The IFNG rs1861494 Single Nucleotide Polymorphism Is Associated with Protection against Tuberculosis Disease in Argentina

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Demographic Characteristics of the Populations Studied
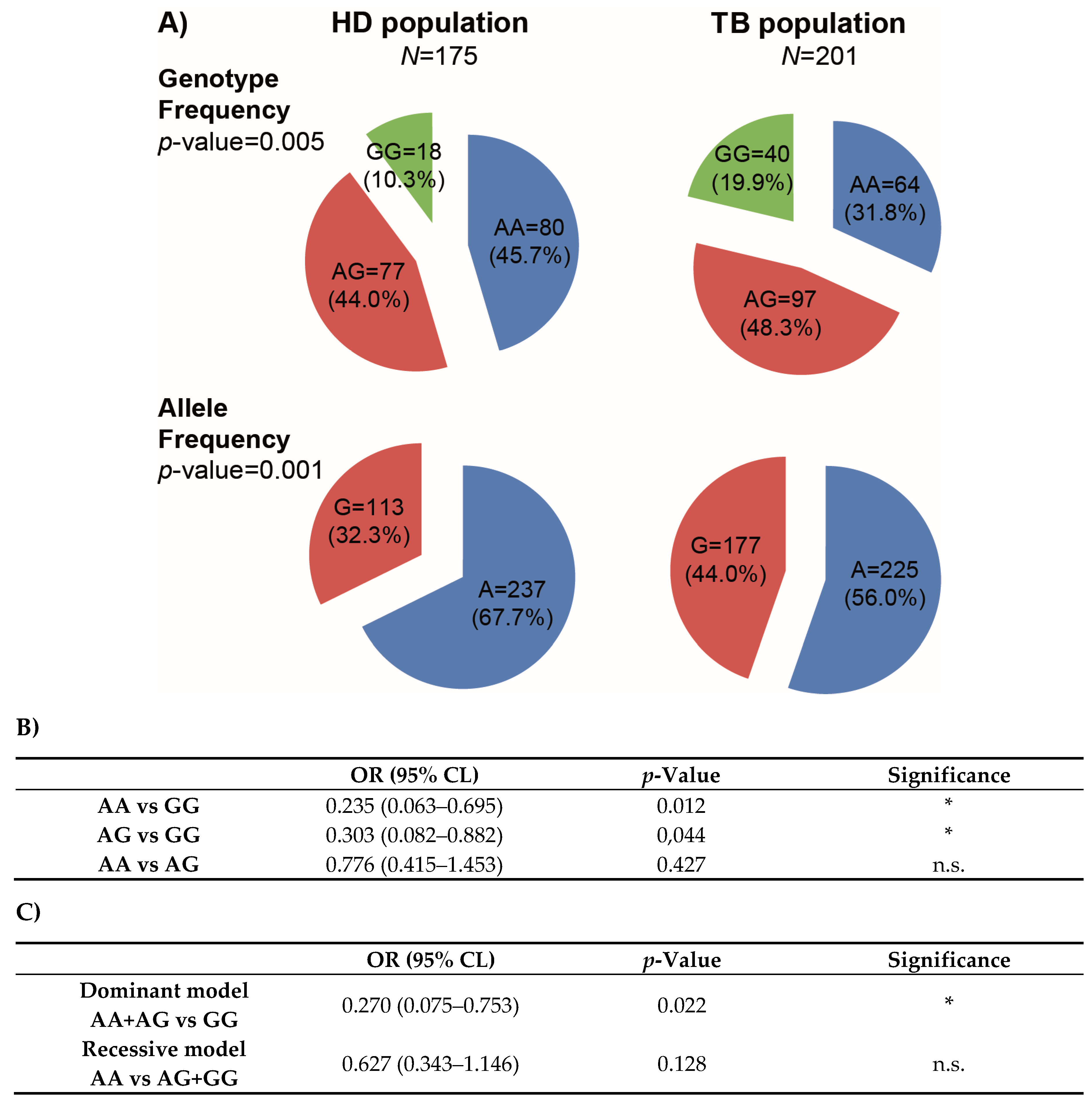
3.2. The IFNG rs1861494 SNP as a Biomarker for Tuberculosis Protection in Argentina
3.3. Lack of Association between the IFNG rs1861494 SNP and the Severity of the Disease
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report. 2016. Available online: http://apps.who.int/iris/bitstream/10665/250441/1/9789241565394-eng.pdf?ua=1 (accessed on 20 October 2017).
