Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Heat-Shock Proteins 70 in Plant Species
2.2. Alignments and Phylogenetic Analyses
2.3. Motif Search, Gene Structure and Synteny Analysis
2.4. Plant Materials, Stress Treatment and Gene Expression Analyses
3. Results
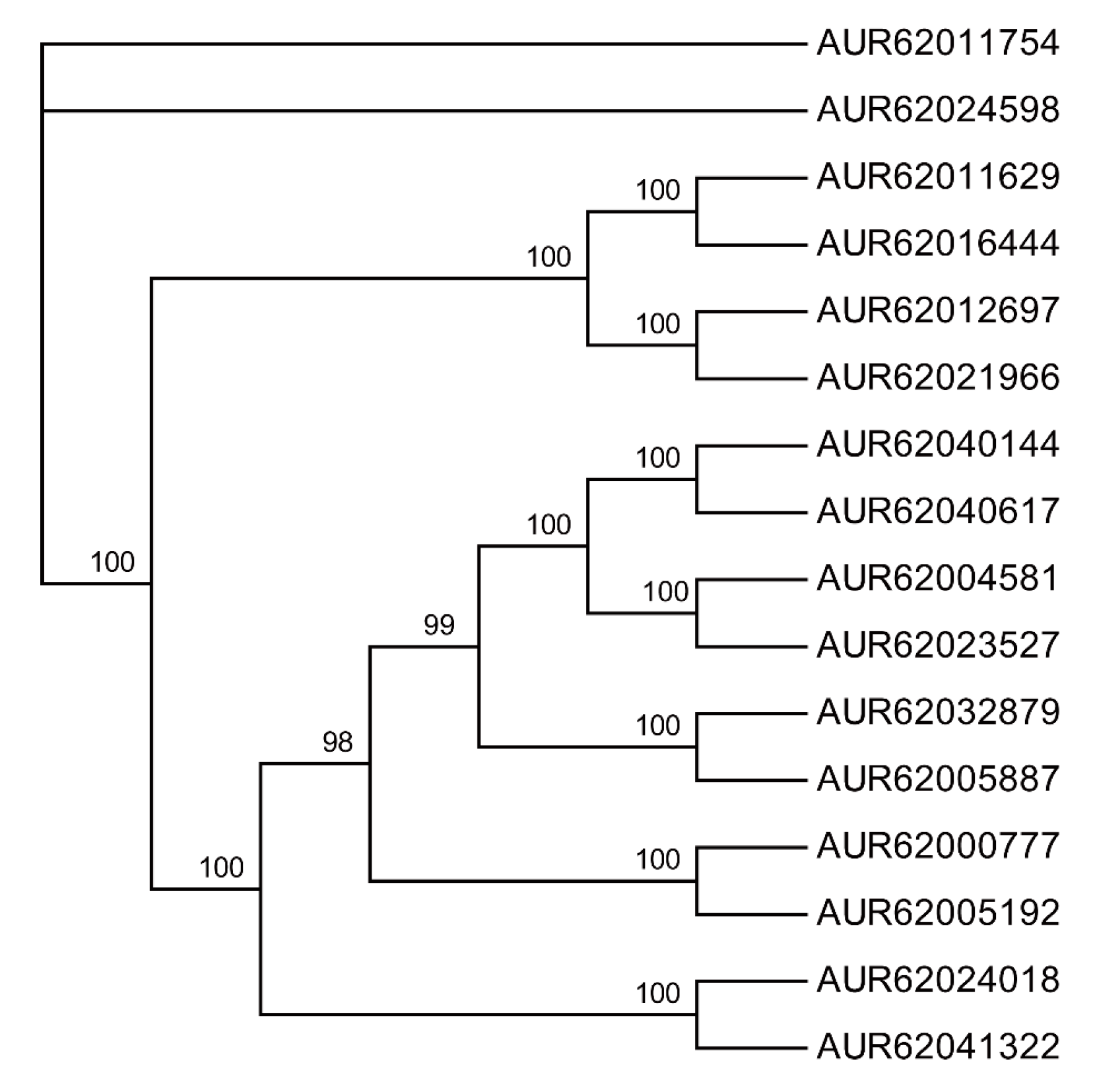
3.1. Identification of HSP70 Proteins in Quinoa Proteome
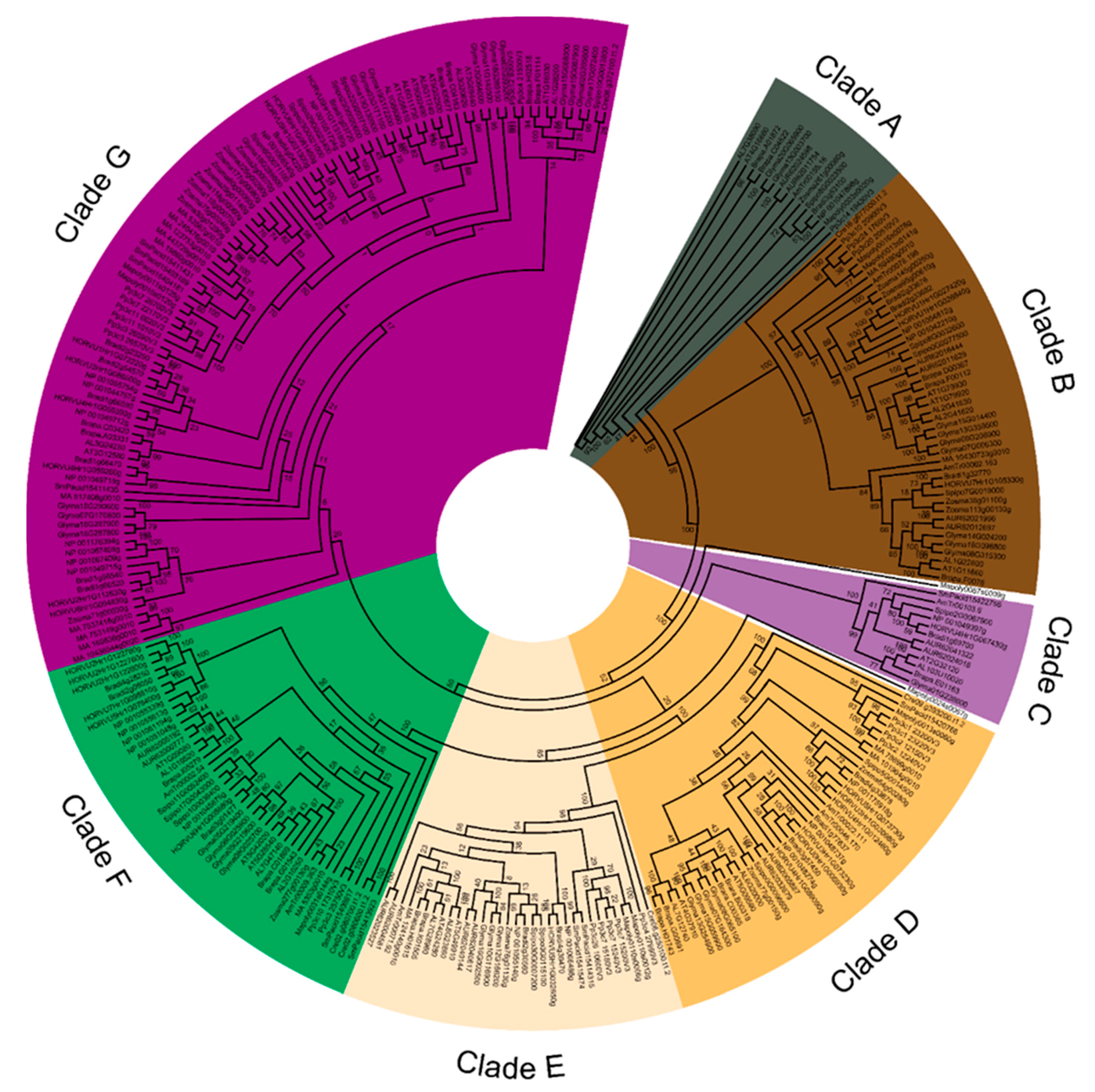
3.2. The Land Plant HSP70 Superfamily Splits into at Least Seven Phylogenetic Clades
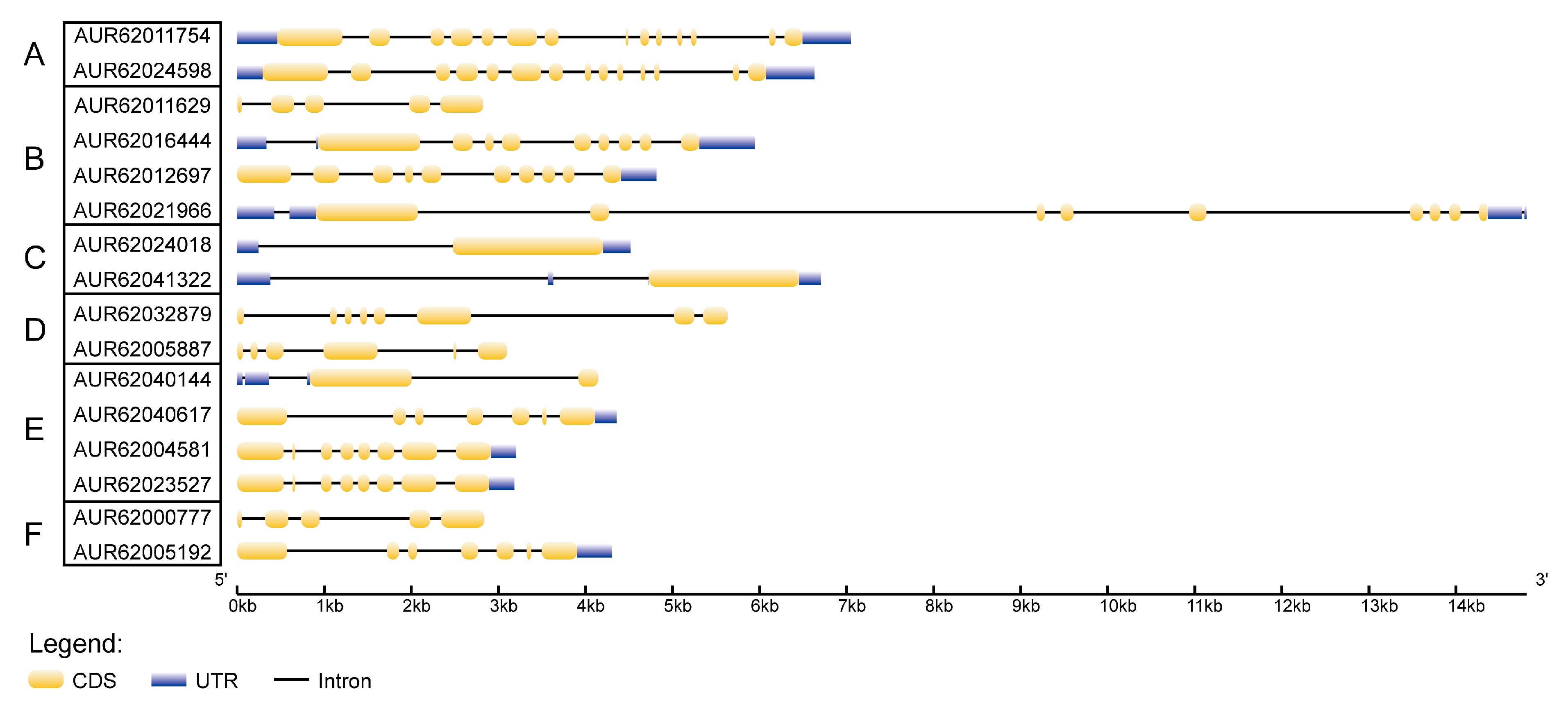
3.3. Gene Structure and Conserved Motif Analyses of Hsp70s and Their Encoding Proteins in Quinoa
3.4. Allopolyploidy Event Contributed to the Expansion of Cqhsp70s
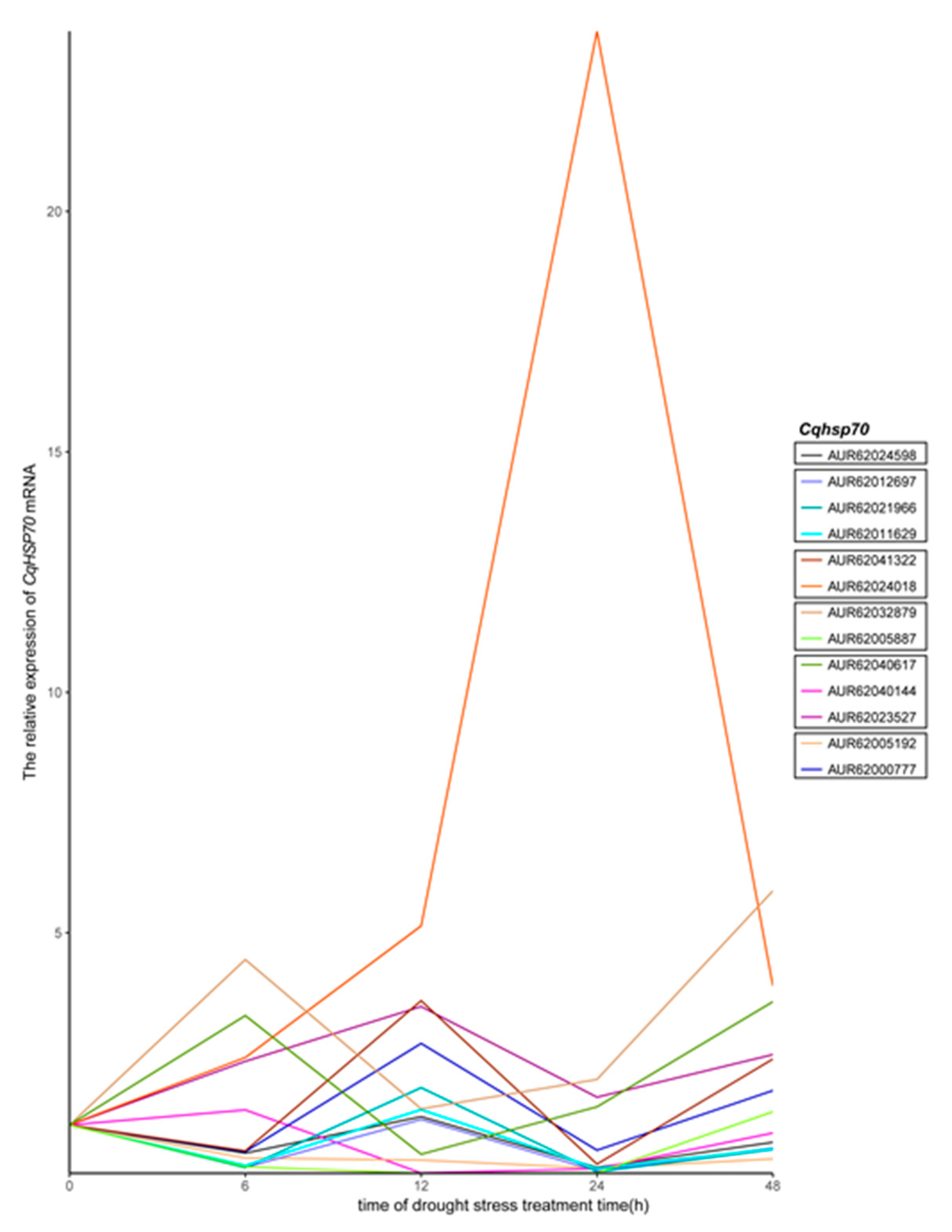
3.5. Cqhsp70 Genes in Quinoa Are Responsive to Drought Stress
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, W.X.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef] [PubMed]
- Moser, D.; Doumbo, O.; Klinkert, M.Q. The humoral response to heat-shock protein-70 in human and Murine schistosomiasis-mansoni. Parasite Immunol. 1990, 12, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Vierling, E. The roles of heat-shock proteins in plants. Annu. Rev. Plant Phys. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Tripp, J.; Mishra, S.K.; Scharf, K.D. Functional dissection of the cytosolic chaperone network in tomato mesophyll protoplasts. Plant Cell Environ. 2009, 32, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.P.; Kaneko, Y.; Subjeck, J.R. The Hsp110 and Grp1 70 stress proteins: Newly recognized relatives of the Hsp70s. Cell Stress Chaperones 2000, 5, 276–290. [Google Scholar] [CrossRef]
