The Unforeseen Non-Coding RNAs in Head and Neck Cancer

 ,
,  ,
, 

Abstract
:1. Introduction
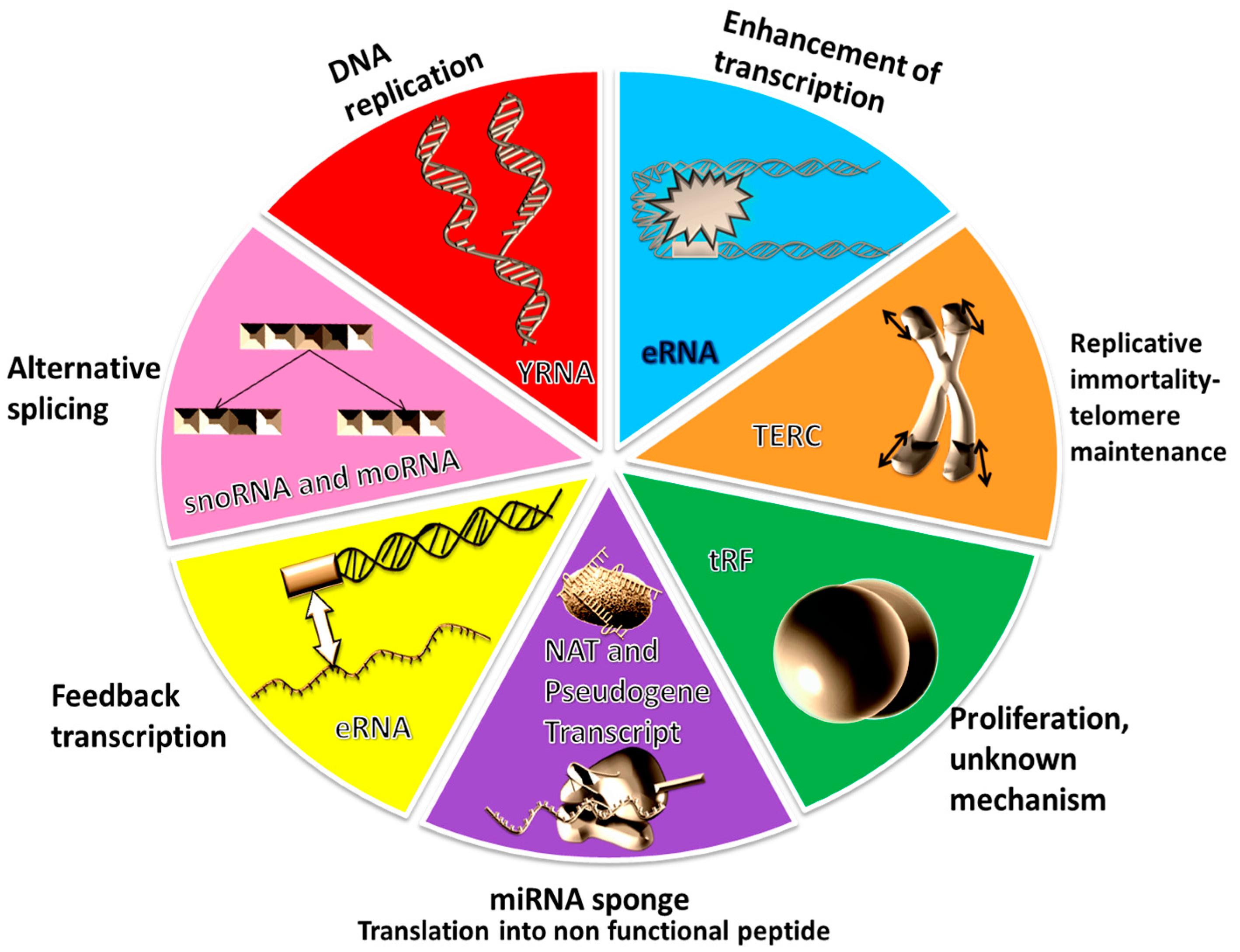
2. The Unforeseen RNAs
3. Enhancer RNAs
4. Stable Intronic Sequence RNAs
5. YRNAs
6. The Human Telomerase RNA Component or Human Telomerase RNA
7. Natural Antisense Transcripts
8. The Small Nucleolar RNAs
9. tRNA Derived Small RNAs
10. Pseudogene Transcripts
11. miRNA-Offset RNAs
12. Ultraconserved Regions of the DNA
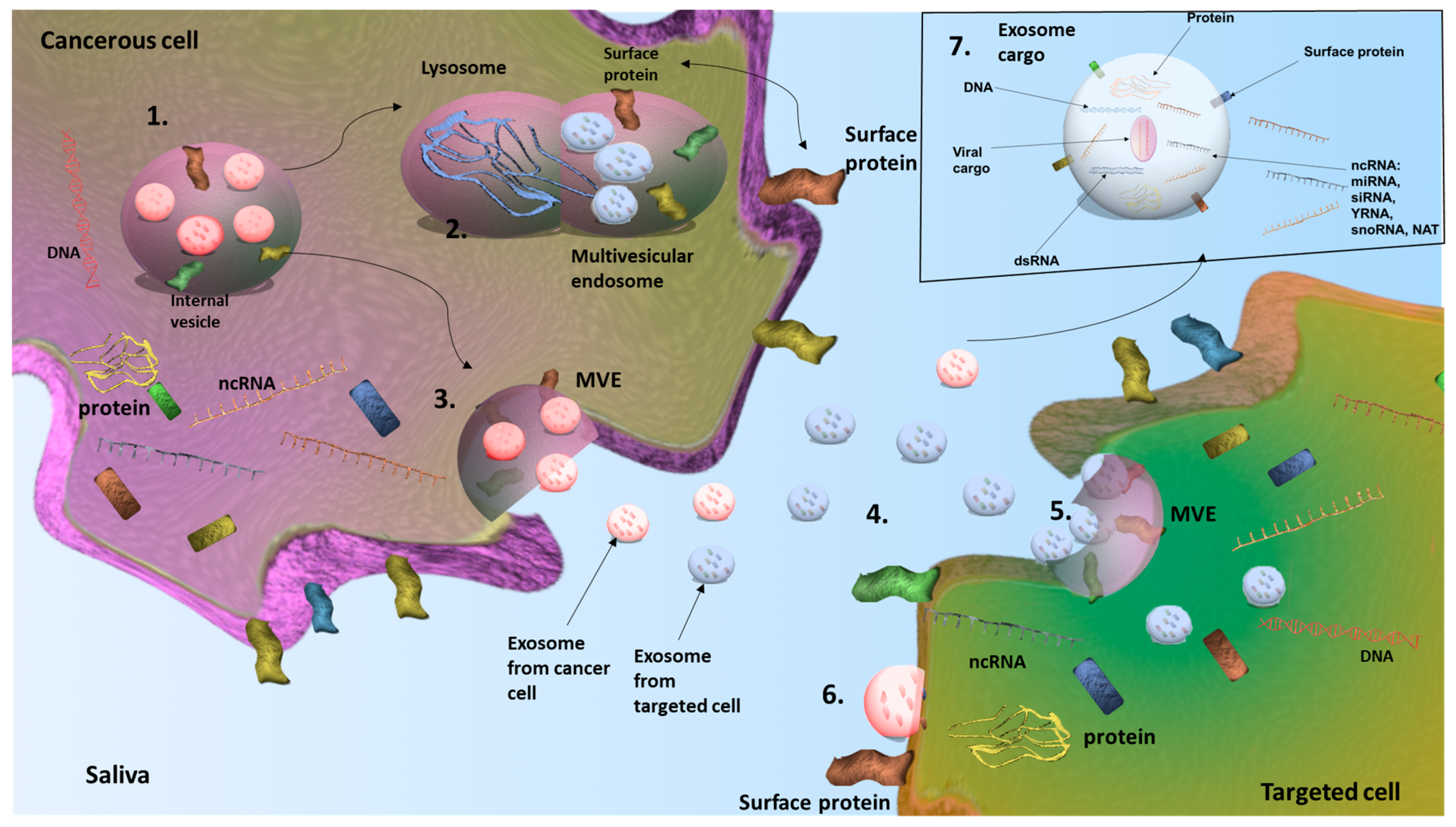
13. Exosomes as Important Sources of Rare ncRNAs
14. Conclusion and Perspective
Conflicts of Interest
References
- Chung, C.H.; Dietz, A.; Gregoire, V.; Guzzo, M.; Hamoir, M.; Leemans, R.; Lefebvre, J.-L.; Licitra, L.; El-Naggar, A.K.; O’Sullivan, B.; et al. Head and Neck Cancer. In Oxford Textbook of Oncology, 3rd ed.; Kerr, D., Haller, D., Velde, C., Baumann, M., Eds.; Oxford Medicine: Oxford, UK, 2016. [Google Scholar]
- Lim, H.; Kim, D.H.; Jung, H.Y.; Gong, E.J.; Na, H.K.; Ahn, J.Y.; Kim, M.Y.; Lee, J.H.; Choi, K.S.; Choi, K.D.; et al. Clinical significance of early detection of esophageal cancer in patients with head and neck cancer. Gut Liver 2015, 9, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Yete, S.; D’Souza, W.; Saranath, D. High-Risk Human Papillomavirus in Oral Cancer: Clinical Implications. Oncology 2018, 94, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Dong, Y. Human papillomavirus and oral squamous cell carcinoma: A review of HPV-positive oral squamous cell carcinoma and possible strategies for future. Curr. Probl. Cancer 2017, 41, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Catana, C.S.; Calin, G.A.; Berindan-Neagoe, I. Inflamma-miRs in Aging and Breast Cancer: Are They Reliable Players? Front. Med. 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.E.; Grandis, J.R.; Ko, A.H. New Strategies in Esophageal Carcinoma: Translational Insights from Signaling Pathways and Immune Checkpoints. Clin. Cancer Res. 2016, 22, 4283–4290. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Cojocneanu-Petric, R.; Berindan-Neagoe, I.; Campian, R.S. Novel technologies for oral squamous carcinoma biomarkers in diagnostics and prognostics. Acta Odontol. Scand. 2015, 73, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Gulei, D.; Magdo, L.; Jurj, A.; Raduly, L.; Cojocneanu-Petric, R.; Moldovan, A.; Moldovan, C.; Florea, A.; Pasca, S.; Pop, L.A.; et al. The silent healer: miR-205–5p up-regulation inhibits epithelial to mesenchymal transition in colon cancer cells by indirectly up-regulating E-cadherin expression. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Monroig Pdel, C.; Pasculli, B.; Calin, G.A. MicroRNAome genome: A treasure for cancer diagnosis and therapy. Cancer J. Clin. 2014, 64, 311–336. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Braicu, C.; Gulei, D.; Tomuleasa, C.; Calin, G.A. Noncoding RNAs in Lung Cancer Angiogenesis. In Physiologic and Pathologic Angiogenesis—Signaling Mechanisms and Targeted Therapy; Simionescu, D., Simionescu, A., Eds.; InTech: Rijeka, Croacia, 2017. [Google Scholar]
