Characterization of the Symbiotic Nitrogen-Fixing Common Bean Low Phytic Acid (lpa1) Mutant Response to Water Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Analysis of Symbiotic Phenotype
2.3. Statistical Analysis
2.4. Real-Time Quantitative PCR Analysis
3. Results
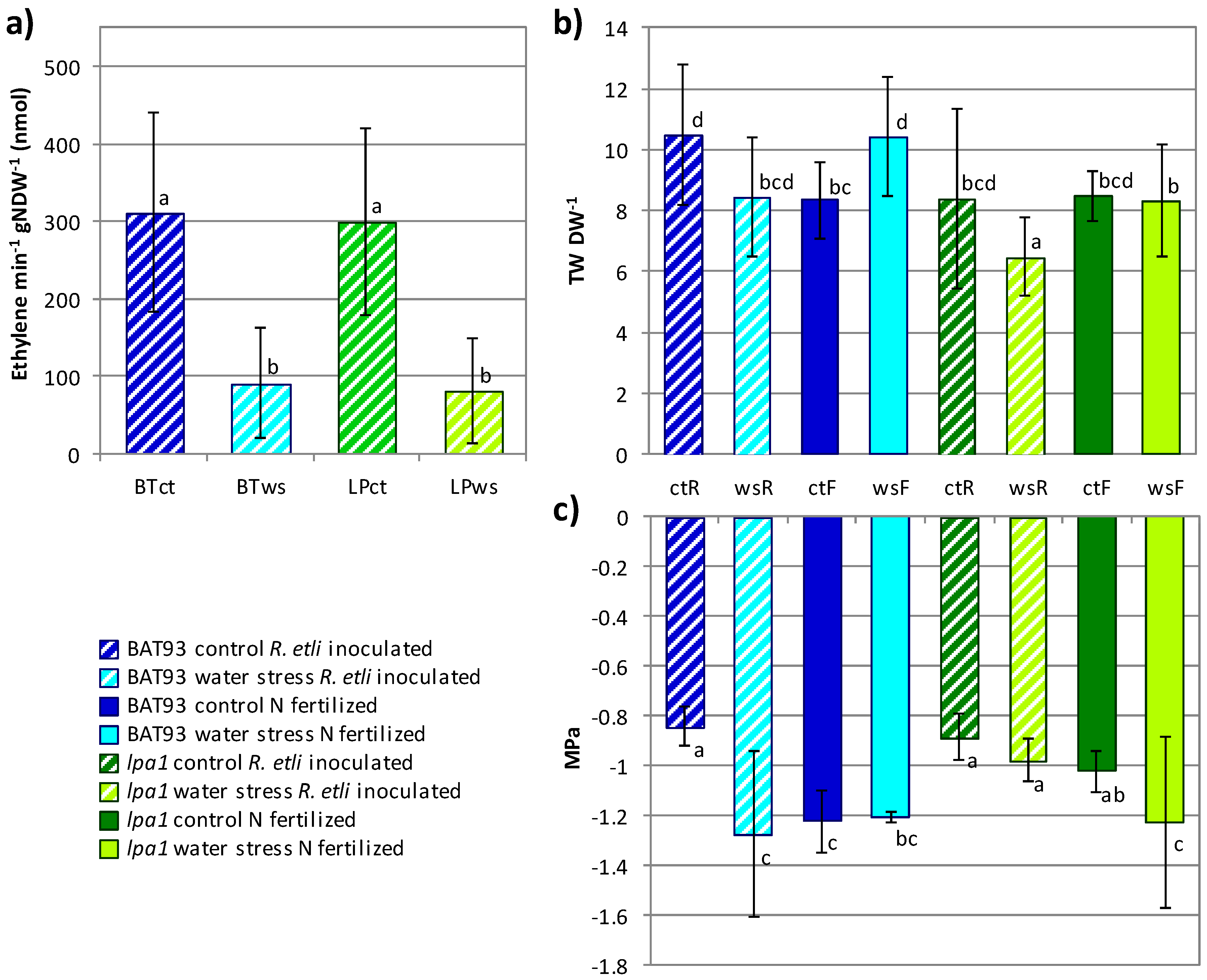
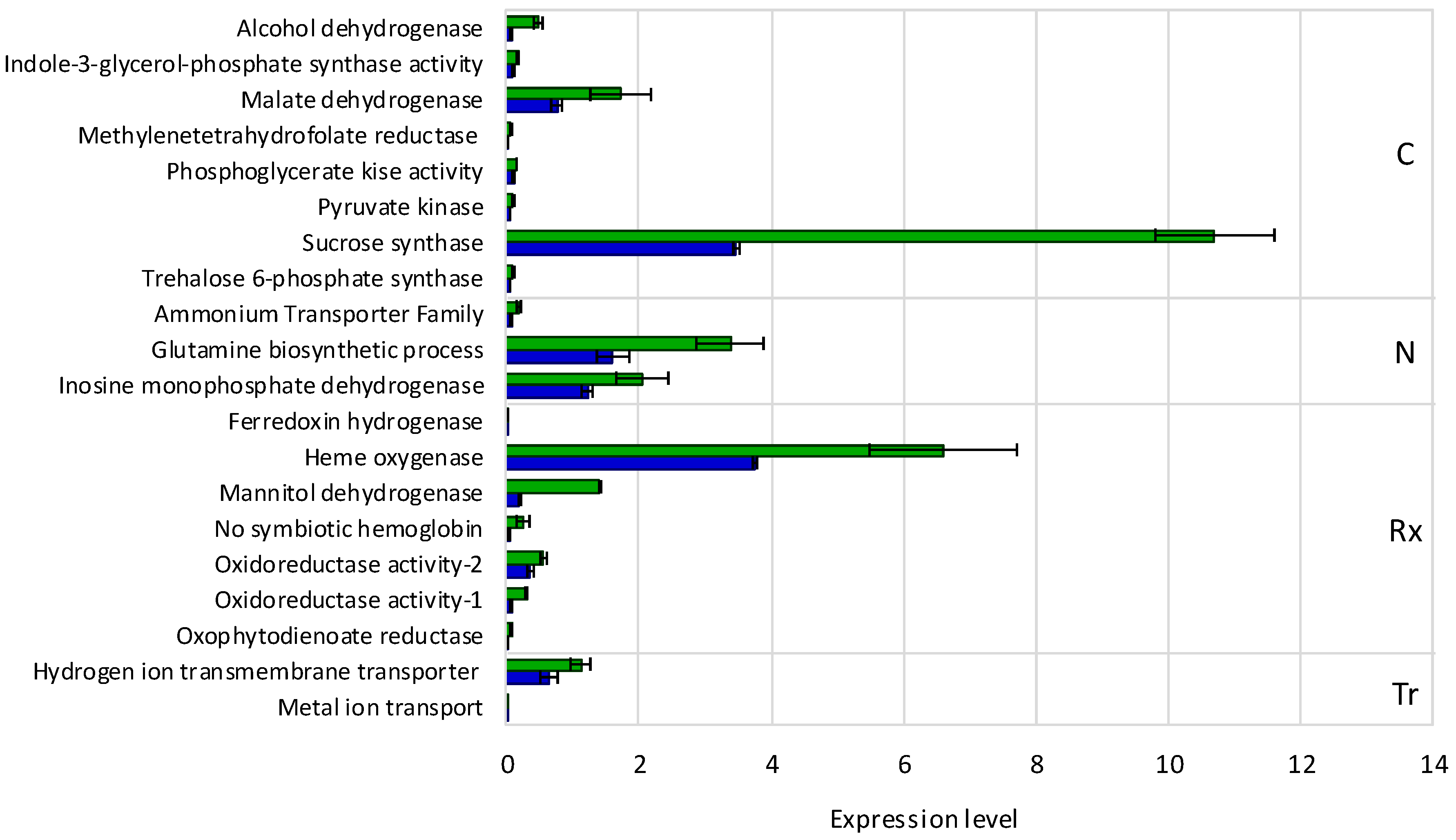
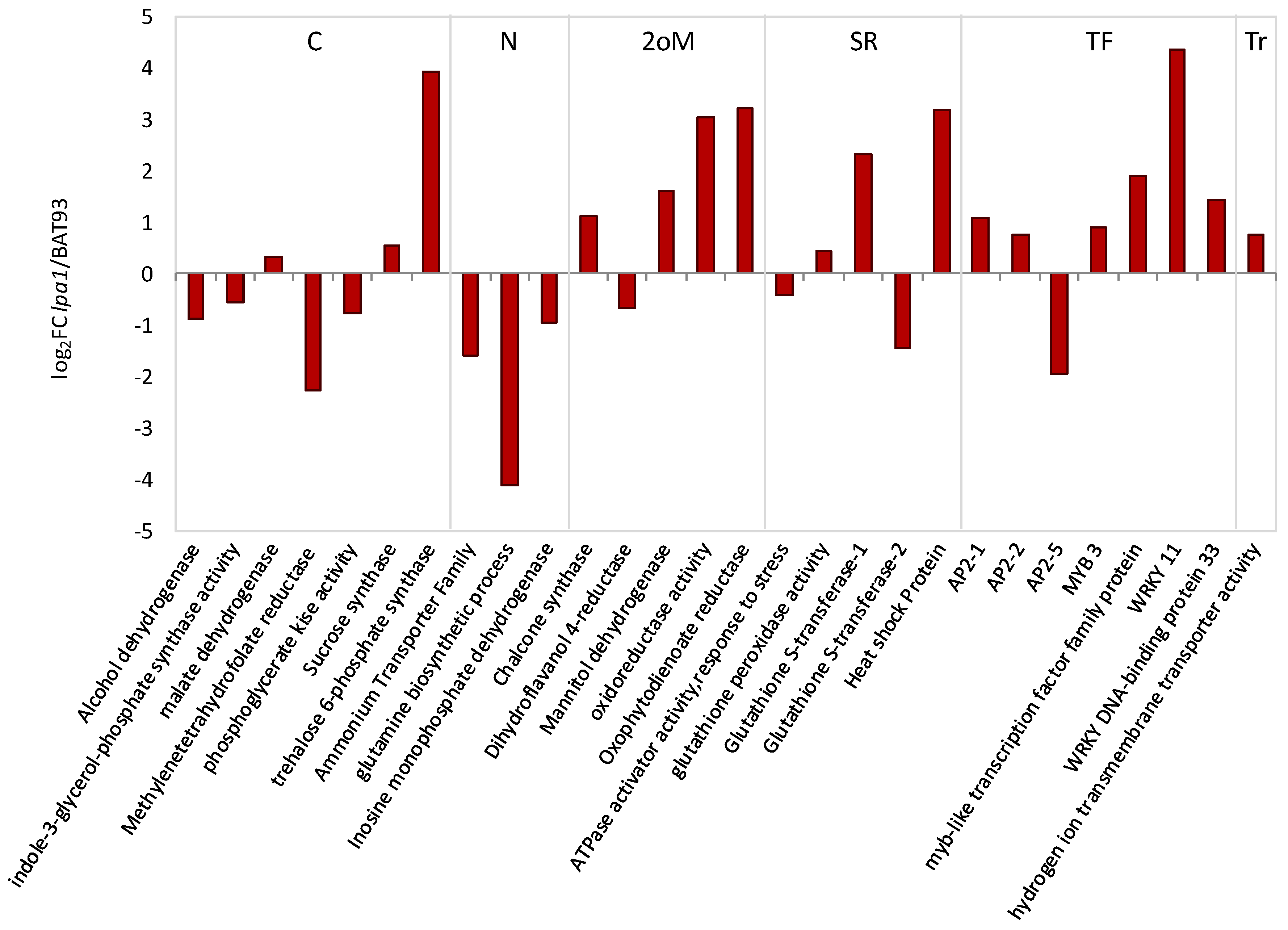
3.1. The Symbiotic Nitrogen Fixation Phenotype and Nodule Gene Expression Profile of lpa1 Plants
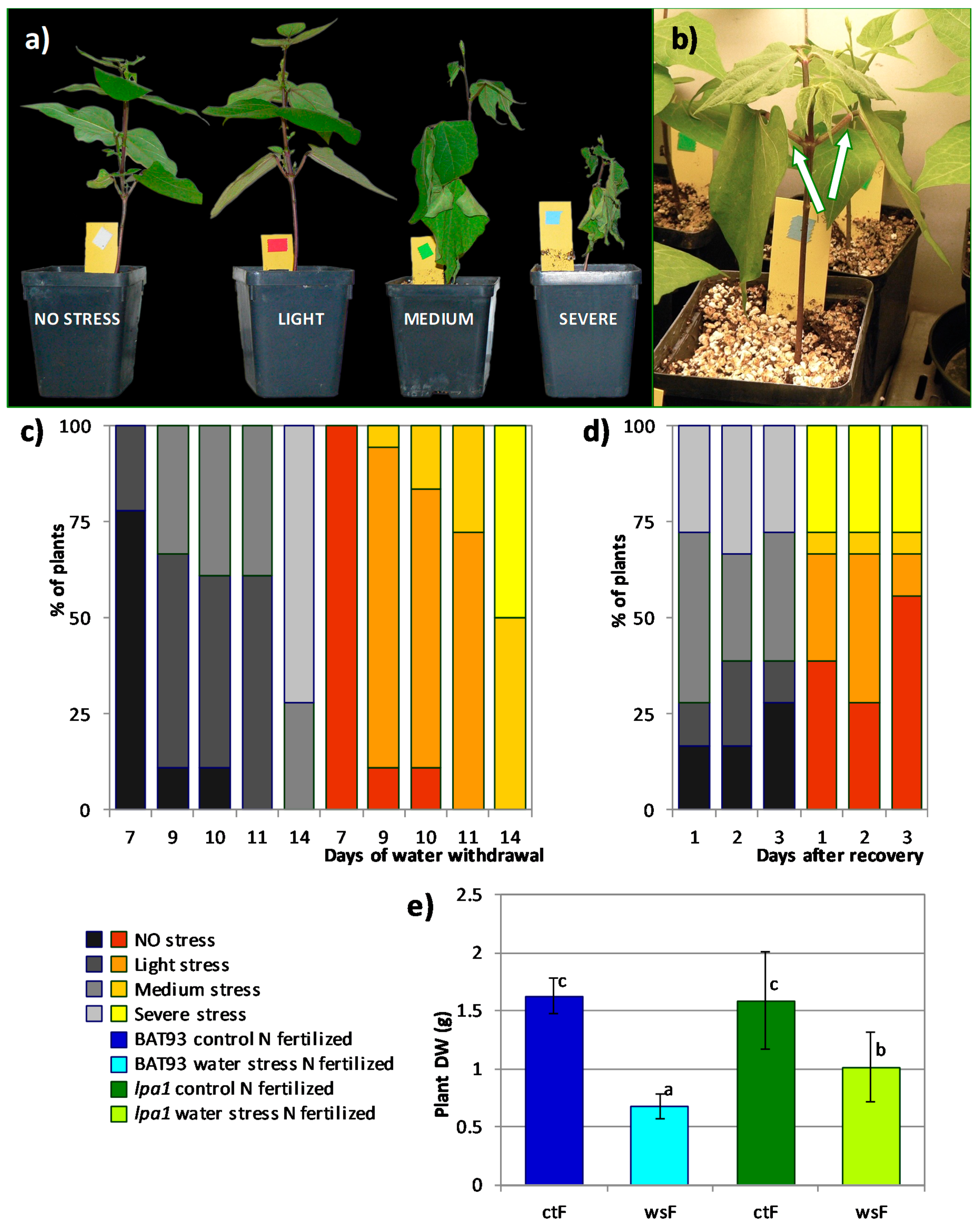
3.2. Comparative Analysis of the Effect of Water Stress and the Response to Rehydration Recovery in lpa1 vs. Wild Type Fertilized Plants