- Administración Nacional de Laboratorios e Institutos de Salud (ANLIS). Notificación de Casos de Tuberculosis en la República Argentina. Available online: http://www.anlis.gov.ar/iner/wp-content/uploads/2016/11/Notificacion-de-casos-de-Tuberculosis-en-la-Republica-Argentina-Periodo-1980-2015.pdf (accessed on 20 October 2017).
- Fortin, A.; Abel, L.; Casanova, J.L.; Gros, P. Host genetics of mycobacterial diseases in mice and men: Forward genetic studies of BCG-osis and tuberculosis. Annu. Rev. Genom. Hum. Genet. 2007, 8, 163–192. [Google Scholar] [CrossRef] [PubMed]
- Philips, J.A.; Ernst, J.D. Tuberculosis pathogenesis and immunity. Annu. Rev. Pathol. 2012, 7, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Jasenosky, L.D.; Scriba, T.J.; Hanekom, W.A.; Goldfeld, A.E. T cells and adaptive immunity to Mycobacterium tuberculosis in humans. Immunol. Rev. 2015, 264, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Triebold, K.J.; Dalton, D.K.; Stewart, T.A.; Bloom, B.R. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J. Exp. Med. 1993, 178, 2249–2254. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Dalton, D.K.; Stewart, T.A.; Griffin, J.P.; Russell, D.G.; Orme, I.M. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J. Exp. Med. 1993, 178, 2243–2247. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.; Xing, Y.; Magliozzo, R.S.; Bloom, B.R. Killing of virulent Mycobacterium tuberculosis by reactive nitrogen intermediates produced by activated murine macrophages. J. Exp. Med. 1992, 175, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D.; Taylor, G.A.; McKinney, J.D. Immune control of tuberculosis by IFN-γ-inducible LRG-47. Science 2003, 302, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Rovetta, A.I.; Pena, D.; Hernandez Del Pino, R.E.; Recalde, G.M.; Pellegrini, J.; Bigi, F.; Musella, R.M.; Palmero, D.J.; Gutierrez, M.; Colombo, M.I.; et al. IFNG-mediated immune responses enhance autophagy against Mycobacterium tuberculosis antigens in patients with active tuberculosis. Autophagy 2014, 10, 2109–2121. [Google Scholar] [CrossRef] [PubMed]
- Fabri, M.; Stenger, S.; Shin, D.M.; Yuk, J.M.; Liu, P.T.; Realegeno, S.; Lee, H.M.; Krutzik, S.R.; Schenk, M.; Sieling, P.A.; et al. Vitamin D is required for IFN-γ-mediated antimicrobial activity of human macrophages. Sci. Transl. Med. 2011, 3, 104ra102. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, A.; Gong, J.; Silva, C.; Qian, D.; Barnes, P.F. Clinical correlates of interferon gamma production in patients with tuberculosis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1997, 25, 617–620. [Google Scholar] [CrossRef]
- Pasquinelli, V.; Quiroga, M.F.; Martinez, G.J.; Zorrilla, L.C.; Musella, R.M.; Bracco, M.M.; Belmonte, L.; Malbran, A.; Fainboim, L.; Sieling, P.A.; et al. Expression of signaling lymphocytic activation molecule-associated protein interrupts IFN-γ production in human tuberculosis. J. Immunol. 2004, 172, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Thillai, M.; Pollock, K.; Pareek, M.; Lalvani, A. Interferon-γ release assays for tuberculosis: Current and future applications. Expert Rev. Respir. Med. 2014, 8, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.G.; Moraes, M.O. Genetic polymorphisms of infectious diseases in case-control studies. Dis. Mark. 2009, 27, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, J.; Boisson-Dupuis, S.; Abel, L.; Casanova, J.L. Mendelian susceptibility to mycobacterial disease: Genetic, immunological, and clinical features of inborn errors of IFN-γ immunity. Semin. Immunol. 2014, 26, 454–470. [Google Scholar] [CrossRef] [PubMed]
- Hyvarinen, K.; Ritari, J.; Koskela, S.; Niittyvuopio, R.; Nihtinen, A.; Volin, L.; Gallardo, D.; Partanen, J. Genetic polymorphism related to monocyte-macrophage function is associated with graft-versus-host disease. Sci. Rep. 2017, 7, 15666. [Google Scholar] [CrossRef] [PubMed]