- Flaherty, K.M.; Delucaflaherty, C.; Mckay, D.B. Three-dimensional structure of the ATPase fragment of a 70k heat-shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.T.; Zhao, X.; Burkholder, W.F.; Gragerov, A.; Ogata, C.M.; Gottesman, M.E.; Hendrickson, W.A. Structural analysis of substrate binding by the molecular chaperone DnaK. Science 1996, 272, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Golding, G.B. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. J. Mol. Evol. 1993, 37, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Munro, S.; Pelham, H.R.B. An hsp70-like protein in the ER: Identity with the 78 kd glucose-regulated protein and immunoglobulin heavy-chain binding-protein. Cell 1986, 46, 291–300. [Google Scholar] [CrossRef]
- Engman, D.M.; Kirchhoff, L.V.; Donelson, J.E. Molecular cloning of mtp70, a mitochondrial member of the hsp70 family. Mol. Cell. Biol. 1989, 9, 5163–5168. [Google Scholar] [CrossRef] [PubMed]
- Leustek, T.; Dalie, B.; Amirshapira, D.; Brot, N.; Weissbach, H. A member of the hsp70 family is localized in mitochondria and resembles Escherichia coli DnaK. Proc. Natl. Acad. Sci. USA 1989, 86, 7805–7808. [Google Scholar] [CrossRef] [PubMed]