- Braicu, C.; Calin, G.A.; Berindan-Neagoe, I. MicroRNAs and cancer therapy— from bystanders to major players. Curr. Med. Chem. 2013, 20, 3561–3573. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Sonea, L.; Zimta, A.A.; Cojocneanu-Petric, R.; Tonchev, K.; Mehterov, N.; Diudea, D.; Buduru, S.; Berindan-Neagoe, I. A Looking-Glass of Non-coding RNAs in oral cancer. Int. J. Mol. Sci. 2017, 18, 2620. [Google Scholar] [CrossRef] [PubMed]
- Pop-Bica, C.; Gulei, D.; Cojocneanu-Petric, R.; Braicu, C.; Petrut, B.; Berindan-Neagoe, I. Understanding the Role of Non-Coding RNAs in Bladder Cancer: From Dark Matter to Valuable Therapeutic Targets. Int. J. Mol. Sci. 2017, 18, 1514. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Revisiting the central dogma in the 21st century. Ann. N. Y. Acad. Sci. 2009, 1178, 6–28. [Google Scholar] [CrossRef] [PubMed]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Catana, C.S.; Pichler, M.; Giannelli, G.; Mader, R.M.; Berindan-Neagoe, I. Non-coding RNAs, the Trojan horse in two-way communication between tumor and stroma in colorectal and hepatocellular carcinoma. Oncotarget 2017, 8, 29519–29534. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Catana, C.; Calin, G.A.; Berindan-Neagoe, I. NCRNA combined therapy as future treatment option for cancer. Curr. Pharm. Des. 2014, 20, 6565–6574. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Sampath, K. cncRNAs: Bi-functional RNAs with protein coding and non-coding functions. Semin. Cell Dev. Biol. 2015, 47–48, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Garzon, R.; Andreeff, M.; Kantarjian, H.M.; Garcia-Manero, G.; Calin, G.A. MicroRNAs and noncoding RNAs in hematological malignancies: Molecular, clinical and therapeutic implications. Leukemia 2008, 22, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Cojocneanu-Petric, R.; Chira, S.; Truta, A.; Floares, A.; Petrut, B.; Achimas-Cadariu, P.; Berindan-Neagoe, I.; et al. Clinical and pathological implications of miRNA in bladder cancer. Int. J. Nanomed. 2015, 10, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Pickard, K.; Ivan, C.; Isella, C.; Ikuo, M.; Mitter, R.; Spizzo, R.; Bullock, M.; Braicu, C.; Pileczki, V.; et al. The clinical and biological significance of MIR-224 expression in colorectal cancer metastasis. Gut 2016, 65, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Calin, G.A. Molecular pathways: microRNAs, cancer cells, and microenvironment. Clin. Cancer. Res. 2014, 20, 6247–6253. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. microRNAs: Tiny regulators with great potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene LIN-4 encodes small RNAs with antisense complementarity to LIN-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Sannigrahi, M.K.; Sharma, R.; Panda, N.K.; Khullar, M. Role of non-coding RNAs in head and neck squamous cell carcinoma: A narrative review. Oral Dis. 2017, 21. [Google Scholar] [CrossRef] [PubMed]