3.3. Comparative Analysis of the Effect of Water Stress in lpa1 vs. BAT 93 Symbiotic Nitrogen Fixation Plants
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)–model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.P.; Silva, H.; Ledent, J.F.; Pinto, M. Effect of drought stress on the osmotic adjustment, cell wall elasticity and cell volume of six cultivars of common beans (Phaseolus vulgaris L.). Eur. J. Agron. 2007, 26, 30–38. [Google Scholar] [CrossRef]
- Terán, H.; Singh, S.P. Comparison of sources and lines selected for drought resistance in common bean. Crop Sci. 2002, 42, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Kirova, E.; Tzvetkova, N.; Vaseva, I.; Ignatov, G. Photosynthetic responses of nitrate-fed and nitrogen-fixing soybeans to progressive water stress. J. Plant Nutr. 2008, 31, 445–458. [Google Scholar] [CrossRef]
- Lodeiro, A.R.; Gonzàlez, P.; Hernàndez, A.; Balaguè, L.J.; Favelukes, G. Comparison of drought tolerance in nitrogen-fixing and inorganic nitrogen-grown common beans. Plant Sci. 2000, 154, 31–41. [Google Scholar] [CrossRef]
- Arrese-Igor, C.; González, E.; Marino, D.; Ladrera, R.; Larrainzar, E.; Gil-Quintana, E. Physiological response of legume nodules to drought. Plant Stress 2011, 5, 24–31. [Google Scholar]