- Abana, C.O.; Bingham, B.S.; Cho, J.H.; Graves, A.J.; Koyama, T.; Pilarski, R.T.; Chakravarthy, A.B.; Xia, F. IL-6 variant is associated with metastasis in breast cancer patients. PLoS ONE 2017, 12, e0181725. [Google Scholar] [CrossRef] [PubMed]
- Lemos, N.E.; Dieter, C.; Dorfman, L.E.; Assmann, T.S.; Duarte, G.C.K.; Canani, L.H.; Bauer, A.C.; Crispim, D. The rs2292239 polymorphism in ERBB3 gene is associated with risk for type 1 diabetes mellitus in a Brazilian population. Gene 2017. [Google Scholar] [CrossRef] [PubMed]
- Manolova, I.; Miteva, L.; Ivanova, M.; Kundurzhiev, T.; Stoilov, R.; Stanilova, S. The synergistic effect of TNFα and IL10 promoter polymorphisms on genetic predisposition to systemic Lupus erythematosus. Genet. Test. Mol. Biomark. 2018. [Google Scholar] [CrossRef] [PubMed]
- Czepiel, J.; Biesiada, G.; Drozdz, M.; Gdula-Argasinska, J.; Zuranska, J.; Marchewka, J.; Perucki, W.; Wolkow, P.; Garlicki, A. The presence of IL-8 +781 T/C polymorphism is associated with the parameters of severe Clostridium difficile infection. Microb. Pathog. 2017, 114, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Talaat, R.M.; Abdelkhalek, M.S.; El-Maadawy, E.A.; Abdel-Mageed, W.S.; El-Shenawy, S.Z.; Osman, M.A. Association of TNF-Alpha gene polymorphisms and susceptibility to hepatitis B virus infection in Egyptians. Hum. Immunol. 2017, 78, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhou, B.; Wang, W.; Zhang, M.; Zhao, Y.; Wang, Z.; Yang, L.; Zhai, J.; Feng, C.G.; Wang, J.; et al. A functional single-nucleotide polymorphism in the promoter of the gene encoding interleukin 6 is associated with susceptibility to tuberculosis. J. Infect. Dis. 2012, 205, 1697–1704. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Ke, Z.; Liu, S.; Tang, B.; Wang, J.; Huang, H.; Chen, S. IL-1β+3953C/T, -511T/C and IL-6 -174C/G polymorphisms in association with tuberculosis susceptibility: A meta-analysis. Gene 2015, 573, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, G.; Xu, Y.; Song, N.; Shen, S.; Jiang, D.; Zeng, W.; Wang, H. Association between IL12B polymorphisms and tuberculosis risk: A meta-analysis. Infect. Genet. Evol. J. Mol. Epidemiol. Evolut. Genet. Infect. Dis. 2014, 21, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.G.; Cardoso, C.C.; Moraes, M.O. IFNG +874T/A, IL10 -1082G/A and TNF -308G/A polymorphisms in association with tuberculosis susceptibility: A meta-analysis study. Hum. Genet. 2008, 123, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Rolandelli, A.; Hernandez Del Pino, R.E.; Pellegrini, J.M.; Tateosian, N.L.; Amiano, N.O.; de la Barrera, S.; Casco, N.; Gutierrez, M.; Palmero, D.J.; Garcia, V.E. The IL-17A rs2275913 single nucleotide polymorphism is associated with protection to tuberculosis but related to higher disease severity in Argentina. Sci. Rep. 2017, 7, 40666. [Google Scholar] [CrossRef] [PubMed]
- Gonsky, R.; Deem, R.L.; Landers, C.J.; Haritunians, T.; Yang, S.; Targan, S.R. IFNG rs1861494 polymorphism is associated with IBD disease severity and functional changes in both IFNG methylation and protein secretion. Inflamm. Bowel Dis. 2014, 20, 1794–1801. [Google Scholar] [CrossRef] [PubMed]
- Peresi, E.; Oliveira, L.R.; da Silva, W.L.; da Costa, E.A.; Araujo, J.P., Jr.; Ayres, J.A.; Fortes, M.R.; Graviss, E.A.; Pereira, A.C.; Calvi, S.A. Cytokine polymorphisms, their influence and levels in Brazilian patients with pulmonary tuberculosis during antituberculosis treatment. Tuberc. Res. Treat. 2013, 2013, 285094. [Google Scholar] [CrossRef] [PubMed]
- Little, S. Amplification-refractory mutation system (ARMS) analysis of point mutations. Curr. Protoc. Hum. Genet. 2001, 9, 9.8.1–9.8.12. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 20 October 2017).
