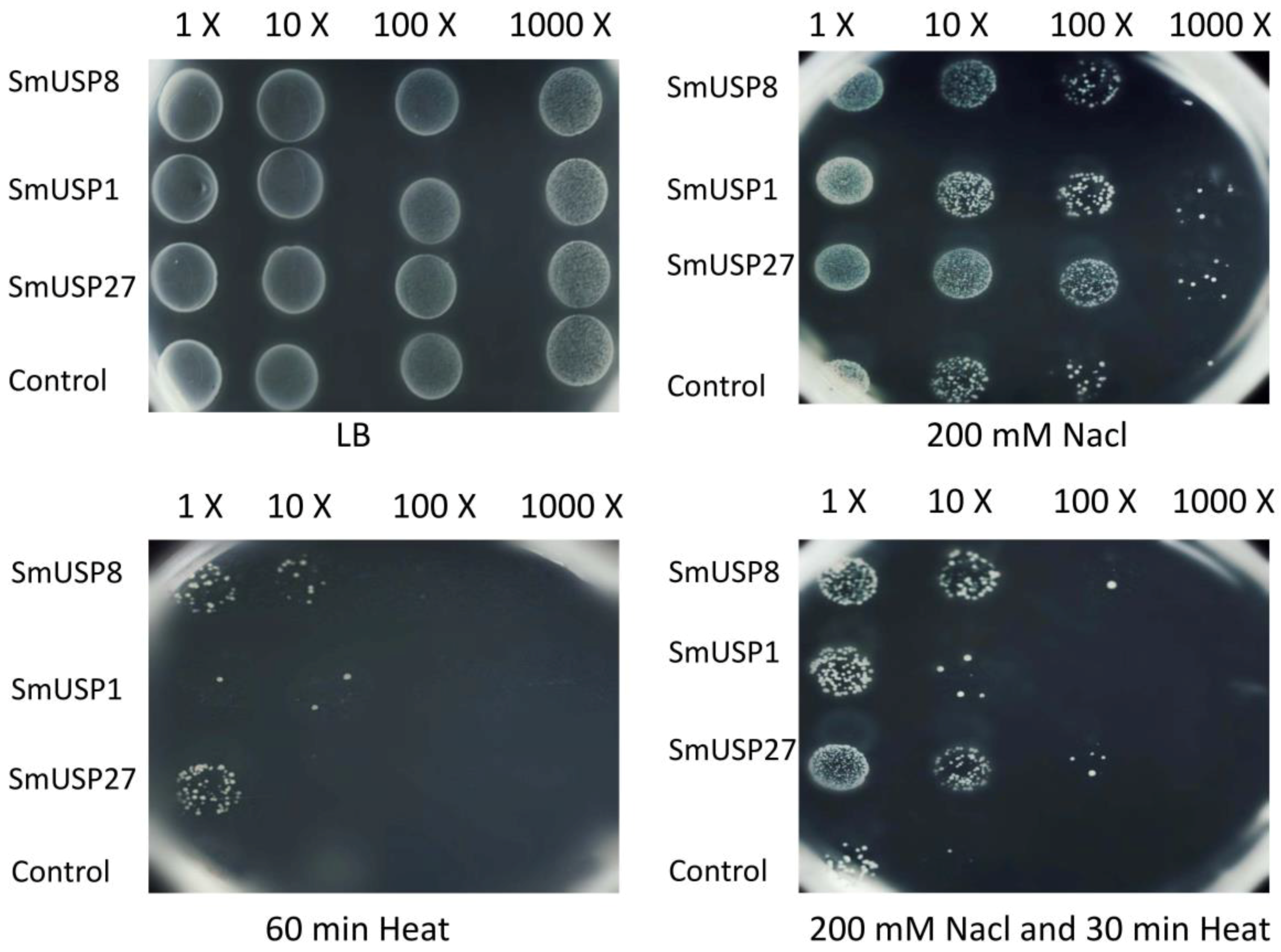
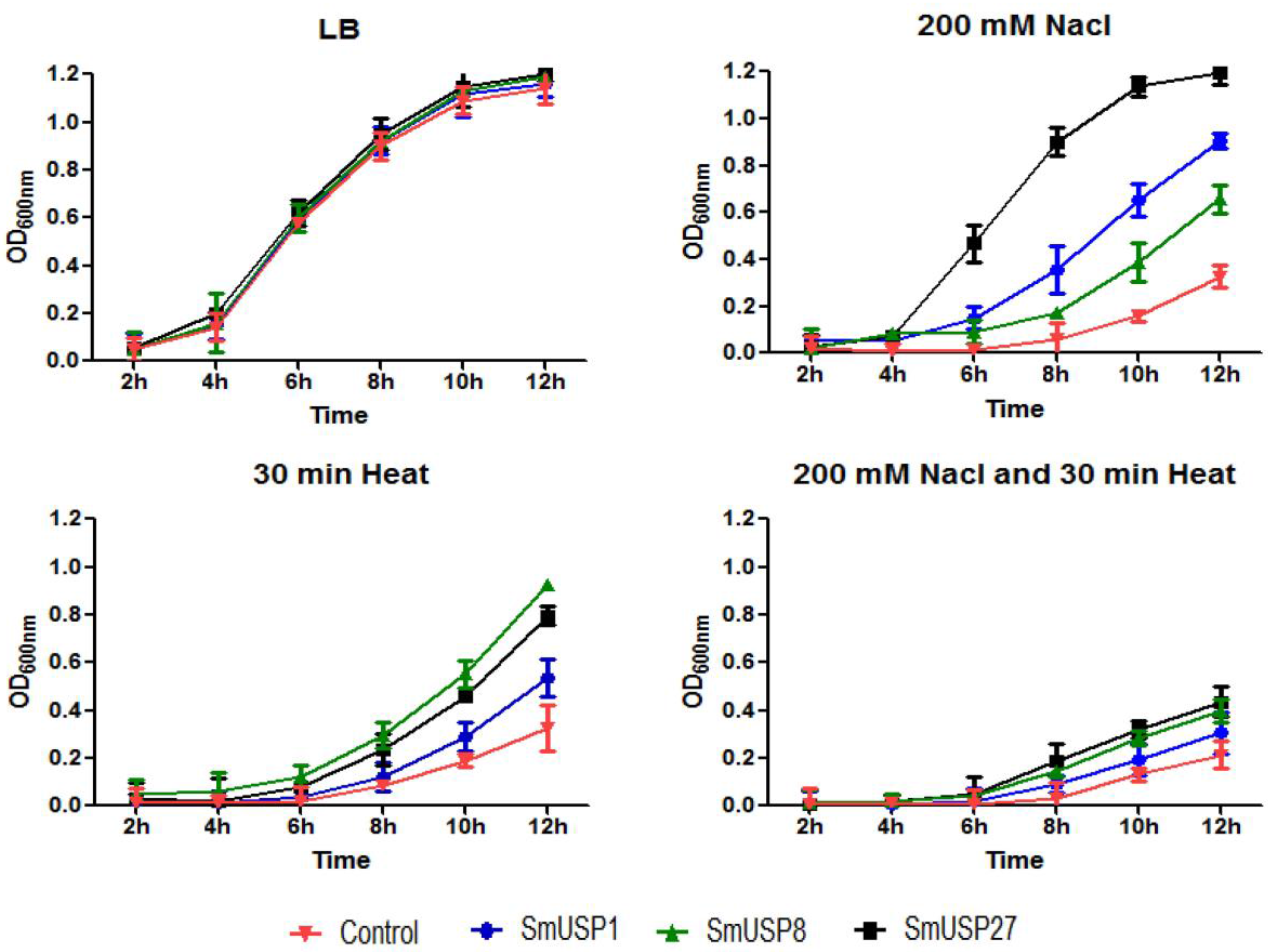
3.5. Heterologous Expression and Functional Validation in E. coli
4. Discussion
4.1. Members of the SmUSP Gene Family and Their Evolution
4.2. Variations in Gene Expression Patterns
4.3. SmUSP Genes Enhance Stress Tolerance in E. coli
4.4. Function of SmUSPs under Simultaneous Stresses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Serrano, R.; Rodriguez-Navarro, A. Ion homeostasis during salt stress in plants. Curr. Opin. Cell Biol. 2001, 13, 399–404. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Heremans, K.; Smeller, L. Protein structure and dynamics at high pressure. Biochim. Biophys. Acta 1998, 1386, 353–370. [Google Scholar] [CrossRef]
- Howarth, C.J. Genetic Improvements of Tolerance to High Temperature; Howarth Press: Philadelphia, PA, USA, 2005. [Google Scholar]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Kerk, D.; Bulgrien, J.; Smith, D.W.; Gribskov, M. Arabidopsis proteins containing similarity to the universal stress protein domain of bacteria. Plant Physiol. 2003, 131, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Ndimba, B.K.; Chivasa, S.; Simon, W.J.; Slabas, A.R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 4185–4196. [Google Scholar] [CrossRef] [PubMed]
- Persson, O.; Valadi, A.; Nyström, T.; Farewell, A. Metabolic control of the Escherichia coli universal stress protein response through fructose-6-phosphate. Mol. Microbiol. 2007, 65, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Kvint, K.; Nachin, L.; Diez, A.; Nyström, T. The bacterial universal stress protein: Function and regulation. Curr. Opin. Microbiol. 2003, 6, 140–145. [Google Scholar] [CrossRef]
- Rachid, L.; Wang, T.; Bo, O.; Khurram, Z.; Li, H.; Zhang, J. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar]