- Mohammadi, K.; Sohrabi, Y.; Heidari, G.; Khalesro, S.; Majidi, M. Effective factors on biological nitrogen fixation. Afr. J. Agric. Res. 2012, 7, 1782–1788. [Google Scholar] [CrossRef]
- González, E.M.; Aparicio-Tejo, P.M.; Gordon, A.J.; Minchin, F.R.; Royuela, M.; Arrese-Igor, C. Water-deficit effects on carbon and nitrogen metabolism of pea nodules. J. Exp. Bot. 1998, 49, 1705–1714. [Google Scholar] [CrossRef]
- Serraj, R. Effects of drought stress on legume symbiotic nitrogen fixation: Physiological mechanisms. Indian J. Exp. Biol. 2003, 41, 1136–1141. [Google Scholar] [PubMed]
- Gálvez, L.; González, E.M.; Arrese-Igor, C. Evidence for carbon flux shortage and strong carbon/nitrogen interactions in pea nodules at early stages of water stress. J. Exp. Bot. 2005, 56, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Marino, D.; Hohnjec, N.; Küster, H.; Moran, J.F.; González, E.M.; Arrese-Igor, C. Evidence for transcriptional and post-translational regulation of sucrose synthase in pea nodules by the cellular redox state. Mol. Plant Microbe Interact. 2008, 21, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Harvest Plus. Available online: http://www.harvestplus.org/what-we-do/crops.