- De Maio, A.; Vazquez, D. Extracellular heat shock proteins: A new location, a new function. Shock 2013, 40, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.L.; Wang, J.S.; Liu, H.C.; Chen, R.W.; Meyer, Y.; Barakat, A.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201–208. [Google Scholar] [CrossRef]
- Sarkar, N.K.; Kundnani, P.; Grover, A. Functional analysis of Hsp70 superfamily proteins of rice (Oryza sativa). Cell Stress Chaperones 2013, 18, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Yu, A.M.; Li, P.; Yang, H.; Liu, G.J.; Liu, L. Sequence analysis of the Hsp70 family in moss and evaluation of their functions in abiotic stress responses. Sci. Rep.-UK 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmockel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.A.; Nishiyama, T.; Hasebe, M.; Bowman, J.L.; Gribskov, M.; dePamphilis, C.; Albert, V.A.; Aono, N.; Aoyama, T.; Ambrose, B.A.; et al. The Selaginella genome identifies genetic changes associated with the evolution of vascular plants. Science 2011, 332, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Haberer, G.; Gundlach, H.; Gläßer, C.; Nussbaumer, T.; Luo, M.C.; Lomsadze, A.; Borodovsky, M.; Kerstetter, R.A.; Shanklin, J.; et al. The Spirodela polyrhiza genome reveals insights into its neotenous reduction fast growth and aquatic lifestyle. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.Y.; Li, W.W.; Noble, W.S. MEME suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Small, I.; Peeters, N.; Legeai, F.; Lurin, C. Predotar: A tool for rapidly screening proteomes for N-terminal targeting