- Li, W.T.; Wei, Y.M.; Wang, J.R.; Liu, C.J.; Lan, X.J.; Jiang, Q.T. Identification, localization, and characterization of putative USP genes in barley. Theor. Appl. Genet. 2010, 121, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Tkaczuk, K.L.; Shumilin, I.A.; Chruszcz, M.; Evdokimova, E.; Savchenko, A.; Minor, W. Structural and functional insight into the universal stress protein family. Evolut. Appl. 2013, 6, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Sauter, M.; Rzewuski, G.; Marwedel, T.; Lorbiecke, R.A. The novel ethylene-regulated gene OsUsp1 from rice encodes a member of a plant protein family related to prokaryotic universal stress proteins. J. Exp. Bot. 2002, 53, 2325–2331. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, A.; Zahur, M.; Husnain, T.; Riazuddin, S. GUSP1 and GUSP2, two drought-responsive genes in Gossypium arboreum have homology to Universal Stress Proteins. Plant Mol. Biol. Rep. 2009, 27, 109–114. [Google Scholar] [CrossRef]
- Chou, M.X.; Wei, X.Y.; Chen, D.S.; Zhou, J.C. A novel nodule enhanced gene encoding a putative universal stress protein from Astragalus sinicus. J. Plant Physiol. 2007, 164, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.D.; Moreira, L.M.; Kapp, D. The nodulin vfENOD18 is an ATP-binding protein in infected cells of Vicia faba L. nodules. Plant Mol. Biol. 2001, 47, 749–759. [Google Scholar] [CrossRef]
- Isokpehi, R.D.; Simmons, S.S.; Cohly, H.H.P.; Ekunwe, S.I.N.; Begonia, G.B.; Ayensu, W.K. identification of drought-responsive universal stress proteins in Viridiplantae. Bioinform. Biol. Insights 2012, 5, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Zahur, M.; Maqbool, A.; Irfan, M.; Barozai, M.Y.K.; Rashid, B.; Riazuddin, S. Isolation and functional analysis of cotton universal stress protein promoter in response to phytohormones and abiotic stresses. Mol. Biol. 2009, 43, 578–585. [Google Scholar] [CrossRef]