- Petry, N.; Egli, I.; Campion, B.; Nielsen, E.; Hurrell, R. Genetic reduction of phytate in common bean (Phaseolus vulgaris L.) seeds increases iron absorption in young women. J. Nutr. 2013, 143, 1219–1224. [Google Scholar] [CrossRef] [PubMed]
- Raboy, V. Approaches and challenges to engineering seed phytate and total phosphorus. Plant Sci. 2009, 177, 281–296. [Google Scholar] [CrossRef]
- Campion, B.; Glahn, R.P.; Tava, A.; Perrone, D.; Doria, E.; Sparvoli, F. Genetic reduction of antinutrients in common bean (Phaseolus vulgaris L.) seed, increases nutrients and in vitro iron bioavailability without depressing main agronomic traits. Field Crops Res. 2013, 141, 27–37. [Google Scholar] [CrossRef]
- Campion, B.; Sparvoli, F.; Doria, E.; Tagliabue, G.; Galasso, I.; Fileppi, M. Isolation and characterisation of an lpa (low phytic acid) mutant in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2009, 118, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Panzeri, D.; Cassani, E.; Doria, E.; Tagliabue, G.; Forti, L.; Campion, B. A defective ABC transporter of the MRP family, responsible for the bean lpa1 mutation, affects the regulation of the phytic acid pathway, reduces seed myo-inositol and alters ABA sensitivity. New Phytol. 2011, 191, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cominelli, E.; Confalonieri, M.; Carlessi, M.; Cortinovis, G.; Daminati, M.G.; Porch, T.G.; Losa, A.; Sparvoli, F. Phytic acid transport in Phaseolus vulgaris: A new low phytic acid mutant in the PvMRP1 gene and study of the PvMRPs promoters in two different plant systems. Plant Science 2018, 270, 1–12. [Google Scholar]
- Sparvoli, F.; Cominelli, E. Seed biofortification and phytic acid reduction: A conflict of interest for the plant? Plants 2015, 4, 728–755. [Google Scholar] [CrossRef] [PubMed]
- Bregitzer, P.; Raboy, V. Effects of four independent low-phytate mutations on barley agronomic performance. Crop Sci. 2006, 46, 1318–1322. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Peterson, K.M.; Souza, E.J. Mineral distributions in milling fractions of low phytic acid wheat. Crop Sci. 2006, 46, 2692–2698. [Google Scholar] [CrossRef]
- Pilu, R.; Landoni, M.; Cassani, E.; Doria, E.; Nielsen, E. The maize lpa241 mutation causes a remarkable variability of expression and some pleiotropic effects. Crop Sci. 2005, 45, 2096–2105. [Google Scholar] [CrossRef]
- Klein, M.; Perfus-Barbeoch, L.; Frelet, A.; Gaedeke, N.; Reinhardt, D.; Mueller-Roeber, B. The plant multidrug resistance ABC transporter AtMRP5 is involved in guard cell hormonal signalling and water use. Plant J. 2003, 33, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Hitz, W.D.; Carlson, T.J.; Kerr, P.S.; Sebastian, S.A. Biochemical and molecular characterization of a mutation that confers a decreased raffinosaccharide and phytic acid phenotype on soybean seeds. Plant Physiol. 2002, 128, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, J.R.; Premachandra, G.S.; Young, K.A.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci. 2000, 40, 1601–1605. [Google Scholar] [CrossRef]