sequences. Proteomics 2004, 4, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Tang, H.B.; DeBarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.; Zurita-Silva, A.; Maldonado, J.; Silva, H. Transcriptional responses of Chilean quinoa (Chenopodium quinoa Willd.) under water deficit conditions uncovers ABA-independent expression patterns. Front. Plant Sci. 2017, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Molina-Montenegro, M.A.; Oses, R.; Torres-Díaz, C.; Atala, C.; Zurita-Silva, A.; Ruiz-Lara, S. Root-endophytes improve the ecophysiological performance and production of an agricultural species under drought condition. AoB Plants 2016, 8, plw062. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Carrasco, K.; Antognoni, F.; Coulibaly, A.K.; Lizardi, S.; Covarrubias, A.; Martínez, E.A.; Molina-Montenegro, M.A.; Biondi, S.; Zurita-Silva, A. Variation in salinity tolerance of four lowland genotypes of quinoa (Chenopodium quinoa Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. 2011, 49, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Amborella Genome, P. The Amborella genome and the evolution of flowering plants. Science 2013, 342, 1241089. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.X.; Theg, S.M. A stromal heat shock protein 70 system functions in protein import into chloroplasts in the moss Physcomitrella patens. Plant Cell 2010, 22, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Jungkunz, I.; Link, K.; Vogel, F.; Voll, L.M.; Sonnewald, S.; Sonnewald, U. AtHsp70-15-deficient