Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sheep Blood and Wool Samples
2.2. Search for an Ovine Homologue of Human KRTAP26-1 in the Sheep Genome
2.3. PCR Primers and Amplification of Sheep Genomic DNA
2.4. Screening for Variation in KRTAP26-1
2.5. Sequencing of Allelic Variants and Sequence Analysis
2.6. Statistical Analyses
3. Results
3.1. Correlations between Wool Traits
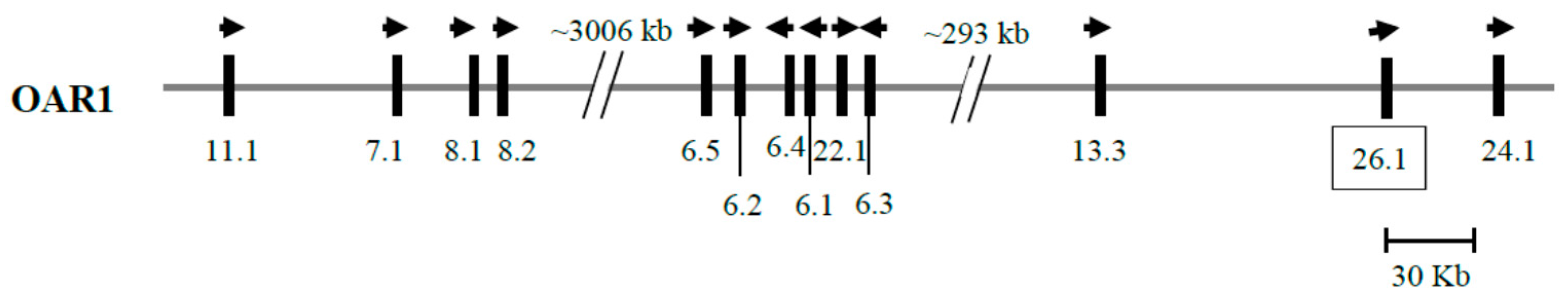
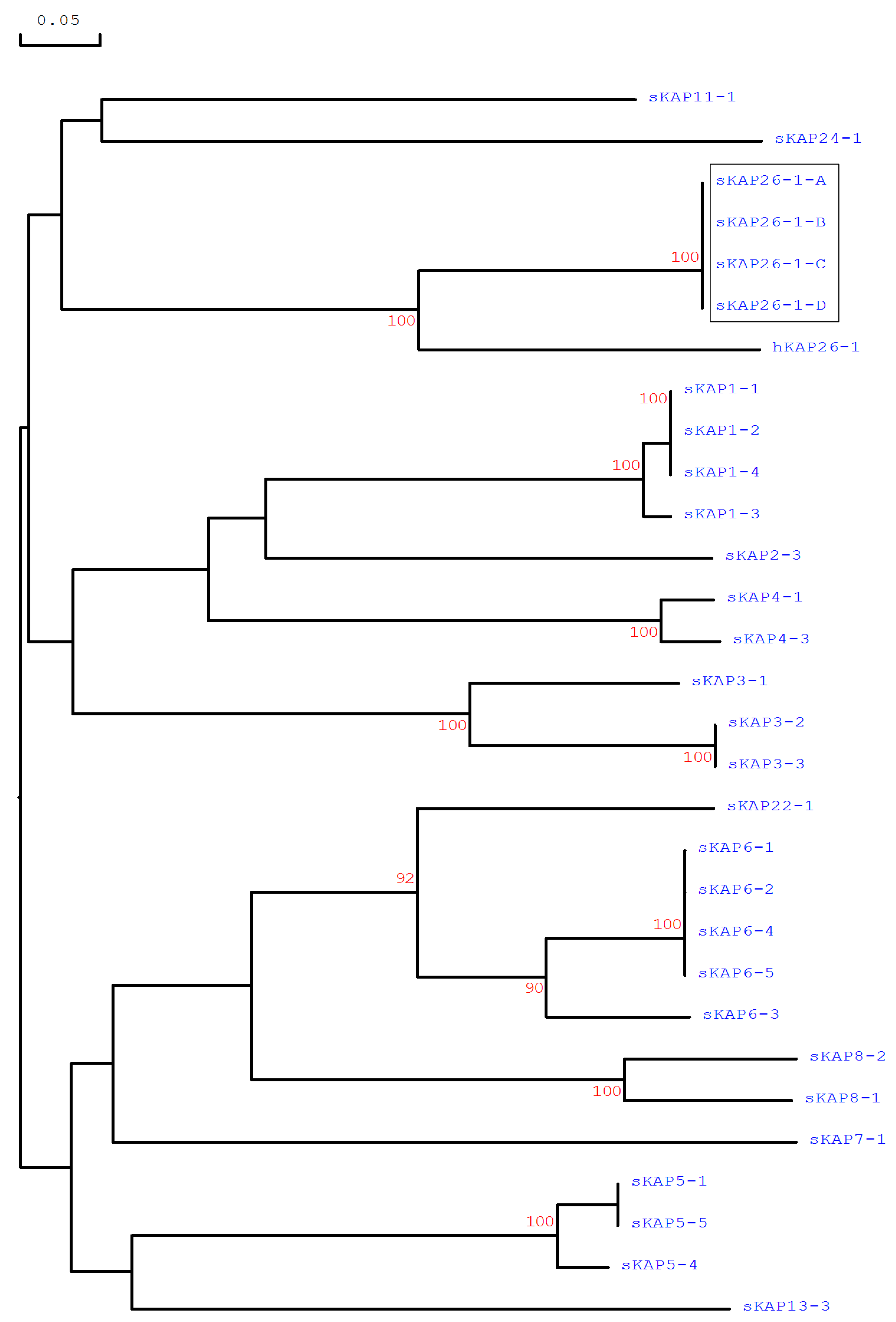
3.2. Identification of KRTAP26-1 in the Sheep Genome
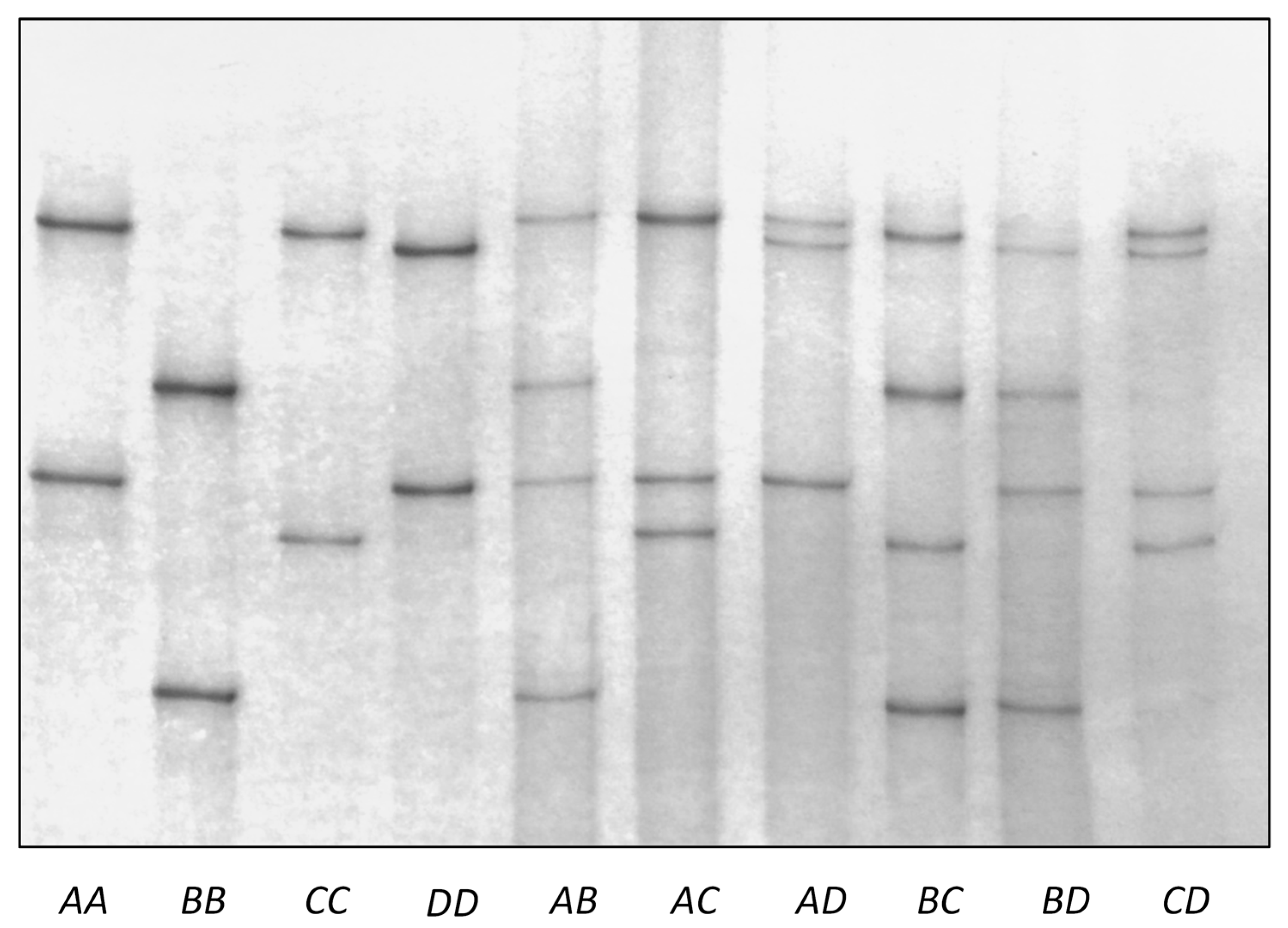
3.3. Variation in Ovine KRTAP26-1
3.4. Comparison of Variant and Genotype Frequencies between NZ Romney and Merino × Southdown-Cross Sheep
3.5. Effect of Variation in KRTAP26-1 on Wool Traits
3.6. Effect of Common Genotypes on Wool Traits
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Powell, B.C.; Rogers, G.E. The role of keratin proteins and their genes in the growth, structure and properties of hair. In Formation and Structure of Human Hair; Jollès, P., Zahn, H., Höcker, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1997; pp. 59–148. [Google Scholar]
- Gong, H.; Zhou, H.; McKenzie, G.W.; Yu, Z.; Clerens, S.; Dyer, J.M.; Plowman, J.E.; Wright, M.W.; Arora, R.; Bawden, C.S.; et al. An updated nomenclature for keratin-associated proteins (KAPs). Int. J. Biol. Sci. 2012, 8, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Zhou, H.; Forrest, R.H.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Wool keratin-associated protein genes in sheep—a review. Genes 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Schweizer, J. Human KAP genes, only the half of it? Extensive size polymorphisms in hair keratin-associated protein genes. J. Investig. Dermatol. 2005, 124, vii–ix. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschläger, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human KAP24.1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Investig. Dermatol. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel Wunder, S.; Giehl, K. Characterization and expression analysis of the hair keratin associated protein KAP26.1. Brit. J. Dermatol. 2008, 159, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hickford, J.G.H.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G.H. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Liu, X.; Luo, Y.; Hickford, J.G. Identification of the ovine keratin-associated protein 22-1 (KAP22-1) gene and its effect on wool traits. Genes 2017, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hodge, S.; Dyer, J.M.; Hickford, J.G. Association of wool traits with variation in the ovine KAP1-2 gene in Merino cross lambs. Small Rum. Res. 2015, 124, 24–29. [Google Scholar] [CrossRef]
- Safari, E.; Fogarty, N.M.; Gilmour, A.R.; Atkins, K.D.; Mortimer, S.I.; Swan, A.A.; Brien, F.D.; Greeff, J.C.; van der Werf, J.H. Genetic correlations among and between wool, growth and reproduction traits in Merino sheep. J. Anim. Breed. Genet. 2007, 124, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Huisman, A.; Brown, D. Genetic parameters for bodyweight, wool, and disease resistance and reproduction traits in Merino sheep. 4. Genetic relationships between and within wool traits. Anim. Prod. Sci. 2009, 49, 289–296. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Yan, W.; Luo, Y.Z.; Hickford, J.G.H. Identification and sequence analysis of the keratin-associated protein 24-1 (KAP24-1) gene homologue in sheep. Gene 2012, 511, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. Identification of the ovine KAP11-1 gene (KRTAP11-1) and genetic variation in its coding sequence. Mol. Biol. Rep. 2011, 38, 5429–5433. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Dyer, J.M.; Plowman, J.E.; Hickford, J.G. Identification of the keratin-associated protein 13-3 (KAP13-3) gene in sheep. Open J. Gen. 2011, 1, 60–64. [Google Scholar] [CrossRef]