- Yuan, F.-J.; Zhao, H.-J.; Ren, X.-L.; Zhu, S.-L.; Fu, X.-J.; Shu, Q.-Y. Generation and characterization of two novel low phytate mutations in soybean (Glycine max L. Merr.). Theor. Appl. Genet. 2007, 115, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Shunmugam, A.S.K.; Bock, C.; Arganosa, G.C.; Georges, F.; Gray, G.R.; Warkentin, T.D. Accumulation of phosphorus-containing compounds in developing seeds of low-phytate pea (Pisum sativum L.) mutants. Plants 2015, 4, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.; Capella-Gutiérrez, S.; Rendón-Anaya, M.; Hernández-Oñate, M.; Minoche, A.E.; Erb, I. Genome and transcriptome analysis of the Mesoamerican common bean and the role of gene duplications in establishing tissue and temporal specialization of genes. Genome Biol. 2016, 17, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerfield, R.J.; Huxley, P.A.; Minchin, F.R. Plant husbandry and management techniques for growing grain legumes under simulated tropical conditions in controlled environments. Exp. Agric. 1977, 13, 81–92. [Google Scholar] [CrossRef]
- Hardy, R.W.F.; Holsten, R.D.; Jackson, E.K.; Burns, R.C. The acetylene-ethylene assay for N2 fixation: Laboratory and field evaluation. Plant Physiol. 1968, 43, 1185–1207. [Google Scholar] [CrossRef] [PubMed]
- Rosales, M.A.; Cuellar-Ortiz, S.M.; de la Paz Arrieta-Montiel, M.; Acosta-Gallegos, J.; Covarrubias, A.A. Physiological traits related to terminal drought resistance in common bean (Phaseolus vulgaris L.). J. Sci. Food Agric. 2013, 93, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Castro-Camus, E.; Palomar, M.; Covarrubias, A.A. Leaf water dynamics of Arabidopsis thaliana monitored in-vivo using terahertz time-domain spectroscopy. Sci. Rep. 2013, 3, 2910. [Google Scholar] [CrossRef] [PubMed]
- Hernández, G.; Ramírez, M.; Valdés-López, O.; Tesfaye, M.; Graham, M.A.; Czechowski, T. Phosphorus stress in common bean: Root transcript and metabolic responses. Plant Physiol. 2007, 144, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Guillén, G.; Fuentes, S.I.; Íñiguez, L.P.; Aparicio-Fabre, R.; Zamorano-Sánchez, D. Transcript profiling of common bean nodules subjected to oxidative stress. Physiol. Plant. 2013, 149, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Íñiguez, L.P.; Guerrero, G.; Sparvoli, F.; Hernández, G. Rhizobium etli bacteroids engineered for Vitreoscilla hemoglobin expression alleviate oxidative stress in common bean nodules that reprogramme global gene expression. Plant. Biotechnol. Rep. 2016, 10, 463–474. [Google Scholar] [CrossRef]
- Suárez, R.; Wong, A.; Ramírez, M.; Barraza, A.; Orozco, M.D.C.; Cevallos, M.A. Improvement of drought tolerance and grain yield in common bean by overexpressing trehalose-6-phosphate synthase in rhizobia. Mol. Plant Microbe Interact. 2008, 21, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.; Brom, S.; Dàvalos, A.; Lòpez, O.; Soberòn, M.; Romero, D. Differential regulation of fixN-reiterated genes in Rhizobium etli by a novel fixL—fixK cascade. Mol. Plant Microbe Interact. 2000, 13, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, R.; Tongoona, P.; Derera, J.; Laing, M.; Watson, G. Combining ability of low phytic acid (lpa1-1) and quality protein maize (QPM) lines for seed germination and vigour under stress and non-stress conditions. Euphytica 2012, 185, 529–541. [Google Scholar] [CrossRef]
- Cerino Badone, F.; Amelotti, M.; Cassani, E.; Pilu, R. Study of low phytic acid1-7 (lpa1-7), a new ZmMRP4 mutation in maize. J. Hered. 2012, 103, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Nagy, R.; Grob, H.; Weder, B.; Green, P.; Klein, M.; Frelet-Barrand, A. The Arabidopsis ATP-binding cassette protein AtMRP5/AtABCC5 is a high affinity inositol hexakisphosphate transporter involved in guard cell signaling and phytate storage. J. Biol. Chem. 2009, 284, 33614–33622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidinger, F.R.; Mahalakshimi, V.; Rao, G.D.P. Assessment of drought resistance in pearl millet (Pennisetum americanum (L.) Leeke). II estimation of genotype response to stress. Aust. J. Agric. Res. 1987, 38, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Sparvoli, F. (IBBA-CNR, Milan, Italy). Leaves transcriptomic microarray analysis of lpa1 vs wt. 2010. [Google Scholar]