- Morris, G.A.; Edwards, D.R.; Hill, P.C.; Wejse, C.; Bisseye, C.; Olesen, R.; Edwards, T.L.; Gilbert, J.R.; Myers, J.L.; Stryjewski, M.E.; et al. Interleukin 12b (IL12B) genetic variation and pulmonary tuberculosis: A study of cohorts from the Gambia, Guinea-Bissau, United States and Argentina. PLoS ONE 2011, 6, e16656. [Google Scholar] [CrossRef]
- NIH. dbSNP Short Genetic Variations. Available online: https://www.ncbi.nlm.nih.gov/projects/SNP/ (accessed on 20 October 2017).
- Gao, J.; Wei, L.; Liu, X.; Wang, L.; Niu, D.; Jin, T.; Yao, G.; Wang, M.; Yu, Q.; Fu, R. Association between IFN-γ gene polymorphisms and IgA nephropathy in a Chinese Han population. Kidney Blood Press. Res. 2017, 42, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Karam, R.; Essam, H.; Khater, M. Genetic variants of INF-gamma and its mRNA expression and inflammatory parameters in vitiligo pathogenesis. Biochem. Cell Biol. 2017, 95, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.C.; Pereira, A.C.; Brito-de-Souza, V.N.; Dias-Baptista, I.M.; Maniero, V.C.; Venturini, J.; Vilani-Moreno, F.R.; de Souza, F.C.; Ribeiro-Alves, M.; Sarno, E.N.; et al. IFNG +874 T>A single nucleotide polymorphism is associated with leprosy among Brazilians. Hum. Genet. 2010, 128, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Varahram, M.; Farnia, P.; Nasiri, M.J.; Karahrudi, M.A.; Dizagie, M.K.; Velayati, A.A. Association of Mycobacterium tuberculosis lineages with IFN-γ and TNF-α gene polymorphisms among pulmonary tuberculosis patient. Mediterr. J. Hematol. Infect. Dis. 2014, 6, e2014015. [Google Scholar] [CrossRef] [PubMed]
- Abhimanyu, M.B.; Jha, P. Indian Genome Variation Consortium. Footprints of genetic susceptibility to pulmonary tuberculosis: Cytokine gene variants in north Indians. Indian J. Med. Res. 2012, 135, 763–770. [Google Scholar]
- Lee, S.W.; Chuang, T.Y.; Huang, H.H.; Lee, K.F.; Chen, T.T.; Kao, Y.H.; Wu, L.S. Interferon gamma polymorphisms associated with susceptibility to tuberculosis in a Han Taiwanese population. J. Microbiol. Immunol. Infect. 2015, 48, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Etokebe, G.E.; Bulat-Kardum, L.; Johansen, M.S.; Knezevic, J.; Balen, S.; Matakovic-Mileusnic, N.; Matanic, D.; Flego, V.; Pavelic, J.; Beg-Zec, Z.; et al. Interferon-γ gene (T874A and G2109A) polymorphisms are associated with microscopy-positive tuberculosis. Scand. J. Immunol. 2006, 63, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Cui, W.; Guan, L.; Shen, F.; Xu, J.; Zhou, F.; Li, M.; Gao, C.; Jin, Q.; et al. Potential association of pulmonary tuberculosis with genetic polymorphisms of toll-like receptor 9 and interferon-γ in a chinese population. BMC Infect. Dis. 2013, 13, 511. [Google Scholar] [CrossRef] [PubMed]