- Nakamura, A.; Arimoto, M.; Takeuchi, K.; Fujii, T. A rapid extraction procedure of human hair proteins and identification of phosphorylated species. Biol. Pharm. Bull. 2002, 25, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Hickford, J.G.; Fang, Q.; Lin, Y.S. Allelic variation of the ovine Toll-like receptor 4 gene. Dev. Comp. Immunol. 2007, 1, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G. Polymorphism of the ovine keratin-associated protein 1-4 gene (KRTAP1-4). Mol. Biol. Rep. 2010, 37, 3377–3380. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Yu, Z.; Dyer, J.; Plowman, J.E.; Hickford, J. Identification of the ovine keratin-associated protein KAP1-2 gene (KRTAP1-2). Exp. Dermatol. 2011, 20, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, A.J.; OlivosBGlander, I.; Kusukawa, N.; Highsmith, W.E., Jr. Single-strand conformation polymorphism and heteroduplex analysis for gel-based mutation detection. Electrophoresis 1999, 20, 1177–1185. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Malau-Aduli, A.E.O. A review of sheep wool quality traits. Annu. Rev. Res. Biol. 2012, 2, 1–14. [Google Scholar]
- Rippon, J.A. The structure of wool. In The Coloration of Wool and Other Keratin Fibres; Lewis, D.M., Rippon, J.A., Eds.; John Wiley & Sons in association with the Society of Dyers and Colourists; Plenum Press: Bradford, West Yorkshire, UK, 2013; pp. 1–42. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| GFW | CFW | Yield | MSL | MFD | FDSD | CVFD | MSS | MFC | |
|---|---|---|---|---|---|---|---|---|---|
| CFW | 0.916 *** | ||||||||
| Yield | 0.165 ** | 0.528 *** | |||||||
| MSL | 0.463 *** | 0.525 *** | 0.341 *** | ||||||
| MFD | 0.053 | −0.091 | −0.341 *** | −0.270 *** | |||||
| FDSD | 0.160 ** | 0.263 *** | 0.309 *** | 0.350 *** | 0.796 *** | ||||