- Avonce, N.; Leyman, B.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Thevelein, J.M.; Iturriaga, G. The Arabidopsis trehalose-6-p synthase AtTPS1 gene is a regulator of glucose, abscisic acid and stress signaling. Plant Physiol. 2004, 136, 3649–3659. [Google Scholar] [CrossRef] [PubMed]
- Paul, M. Trehalose 6-phosphate. Curr. Opin. Plant Biol. 2007, 10, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 1998, 379, 633–646. [Google Scholar] [PubMed]
- Yanhui, C.; Xiaoyuan, Y.; Kun, H.; Meihua, L.; Jigang, L.; Zhaofeng, G. The MYB transcription factor superfamily of Arabidopsis: Expression analysis and phylogenetic comparison with the rice MYB family. Plant Mol. Biol. 2006, 60, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Talbi, C.; Sànchez, C.; Hidalgo-Garcia, A.; Gonzàlez, E.M.; Arrese-Igor, C.; Girard, L. Enhanced expression of Rhizobium etli cbb3 oxidase improves drought tolerance of common bean symbiotic nitrogen fixation. J. Exp. Bot. 2012, 63, 5035–5043. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Tai, T.H. Identification of genes necessary for wild-type levels of seed phytic acid in Arabidopsis thaliana using a reverse genetics approach. Mol. Genet. Genom. 2011, 286, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Rangarajan, P.; Donahue, J.L.; Williams, S.P.; Land, E.S.; Mandal, M.K. Two inositol hexakisphosphate kinases drive inositol pyrophosphate synthesis in plants. Plant J. 2014, 80, 642–653. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Annotation | lpa1 | BAT 93 | Fold Change lpa1/BAT93 (log2) |
|---|---|---|---|---|
| nifA | Transcriptional regulator NifA protein | 0.128 | 0.075 | 0.778 |
| nifH | Nitrogenase, iron protein | 6.485 | 13.661 | −1.075 |
| fixNd | cbb3 oxidase | 0.150 | 0.018 | 3.037 |
| katG | Catalase | 0.437 | 0.200 | 1.126 |
| oxyR | Hydrogen peroxide sensing transcriptional regulator protein | 0.042 | 0.006 | 2.899 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiozzotto, R.; Ramírez, M.; Talbi, C.; Cominelli, E.; Girard, L.; Sparvoli, F.; Hernández, G. Characterization of the Symbiotic Nitrogen-Fixing Common Bean Low Phytic Acid (lpa1) Mutant Response to Water Stress. Genes 2018, 9, 99. https://doi.org/10.3390/genes9020099
Chiozzotto R, Ramírez M, Talbi C, Cominelli E, Girard L, Sparvoli F, Hernández G. Characterization of the Symbiotic Nitrogen-Fixing Common Bean Low Phytic Acid (lpa1) Mutant Response to Water Stress. Genes. 2018; 9(2):99. https://doi.org/10.3390/genes9020099
Chicago/Turabian StyleChiozzotto, Remo, Mario Ramírez, Chouhra Talbi, Eleonora Cominelli, Lourdes Girard, Francesca Sparvoli, and Georgina Hernández. 2018. "Characterization of the Symbiotic Nitrogen-Fixing Common Bean Low Phytic Acid (lpa1) Mutant Response to Water Stress" Genes 9, no. 2: 99. https://doi.org/10.3390/genes9020099
APA StyleChiozzotto, R., Ramírez, M., Talbi, C., Cominelli, E., Girard, L., Sparvoli, F., & Hernández, G. (2018). Characterization of the Symbiotic Nitrogen-Fixing Common Bean Low Phytic Acid (lpa1) Mutant Response to Water Stress. Genes, 9(2), 99. https://doi.org/10.3390/genes9020099