- Pileczki, V.; Braicu, C.; Gherman, C.D.; Berindan-Neagoe, I. TNF-α gene knockout in triple negative breast cancer cell line induces apoptosis. Int. J. Mol. Sci. 2012, 14, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Jurj, A.; Braicu, C.; Pop, L.A.; Tomuleasa, C.; Gherman, C.D.; Berindan-Neagoe, I. The new era of nanotechnology, an alternative to change cancer treatment. Drug Des. Dev. Ther. 2017, 11, 2871–2890. [Google Scholar] [CrossRef] [PubMed]
- Irimie, A.I.; Braicu, C.; Pileczki, V.; Petrushev, B.; Soritau, O.; Campian, R.S.; Berindan-Neagoe, I. Knocking down of p53 triggers apoptosis and autophagy, concomitantly with inhibition of migration on SSC-4 oral squamous carcinoma cells. Mol. Cell. Biochem. 2016, 419, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Trivedi, P.; Jain, N.K. Advances in siRNA delivery in cancer therapy. Artif. Cells Nanomed. Biotechnol. 2018, 46, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Dana, H.; Chalbatani, G.M.; Mahmoodzadeh, H.; Karimloo, R.; Rezaiean, O.; Moradzadeh, A.; Mehmandoost, N.; Moazzen, F.; Mazraeh, A.; Marmari, V. Molecular Mechanisms and Biological Functions of siRNA. Int. J. Biomed. Sci. 2017, 13, 48–57. [Google Scholar] [PubMed]
- Irimie, A.I.; Sonea, L.; Jurj, A.; Mehterov, N.; Zimta, A.A.; Budisan, L.; Braicu, C.; Berindan-Neagoe, I. Future trends and emerging issues for nanodelivery systems in oral and oropharyngeal cancer. Int. J. Nanomed。 2017, 12, 4593–4606. [Google Scholar] [CrossRef] [PubMed]
- Berindan-Neagoe, I.; Braicu, C.; Irimie, A. Combining the chemotherapeutic effects of epigallocatechin 3-gallate with siRNA-mediated p53 knock-down results in synergic pro-apoptotic effects. Int. J. Nanomed. 2012, 7, 6035–6047. [Google Scholar]
- Lecaros, R.L.; Huang, L.; Lee, T.C.; Hsu, Y.C. Nanoparticle Delivered VEGF-A siRNA Enhances Photodynamic Therapy for Head and Neck Cancer Treatment. Mol. Ther. 2016, 24, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Lecaros, R.L.; Tseng, Y.C.; Huang, L.; Hsu, Y.C. Nanoparticle delivery of HIF1α siRNA combined with photodynamic therapy as a potential treatment strategy for head-and-neck cancer. Cancer Lett. 2015, 359, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Taft, R.J.; Simons, C.; Nahkuri, S.; Oey, H.; Korbie, D.J.; Mercer, T.R.; Holst, J.; Ritchie, W.; Wong, J.J.; Rasko, J.E.; et al. Nuclear-localized tiny RNAs are associated with