| CVFD | 0.304 *** | 0.334 *** | 0.174 ** | 0.306 *** | 0.306 *** | 0.815 *** | |||
| MSS | 0.188 *** | 0.220 *** | 0.143 *** | 0.067 | 0.069 | −0.143 ** | 0.290 *** | ||
| MFC | 0.329 *** | 0.492 *** | 0.550 *** | 0.442 *** | 0.425 *** | 0.397 *** | 0.238 *** | 0.079 | |
| PF | 0.022 | 0.085 | 0.251 *** | 0.244 *** | 0.819 *** | 0.794 *** | 0.440 *** | 0.061 | 0.224 *** |
| Nucleotide Position | Variant | Amino Acid Change | |||
|---|---|---|---|---|---|
| KRTAP26-1*A | KRTAP26-1*B | KRTAP26-1*C | KRTAP26-1*D | ||
| c.72 | C | T | C | C | No change |
| c.111 | A | C | T | A | No change |
| c.123 | C | G | C | C | No change |
| c.215 | G | A | G | G | p.Cys72Tyr |
| c.277 | A | A | G | A | p.Ser93Gly |
| c.336 | G | G | G | A | No change |
| c.549 | C | T | C | C | No change |
| Trait | Variant Assessed | n | Single-Variant Model | Multi-Variant Model | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Absent | Present | Absent | Present | p | Variants Fitted | Absent | Present | p | ||
| GFW | A | 71 | 312 | 2.37 ± 0.06 | 2.37 ± 0.03 | 0.958 | ||||
| (kg) | B | 207 | 176 | 2.37 ± 0.04 | 2.37 ± 0.04 | 0.902 | ||||
| C | 212 | 171 | 2.39 ± 0.03 | 2.33 ± 0.05 | 0.361 | |||||
| CFW | A | 71 | 312 | 1.72 ± 0.05 | 1.71 ± 0.02 | 0.764 | ||||
| (kg) | B | 207 | 176 | 1.72 ± 0.03 | 1.70 ± 0.03 | 0.678 | ||||
| C | 212 | 171 | 1.71 ± 0.03 | 1.72 ± 0.04 | 0.877 | |||||
| Yield | A | 71 | 312 | 72.2 ± 0.74 | 71.7 ± 0.35 | 0.471 | B, C | 73.0 ± 0.82 | 71.8 ± 0.41 | 0.201 |
| (%) | B | 207 | 176 | 72.4 ± 0.47 | 71.2 ± 0.44 | 0.070 | C | 72.4 ± 0.47 | 71.7 ± 0.52 | 0.291 |
| C | 212 | 171 | 71.2 ± 0.40 | 73.0 ± 0.65 | 0.027 | B | 71.3 ± 0.41 | 72.8 ± 0.68 | 0.082 | |
| MSL (mm) | A | 71 | 312 | 84.7 ± 1.66 | 82.6 ± 0.87 | 0.253 | C | 85.6 ± 1.64 | 84.0 ± 0.93 | 0.377 |
| B | 207 | 176 | 83.9 ± 1.12 | 82.2 ± 1.04 | 0.236 | C | 84.3 ± 1.10 | 84.4 ± 1.18 | 0.950 | |