- Avena, S.; Via, M.; Ziv, E.; Perez-Stable, E.J.; Gignoux, C.R.; Dejean, C.; Huntsman, S.; Torres-Mejia, G.; Dutil, J.; Matta, J.L.; et al. Heterogeneity in genetic admixture across different regions of Argentina. PLoS ONE 2012, 7, e34695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homburger, J.R.; Moreno-Estrada, A.; Gignoux, C.R.; Nelson, D.; Sanchez, E.; Ortiz-Tello, P.; Pons-Estel, B.A.; Acevedo-Vasquez, E.; Miranda, P.; Langefeld, C.D.; et al. Genomic insights into the ancestry and demographic history of south America. PLoS Genet. 2015, 11, e1005602. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, V.; Rovetta, A.I.; Alvarez, I.B.; Jurado, J.O.; Musella, R.M.; Palmero, D.J.; Malbran, A.; Samten, B.; Barnes, P.F.; Garcia, V.E. Phosphorylation of mitogen-activated protein kinases contributes to interferon gamma production in response to Mycobacterium tuberculosis. J. Infect. Dis. 2013, 207, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Jurado, J.O.; Pasquinelli, V.; Alvarez, I.B.; Pena, D.; Rovetta, A.I.; Tateosian, N.L.; Romeo, H.E.; Musella, R.M.; Palmero, D.; Chuluyan, H.E.; et al. IL-17 and IFN-γ expression in lymphocytes from patients with active tuberculosis correlates with the severity of the disease. J. Leukoc. Biol. 2012, 91, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Barnes, P.F.; Dobos-Elder, K.M.; Townsend, J.C.; Chung, Y.T.; Shams, H.; Weis, S.E.; Samten, B. Esat-6 inhibits production of IFN-γ by Mycobacterium tuberculosis-responsive human T cells. J. Immunol. 2009, 182, 3668–3677. [Google Scholar] [CrossRef] [PubMed]
- Via, L.E.; Tsytsykova, A.V.; Rajsbaum, R.; Falvo, J.V.; Goldfeld, A.E. The transcription factor NFATp plays a key role in susceptibility to TB in mice. PLoS ONE 2012, 7, e41427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Ghosh, B. A single nucleotide polymorphism (A --> G) in intron 3 of IFNγ gene is associated with asthma. Genes Immun. 2008, 9, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Hsu, Y.W.; Lu, H.F.; Wong, H.S.; Yu, H.R.; Kuo, H.C.; Huang, F.C.; Chang, W.C.; Kuo, H.C. Interferon-gamma genetic polymorphism and expression in Kawasaki disease. Medicine 2016, 95, e3501. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| HD | TB | p-value | ||
|---|---|---|---|---|
| N | 175 | 201 | ||
| Age (years) | 34.25 ± 1.16 | 33.03 ± 0.98 | 0.123 a | |
| Ethnicity | Caucasian | 65.3% | 63.9% | 0.896 b |
| American Indian | 34.7% | 36.1% | ||
| Sex | Male | 36.6% | 72.9% | <0.001 b |
| Female | 63.4% | 27.1% | ||
| rs1861494 Genotypes | HD (N = 175) | TB (N = 201) | |||||
|---|---|---|---|---|---|---|---|
| GG | GA | AA | GG | GA | AA | ||
| Ethnicity | Caucasian | 9 (7.89%) | 62 (54.39%) | 43 (37.72%) | 20 (15.63%) | 59 (46.09%) | 49 (38.28%) |
| American Indian | 6 (9.84%) | 30 (49.18%) | 25 (40.98%) | 17 (23.29%) | 34 (46.58%) | 22 (30.14%) | |
| p-value | 0.783 | 0.308 | |||||
| Sex | Male | 7 (11.11%) | 29 (46.03%) | 27 (42.86%) | 32 (22.07%) | 71 (48.97%) | 42 (28.96%) |
| Female | 11 (9.82%) | 48 (42.86%) | 53 (47.32%) | 8 (14.29%) | 26 (46.43%) | 22 (39.28%) | |
| p-value | 0.847 | 0.268 | |||||
| rs1861494 SNP Genotypes | p-value | |||
|---|---|---|---|---|