transcription initiation and splice sites in metazoans. Nat. Struct. Mol. Biol. 2010, 17, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Umbach, J.L.; Strelow, L.I.; Wong, S.W.; Cullen, B.R. Analysis of rhesus rhadinovirus microRNAs expressed in virus-induced tumors from infected rhesus macaques. Virology 2010, 405, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Schnitzler, G.R.; Iyer, L.K.; Aronovitz, M.J.; Baur, W.E.; Karas, R.H. MicroRNA-Offset RNA Alters Gene Expression and Cell Proliferation. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, J.; Tournier, J.M.; Puchelle, E. In vitro evidence that human airway lysozyme is cleaved and inactivated by Pseudomonas aeruginosa elastase and not by human leukocyte elastase. Infect. Immun. 1985, 47, 555–560. [Google Scholar] [PubMed]
- Lujambio, A.; Portela, A.; Liz, J.; Melo, S.A.; Rossi, S.; Spizzo, R.; Croce, C.M.; Calin, G.A.; Esteller, M. CpG island hypermethylation-associated silencing of non-coding RNAs transcribed from ultraconserved regions in human cancer. Oncogene 2010, 29, 6390–6401. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Liu, C.G.; Ferracin, M.; Hyslop, T.; Spizzo, R.; Sevignani, C.; Fabbri, M.; Cimmino, A.; Lee, E.J.; Wojcik, S.E.; et al. Ultraconserved regions encoding ncRNAs are altered in human leukemias and carcinomas. Cancer Cell 2007, 12, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Fabris, L.; Calin, G.A. Understanding the Genomic Ultraconservations: T-UCRs and Cancer. Int. Rev. Cell. Mol. Biol. 2017, 333, 159–172. [Google Scholar] [PubMed]
- Terreri, S.; Durso, M.; Colonna, V.; Romanelli, A.; Terracciano, D.; Ferro, M.; Perdona, S.; Castaldo, L.; Febbraio, F.; de Nigris, F.; et al. New Cross-Talk Layer between Ultraconserved Non-Coding RNAs, MicroRNAs and Polycomb Protein YY1 in Bladder Cancer. Genes 2016, 7, 127. [Google Scholar] [CrossRef] [PubMed]
- Pek, J.W.; Osman, I.; Tay, M.L.; Zheng, R.T. Stable intronic sequence RNAs have possible regulatory roles in Drosophila melanogaster. J. Cell Biol. 2015, 211, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Osman, I.; Tay, M.L.; Pek, J.W. Stable intronic sequence RNAs (sisRNAs): A new layer of gene regulation. Cell. Mol. Life Sci. 2016, 73, 3507–3519. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.F.; Yang, L.; Zhang, Y.; Xiang, J.F.; Wu, Y.W.; Carmichael, G.G.; Chen, L.L. Long noncoding RNAs with snoRNA ends. Mol. Cell. 2012, 48, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Yamada, K.; Avolio, F.; Scott, M.S.; van Koningsbruggen, S.; Barton, G.J.; Lamond, A.I. Analysis of human small nucleolar RNAs (snoRNA) and the development of snoRNA modulator of gene expression vectors. Mol. Biol. Cell 2010, 21, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.J.; Fuchs, G.; Fu, C.; Wolin, S.L.; Reinisch, K.M. Structural insights into RNA quality control: The Ro autoantigen binds misfolded RNAs via its central cavity. Cell 2005, 121, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Christov, C.P.; Gardiner, T.J.; Szuts, D.; Krude, T. Functional requirement of noncoding Y RNAs for human chromosomal DNA replication. Mol. Cell. Biol. 2006, 26, 6993–7004. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Zorca, C.E.; Traboulsi, T.; Noutahi, E.; Krause, M.R.; Mader, S.; Zenklusen, D. Single-cell profiling reveals that eRNA accumulation at enhancer-promoter loops is not required to sustain transcription. Nucleic Acids Res. 2017, 45, 3017–3030. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.K.; Wang, H.F.; Burns, A.M.; Schroeder, M.R.; Gaspari, M.; Baumann, P. Human Telomerase RNA Processing and Quality Control. Cell Rep. 2015, 13, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Choudury, S.G.; Slotkin, R.K. tRNA-derived small RNAs target transposable element transcripts. Nucleic Acids Res. 2017, 45, 5142–5152. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [PubMed]
- Werner, A.; Swan, D. What are natural antisense transcripts good for? Biochem. Soc. Trans. 2010, 38, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Joska, T.M.; Ruesch, C.E.; Coster, S.J.; Belden, W.J. The frequency natural antisense transcript first promotes, then represses, frequency gene expression via facultative heterochromatin. Proc. Natl. Acad. Sci. USA 2015, 112, 4357–4362. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.S.; Li, S.; Kho, J.R.; Huang, K.; Zhang, Y. Network analysis of pseudogene-gene relationships: From pseudogene evolution to their functional potentials. Pac. Symp. Biocomput. 2018, 23, 536–547. [Google Scholar] [PubMed]
- Ji, Z.; Song, R.; Regev, A.; Struhl, K. Many lncRNAs, 5′UTRs, and pseudogenes are translated and some are likely to express functional proteins. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, J. Are Human Translated Pseudogenes Functional? Mol. Biol. Evol. 2016, 33, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Marranci, A.; Pandolfi, P.P. Pseudogenes in Human Cancer. Front. Med. 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Kwok, Z.H.; Chew, X.H.; Zhang, B.; Liu, C.; Soong, T.W.; Yang, H.; Tay, Y. A FTH1 gene: Pseudogene:microRNA network regulates tumorigenesis in prostate cancer. Nucleic Acids Res. 2017, gkx1248. [Google Scholar] [CrossRef]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genom. Proteom. Bioinform. 2016, 14, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing miRNA-lncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [PubMed]