Arabidopsis plants are characterized by reduced growth, a constitutive cytosolic protein response and enhanced resistance to TuMV. Plant J. 2011, 66, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yun, H.S.; Kwon, C. Molecular communications between plant heat shock responses and disease resistance. Mol. Cells 2012, 34, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.S.; Chen, A.J.; Xiao, L.H.; Muller, H.M.; Ache, P.; Haberer, G.; Zhang, M.L.; Jia, W.; Deng, P.; Huang, R.; et al. A high-quality genome assembly of quinoa provides insights into the molecular basis of salt bladder-based salinity tolerance and the exceptional nutritional value. Cell Res. 2017, 27, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.L.; Rouze, P.; Verhelst, B.; Lin, Y.C.; Bayer, T.; Collen, J.; Dattolo, E.; De Paoli, E.; Dittami, S.; Maumus, F.; et al. The genome of the seagrass Zostera marina reveals angiosperm adaptation to the sea. Nature 2016, 530, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, F.U.; Hayer-Hartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.T.; Pfanner, N. Hsp70 proteins in protein translocation. Adv. Protein Chem. 2001, 59, 223–242. [Google Scholar] [PubMed]
- Pratt, W.B.; Toft, D.O. Regulation of signaling protein function and trafficking by the Hsp90/Hsp70-based chaperone machinery. Exp. Biol. Med. 2003, 228, 111–133. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Number | Phylogenetic Class | Other | ||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | |||