| C | 212 | 171 | 80.7 ± 0.99 | 87.9 ± 1.51 | <0.001 | None | 80.7 ± 0.99 | 87.9 ± 1.51 | <0.001 | |
| MFD | A | 71 | 312 | 19.8 ± 0.25 | 19.6 ± 0.13 | 0.375 | B, C | 19.6 ± 0.27 | 19.3 ± 0.14 | 0.397 |
| (µm) | B | 207 | 176 | 19.3 ± 0.17 | 19.9 ± 0.15 | 0.001 | C | 19.2 ± 0.16 | 19.6 ± 0.17 | 0.081 |
| C | 212 | 171 | 20.1 ± 0.15 | 18.6 ± 0.22 | <0.001 | B | 20.0 ± 0.15 | 18.7 ± 0.23 | <0.001 | |
| FDSD | A | 71 | 312 | 4.26 ± 0.09 | 4.22 ± 0.04 | 0.680 | B, C | 4.21 ± 0.09 | 4.13 ± 0.04 | 0.450 |
| (µm) | B | 207 | 176 | 4.16 ± 0.06 | 4.28 ± 0.05 | 0.098 | C | 4.13 ± 0.06 | 4.17 ± 0.06 | 0.626 |
| C | 212 | 171 | 4.35 ± 0.05 | 3.96 ± 0.08 | <0.001 | B | 4.34 ± 0.05 | 3.96 ± 0.08 | <0.001 | |
| CVFD | A | 71 | 312 | 21.4 ± 0.28 | 21.9 ± 0.14 | 0.166 | C | 21.4 ± 0.28 | 21.7 ± 0.15 | 0.248 |
| (%) | B | 207 | 176 | 21.9 ± 0.18 | 21.7 ± 0.17 | 0.471 | A, C | 21.7 ± 0.26 | 21.4 ± 0.20 | 0.330 |
| C | 212 | 171 | 22.0 ± 0.16 | 21.4 ± 0.25 | 0.067 | A | 21.8 ± 0.19 | 21.3 ± 0.26 | 0.093 | |
| MSS | A | 71 | 312 | 23.9 ± 1.06 | 22.6 ± 0.55 | 0.237 | B | 23.6 ± 1.19 | 22.7 ± 0.55 | 0.496 |
| (N/ktex) | B | 207 | 176 | 22.1 ± 0.71 | 23.4 ± 0.65 | 0.145 | None | 22.1 ± 0.71 | 23.4 ± 0.65 | 0.145 |
| C | 212 | 171 | 23.1 ± 0.64 | 22.2 ± 0.98 | 0.434 | B | 22.9 ± 0.66 | 22.6 ± 1.01 | 0.774 | |
| MFC | A | 71 | 312 | 91.2 ± 1.80 | 89.5 ± 0.85 | 0.367 | B, C | 89.4 ± 2.00 | 89.1 ± 1.01 | 0.862 |
| (°/mm) | B | 207 | 176 | 87.7 ± 1.15 | 91.4 ± 1.06 | 0.016 | C | 87.6 ± 1.16 | 90.7 ± 1.26 | 0.056 |
| C | 212 | 171 | 90.8 ± 0.98 | 87.2 ± 1.60 | 0.070 | B | 90.3 ± 1.02 | 88.0 ± 1.65 | 0.278 | |
| PF | A | 71 | 312 | 3.00 ± 0.45 | 2.79 ± 0.23 | 0.654 | B, C | 2.55 ± 0.49 | 2.44 ± 0.26 | 0.845 |
| (%) | B | 207 | 176 | 2.33 ± 0.30 | 3.24 ± 0.27 | 0.017 | C | 2.23 ± 0.29 | 2.69 ± 0.31 | 0.237 |
| C | 212 | 171 | 3.47 ± 0.26 | 1.41 ± 0.40 | <0.001 | B | 3.39 ± 0.27 | 1.53 ± 0.41 | <0.001 | |