- Jung, H.; Lee, D.; Lee, J.; Park, D.; Kim, Y.J.; Park, W.Y.; Hong, D.; Park, P.J.; Lee, E. Intron retention is a widespread mechanism of tumor-suppressor inactivation. Nat. Genet. 2015, 47, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Shlyueva, D.; Stampfel, G.; Stark, A. Transcriptional enhancers: From properties to genome-wide predictions. Nat. Rev. Genet. 2014, 15, 272–286. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Shiekhattar, R. Enhancer RNAs: The new molecules of transcription. Curr. Opin. Genet. Dev. 2014, 25, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Melamed, P.; Yosefzon, Y.; Rudnizky, S.; Pnueli, L. Transcriptional enhancers: Transcription, function and flexibility. Transcription 2016, 7, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Vuong, B.Q.; Vaidyanathan, B.; Lin, J.Y.; Huang, F.T.; Chaudhuri, J. Non-coding RNA Generated following Lariat Debranching Mediates Targeting of AID to DNA. Cell 2015, 161, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, M.P.; Krude, T. Functional roles of non-coding Y RNAs. Int. J. Biochem. Cell. Biol. 2015, 66, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Dhahbi, J.M.; Spindler, S.R.; Atamna, H.; Boffelli, D.; Mote, P.; Martin, D.I. 5′-YRNA fragments derived by processing of transcripts from specific YRNA genes and pseudogenes are abundant in human serum and plasma. Physiol. Genom. 2013, 45, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Hizir, Z.; Bottini, S.; Grandjean, V.; Trabucchi, M.; Repetto, E. RNY (YRNA)-derived small RNAs regulate cell death and inflammation in monocytes/macrophages. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Rubtsova, M.P.; Vasilkova, D.P.; Naraykina, Y.V.; Dontsova, O.A. Peculiarities of Yeasts and Human Telomerase RNAs Processing. Acta Nat. 2016, 8, 14–22. [Google Scholar] [PubMed]
- Rao, X.; Huang, D.; Sui, X.; Liu, G.; Song, X.; Xie, J.; Huang, D. Overexpression of WRAP53 is associated with development and progression of esophageal squamous cell carcinoma. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.S.; Ono, M. From snoRNA to miRNA: Dual function regulatory non-coding RNAs. Biochimie 2011, 93, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Michel, C.I.; Holley, C.L.; Scruggs, B.S.; Sidhu, R.; Brookheart, R.T.; Listenberger, L.L.; Behlke, M.A.; Ory, D.S.; Schaffer, J.E. Small nucleolar RNAs U32a, U33, and U35a are critical mediators of metabolic stress. Cell Metab. 2011, 14, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xing, Y.; Xu, L.; Chen, W.; Cao, W.; Zhang, C. Decreased expression of pseudogene PTENP1 promotes malignant behaviours and is associated with the poor survival of patients with HNSCC. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Asikainen, S.; Heikkinen, L.; Juhila, J.; Holm, F.; Weltner, J.; Trokovic, R.; Mikkola, M.; Toivonen, S.; Balboa, D.; Lampela, R.; et al. Selective microRNA-Offset RNA expression in human embryonic stem cells. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, D.; Terreri, S.; de Nigris, F.; Costa, V.; Calin, G.A.; Cimmino, A. The role of a new class of long noncoding RNAs transcribed from ultraconserved regions in cancer. Biochim. Biophys. Acta 2017, 1868, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Hemberg, M.; Gray, J.M.; Costa, A.M.; Bear, D.M.; Wu, J.; Harmin, D.A.; Laptewicz, M.; Barbara-Haley, K.; Kuersten, S.; et al. Widespread transcription at neuronal activity-regulated enhancers. Nature 2010, 465, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.T.; Li, W.; Rosenfeld, M.G.; Glass, C.K. Enhancer RNAs and regulated transcriptional programs. Trends Biochem. Sci. 2014, 39, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, F.; Ouyang, Z.; An, G.; Zhao, C.; Shuai, J.; Cai, S.; Bo, X.; Shu, W. Functional annotation of structural ncRNAs within enhancer RNAs in the human genome: Implications for human disease. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Hemberg, M.; Gray, J.M. Enhancer RNAs: A class of long noncoding RNAs synthesized at enhancers. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Gardini, A.; Zhang, A.; Shiekhattar, R. Integrator mediates the biogenesis of enhancer RNAs. Nature 2015, 525, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Notani, D.; Rosenfeld, M.G. Enhancers as non-coding RNA transcription units: Recent insights and future perspectives. Nat. Rev. Genet. 2016, 17, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Bal, E.; Park, H.S.; Belaid-Choucair, Z.; Kayserili, H.; Naville, M.; Madrange, M.; Chiticariu, E.; Hadj-Rabia, S.; Cagnard, N.; Kuonen, F.; et al. Mutations in ACTRT1 and its enhancer RNA elements lead to aberrant activation of Hedgehog signaling in inherited and sporadic basal cell carcinomas. Nat. Med. 2017, 23, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.E.; Golan, T.; Sheinboim, D.; Malcov, H.; Amar, D.; Salamon, A.; Liron, T.; Gelfman, S.; Gabet, Y.; Shamir, R.; et al. Enhancer methylation dynamics contribute to cancer plasticity and patient mortality. Genome Res. 2016, 26, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Younger, S.T.; Kenzelmann-Broz, D.; Jung, H.; Attardi, L.D.; Rinn, J.L. Integrative genomic analysis reveals widespread enhancer regulation by p53 in response to DNA damage. Nucleic Acids Res. 2015, 43, 4447–4462. [Google Scholar] [CrossRef] [PubMed]
- Rowley, H.; Sherrington, P.; Helliwell, T.R.; Kinsella, A.; Jones, A.S. p53 expression and p53 gene mutation in oral cancer and dysplasia. Otolaryngol. Head Neck Surg. 1998, 118, 115–123. [Google Scholar] [CrossRef]
- Gardner, E.J.; Nizami, Z.F.; Talbot, C.C., Jr.; Gall, J.G. Stable intronic sequence RNA (sisRNA), a new class of noncoding RNA from the oocyte nucleus of Xenopus tropicalis. Genes Dev. 2012, 26, 2550–2559. [Google Scholar] [CrossRef] [PubMed]