| Chlamydomonas reinhardtii | 6 | 0 | 1 | 0 | 1 | 1 | 2 | 1 | |
| Metrosideros polymorpha | 11 | 1 | 2 | 0 | 1 | 2 | 1 | 2 | 2 |
| Physcomitrella patens | 23 | 1 | 3 | 0 | 4 | 5 | 2 | 8 | |
| Selaginella moellendorffii | 10 | 0 | 0 | 1 | 1 | 2 | 1 | 4 | 1 |
| Picea abies | 16 | 0 | 2 | 0 | 2 | 1 | 1 | 10 | |
| Amborella trichopoda | 9 | 1 | 2 | 1 | 2 | 1 | 2 | ||
| Spirodela polyrhiza | 18 | 1 | 3 | 1 | 2 | 2 | 3 | 6 | |
| Zostera marina | 19 | 1 | 4 | 0 | 2 | 1 | 1 | 10 | |
| Oryza sativa | 24 | 1 | 2 | 1 | 3 | 2 | 5 | 10 | |
| Aponogeton distachyos | 21 | 1 | 3 | 1 | 3 | 2 | 3 | 8 | |
| Hordeum vulgare | 26 | 0 | 3 | 1 | 6 | 1 | 6 | 9 | |
| Chenopodium quinoa | 16 | 2 | 4 | 2 | 2 | 4 | 2 | 0 | |
| Glycine max | 37 | 2 | 7 | 1 | 4 | 3 | 4 | 16 | |
| Brassica rapa | 21 | 2 | 3 | 1 | 4 | 2 | 3 | 6 | |
| Arabidopsis lyrata | 18 | 1 | 3 | 1 | 2 | 2 | 3 | 6 | |
| Arabidopsis thaliana | 18 | 1 | 3 | 1 | 2 | 2 | 3 | 6 | |
| HSP70s in C.quinoa | HSP70s in A.thaliana | |||
|---|---|---|---|---|
| Quinoa proteome ID | GeneBank ID | TAIR ID | GeneBank ID | similar expression pattern |
| AUR62024598 | XP_021768679.1 | AT4G16660 | NP_567510.1 | yes |
| AUR62011754 | XP_021769992.1 | AT4G16660 | NP_567510.1 | - |
| AUR62012697 | XP_021719698.1 | AT1G11660 | NP_172631.2 | no |
| AUR62021966 | XP_021766890.1 | AT1G11660 | NP_172631.2 | no |
| AUR62011629 | XP_021731972.1 | AT1G79930 | NP_178111.1 | yes |