| GG | AG | AA | ||
| HD (N = 21) | ||||
| IFNG levels in plasma (pg/mL) | 13.59 (±5.78) | 9.44 (±1.64) | 15.33 (±6.72) | 0.797 |
| IFNG levels in culture supernatants (pg/mL) | 5185 (±1491) | 14,195 (±3835) | 20,989 (±5218) | 0.034 |
| % CD4+ T cells IFNG+ | 9.06 (±2.08) | 15.47 (±3.66) | 20.44 (±2.48) | 0.035 |
| TB (N = 21) | ||||
| IFNG levels in plasma (pg/mL) | 67.27 (±59.21) | 46.31 (±20.25) | 170.5 (±68.92) | 0.144 |
| IFNG levels in culture supernatants (pg/mL) | 6772 (±2750) | 7575 (±2011) | 15,066 (±4553) | 0.357 |
| % CD4+ T cells IFNG+ | 7.91 (±1.38) | 12.39 (±4.99) | 15.97 (±4.29) | 0.809 |
| rs1861494 SNP Genotypes | p-value | |||
|---|---|---|---|---|
| GG | AG | AA | ||
| Hematologic Studies (N = 105) | ||||
| Leucocytes (cells/mL) | 10,754 (±1557) | 10,205 (±388.8) | 9929 (±702.1) | 0.432 a |
| Lymphocytes (cells/mL) | 1964 (±349.5) | 1491 (±76.45) | 1573 (±126.5) | 0.762 a |
| Monocytes (cells/mL) | 1013 (±138.0) | 809.3 (±58.38) | 878.6 (±83.13) | 0.584 a |
| Neutrophils (cells/mL) | 8097 (±1557) | 7524 (±586.6) | 5674 (±574.4) | 0.089 a |
| AFB in sputum smear (N = 150) | ||||
| BAAR− or BAAR+ | 14 (18.0%) | 43 (55.1%) | 21 (26.9%) | 0.415 b |
| BAAR++ or BAAR+++ | 17 (23.6%) | 32 (44.4%) | 23 (32.0%) | |
| Radiological Lesions (N = 136) | ||||
| Mild or Moderate | 11 (21.1%) | 29 (55.8%) | 12 (23.1%) | 0.367 b |
| Severe | 17 (20.3%) | 39 (46.4%) | 28 (33.3%) | |
| Months of disease progression (N = 105) | 2.15 (±0.29) | 2.92 (±3.56) | 2.29 (±0.35) | 0.447 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rolandelli, A.; Pellegrini, J.M.; Amiano, N.O.; Santilli, M.C.; Morelli, M.P.; Castello, F.A.; Tateosian, N.L.; Levi, A.; Casco, N.; Palmero, D.J.; et al. The IFNG rs1861494 Single Nucleotide Polymorphism Is Associated with Protection against Tuberculosis Disease in Argentina. Genes 2018, 9, 46. https://doi.org/10.3390/genes9010046
Rolandelli A, Pellegrini JM, Amiano NO, Santilli MC, Morelli MP, Castello FA, Tateosian NL, Levi A, Casco N, Palmero DJ, et al. The IFNG rs1861494 Single Nucleotide Polymorphism Is Associated with Protection against Tuberculosis Disease in Argentina. Genes. 2018; 9(1):46. https://doi.org/10.3390/genes9010046
Chicago/Turabian StyleRolandelli, Agustín, Joaquín M. Pellegrini, Nicolás O. Amiano, María C. Santilli, María P. Morelli, Florencia A. Castello, Nancy L. Tateosian, Alberto Levi, Nicolás Casco, Domingo J. Palmero, and et al. 2018. "The IFNG rs1861494 Single Nucleotide Polymorphism Is Associated with Protection against Tuberculosis Disease in Argentina" Genes 9, no. 1: 46. https://doi.org/10.3390/genes9010046
APA StyleRolandelli, A., Pellegrini, J. M., Amiano, N. O., Santilli, M. C., Morelli, M. P., Castello, F. A., Tateosian, N. L., Levi, A., Casco, N., Palmero, D. J., & García, V. E. (2018). The IFNG rs1861494 Single Nucleotide Polymorphism Is Associated with Protection against Tuberculosis Disease in Argentina. Genes, 9(1), 46. https://doi.org/10.3390/genes9010046