- Jung, Y.J.; Melencion, S.M.B.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T. universal stress protein exhibits a redox-dependent chaperone function in arabidopsis and enhances plant tolerance to heat shock and oxidative stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Wu, J.Y. Tanshinone biosynthesis in Salvia miltiorrhiza and production in plant tissue cultures. Appl. Microbiol. Biotechnol. 2010, 88, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Lin, Z.; Zhou, C.C.; Xiao, J.B.; Pan, L.; Kai, G.Y. Metabolic regulation and genetic engineering of pharmaceutical component tanshinone biosynthesis in Salvia miltiorrhiza. J. Med. Plant Res. 2011, 4, 2591–2597. [Google Scholar] [CrossRef]
- Dong, Y.; Morris-Natschke, S.L.; Lee, K.H. Biosynthesis, total syntheses, and antitumor activity of tanshinones and their analogs as potential therapeutic agents. Nat. Prod. Rep. 2011, 28, 529. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lu, S. Genome-wide characterization and comparative analysis of R2R3-MYB transcription factors shows the complexity of MYB-associated regulatory networks in Salvia miltiorrhiza. BMC Genom. 2014, 15, 277. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.B.; Song, J.Y.; Luo, H.M.; Chen, S.L. Analysis of the genome sequence of the medicinal plant Salvia miltiorrhiza. Mol. Plant 2016, 9, 949. [Google Scholar] [CrossRef] [PubMed]
- Udawat, P.; Mishra, A.; Jha, B. Heterologous expression of an uncharacterized universal stress protein gene (SbUSP) from the extreme halophyte, Salicornia brachiata, which confers salt and osmotic tolerance to E. coli. Gene 2014, 536, 163. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.C.; Ji, A.J.; Song, J.Y.; Chen, S.L. Genome-wide analysis of auxin response factor gene family members in medicinal model plant Salvia miltiorrhiza. Biol. Open. 2016, 5, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, R.; Williams, H.D. Universal stress proteins and Mycobacterium tuberculosis. Res. Microbiol. 2003, 154, 387–392. [Google Scholar] [CrossRef]