| AUR62016444 | XP_021736828.1 | AT1G79930 | NP_178111.1 | - |
| AUR62041322 | XP_021729859.1 | AT2G32120 | NP_180771.1 | yes |
| AUR62024018 | XP_021732896.1 | AT2G32120 | NP_180771.1 | no |
| AUR62032879 | XP_021747611.1 | AT5G09590 | NP_196521.1 | no |
| AUR62005887 | XP_021747614.1 | AT5G09590 | NP_196521.1 | yes |
| AUR62040617 | XP_021743068.1 | AT5G49910 | NP_199802.1 | yes |
| AUR62040144 | XP_021719138.1 | AT5G49910 | NP_199802.1 | yes |
| AUR62023527 | XP_021760397.1 | AT5G49910 | NP_199802.1 | yes |
| AUR62004581 | XP_021748808.1 | AT5G49910 | NP_199802.1 | yes |
| AUR62005192 | XP_021742552.1 | AT5G42020 | NP_851119.1 | yes |
| AUR62000777 | XP_021772891.1 | AT5G42020 | NP_851119.1 | no |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wang, R.; Liu, W.; Zhang, H.; Guo, Y.; Wen, R. Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress. Genes 2018, 9, 35. https://doi.org/10.3390/genes9020035
Liu J, Wang R, Liu W, Zhang H, Guo Y, Wen R. Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress. Genes. 2018; 9(2):35. https://doi.org/10.3390/genes9020035
Chicago/Turabian StyleLiu, Jianxia, Runmei Wang, Wenying Liu, Hongli Zhang, Yaodong Guo, and Riyu Wen. 2018. "Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress" Genes 9, no. 2: 35. https://doi.org/10.3390/genes9020035
APA StyleLiu, J., Wang, R., Liu, W., Zhang, H., Guo, Y., & Wen, R. (2018). Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress. Genes, 9(2), 35. https://doi.org/10.3390/genes9020035