| Trait | AA (n = 68) | AB (n = 116) | AC (n = 123) | BB (n = 22) | BC (n = 37) | p |
|---|---|---|---|---|---|---|
| GFW (kg) | 2.39 ± 0.05 | 2.36 ± 0.04 | 2.36 ± 0.05 | 2.39 ± 0.09 | 2.40 ± 0.08 | 0.971 |
| CFW (kg) | 1.72 ± 0.04 | 1.68 ± 0.03 | 1.72 ± 0.04 | 1.69 ± 0.07 | 1.77 ± 0.06 | 0.814 |
| Yield (%) | 73.5 ± 1.32 | 72.7 ± 1.22 | 74.6 ± 1.28 | 72.7 ± 1.65 | 74.5 ± 1.58 | 0.310 |
| MSL (mm) | 82.6 ± 1.41 a,b | 81.2 ± 1.07 b | 86.5 ± 1.45 a | 78.2 ± 2.48 b | 88.2 ± 2.17 a | 0.002 |
| MFD (µm) | 19.7 ± 0.22 a,b | 20.2 ± 0.17 a | 18.7 ± 0.23 c | 20.8 ± 0.39 a | 19.1 ± 0.34 b,c | <0.001 |
| FDSD (µm) | 4.41 ± 0.08 a | 4.46 ± 0.06 a | 4.14 ± 0.08 b | 4.67 ± 0.13 a | 4.15 ± 0.12 b | 0.001 |
| CVFD (%) | 22.3 ± 0.26 | 22.0 ± 0.20 | 22.0 ± 0.27 | 22.4 ± 0.47 | 21.7 ± 0.41 | 0.624 |
| MSS (N/ktex) | 22.1 ± 0.89 | 22.7 ± 0.68 | 21.4 ± 0.92 | 25.6 ± 1.58 | 22.7 ± 1.38 | 0.202 |
| MFC (o/mm) | 89.1 ± 1.76 | 91.8 ± 1.31 | 87.0 ± 1.69 | 92.1 ± 3.01 | 88.8 ± 2.61 | 0.260 |
| PF (%) | 3.21 ± 0.40 a,b | 3.50 ± 0.32 a | 1.36 ± 0.15 b | 5.04 ± 1.08 a | 1.39 ± 0.24 b | <0.001 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Hu, J.; Luo, Y.; Hickford, J.G.H. Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits. Genes 2017, 8, 225. https://doi.org/10.3390/genes8090225
Li S, Zhou H, Gong H, Zhao F, Hu J, Luo Y, Hickford JGH. Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits. Genes. 2017; 8(9):225. https://doi.org/10.3390/genes8090225
Chicago/Turabian StyleLi, Shaobin, Huitong Zhou, Hua Gong, Fangfang Zhao, Jiang Hu, Yuzhu Luo, and Jon G. H. Hickford. 2017. "Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits" Genes 8, no. 9: 225. https://doi.org/10.3390/genes8090225
APA StyleLi, S., Zhou, H., Gong, H., Zhao, F., Hu, J., Luo, Y., & Hickford, J. G. H. (2017). Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits. Genes, 8(9), 225. https://doi.org/10.3390/genes8090225