- Gilbert, W. The exon theory of genes. Cold Spring Harb. Symp. Quant. Biol. 1987, 52, 901. [Google Scholar] [CrossRef] [PubMed]
- Patthy, L. Intron-dependent evolution: Preferred types of exons and introns. FEBS Lett. 1987, 214, 1–7. [Google Scholar] [CrossRef]
- Gutiérrez-Beltrán, E.; Personat, J.M.; de la Torre, F.; del Pozo, O. A universal stress protein involved in oxidative stress is a phosphorylation target for protein kinase CIPK6. Plant Physiol. 2017, 173, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Pvv, P.; Pisipati, S.R.; Momčilović, I.; Ristic, Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop Sci. 2012, 197, 430–441. [Google Scholar]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2014, 37, 1059. [Google Scholar] [CrossRef] [PubMed]









© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-F.; Su, J.; Yang, N.; Zhang, H.; Cao, X.-Y.; Kang, J.-F. Functional Characterization of Selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes 2017, 8, 224. https://doi.org/10.3390/genes8090224
Wang X-F, Su J, Yang N, Zhang H, Cao X-Y, Kang J-F. Functional Characterization of Selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes. 2017; 8(9):224. https://doi.org/10.3390/genes8090224
Chicago/Turabian StyleWang, Xiao-Fan, Jiao Su, Na Yang, Hui Zhang, Xiao-Yan Cao, and Jie-Fang Kang. 2017. "Functional Characterization of Selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli" Genes 8, no. 9: 224. https://doi.org/10.3390/genes8090224
APA StyleWang, X. -F., Su, J., Yang, N., Zhang, H., Cao, X. -Y., & Kang, J. -F. (2017). Functional Characterization of Selected Universal Stress Protein from Salvia miltiorrhiza (SmUSP) in Escherichia coli. Genes, 8(9), 224. https://doi.org/10.3390/genes8090224