- Shimakage, M.; Horii, K.; Tempaku, A.; Kakudo, K.; Shirasaka, T.; Sasagawa, T. Association of Epstein-Barr virus with oral cancers. Hum. Pathol. 2002, 33, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Moss, W.N.; Steitz, J.A. Genome-wide analyses of Epstein-Barr virus reveal conserved RNA structures and a novel stable intronic sequence RNA. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Farris, A.D.; O’Brien, C.A.; Harley, J.B. Y3 is the most conserved small RNA component of Ro ribonucleoprotein complexes in vertebrate species. Gene 1995, 154, 193–198. [Google Scholar] [CrossRef]
- Kohn, M.; Pazaitis, N.; Huttelmaier, S. Why YRNAs? About Versatile RNAs and Their Functions. Biomolecules 2013, 3, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Mosig, A.; Guofeng, M.; Stadler, B.M.; Stadler, P.F. Evolution of the vertebrate Y RNA cluster. Theory Biosci. 2007, 126, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.E.; Turnbull, C.; Dalmay, T. Y RNAs: Recent developments. Biomol. Concepts 2013, 4, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Perreault, J.; Perreault, J.P.; Boire, G. Ro-associated Y RNAs in metazoans: Evolution and diversification. Mol. Biol. Evol. 2007, 24, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Tolkach, Y.; Stahl, A.F.; Niehoff, E.-M.; Zhao, C.; Kristiansen, G.; Müller, S.C.; Ellinger, J. YRNA expression predicts survival in bladder cancer patients. BMC Cancer 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- Hendrick, J.P.; Wolin, S.L.; Rinke, J.; Lerner, M.R.; Steitz, J.A. Ro small cytoplasmic ribonucleoproteins are a subclass of La ribonucleoproteins: Further characterization of the Ro and La small ribonucleoproteins from uninfected mammalian cells. Mol. Cell. Biol. 1981, 1, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Dhahbi, J.M.; Spindler, S.R.; Atamna, H.; Boffelli, D.; Martin, D.I.K. Deep Sequencing of Serum Small RNAs Identifies Patterns of 5′ tRNA Half and YRNA Fragment Expression Associated with Breast Cancer. Biomark. Cancer 2014, 6, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Victoria Martinez, B.; Dhahbi, J.M.; Nunez Lopez, Y.O.; Lamperska, K.; Golusinski, P.; Luczewski, L.; Kolenda, T.; Atamna, H.; Spindler, S.R.; Golusinski, W.; et al. Circulating small non-coding RNA signature in head and neck squamous cell carcinoma. Oncotarget 2015, 6, 19246–19263. [Google Scholar] [PubMed]
- Repetto, E.; Lichtenstein, L.; Hizir, Z.; Tekaya, N.; Benahmed, M.; Ruidavets, J.-B.; Zaragosi, L.-E.; Perret, B.; Bouchareychas, L.; Genoux, A.; et al. RNY-derived small RNAs as a signature of coronary artery disease. BMC Med. 2015, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baena-Del Valle, J.A.; Zheng, Q.; Esopi, D.M.; Rubenstein, M.; Hubbard, G.K.; Moncaliano, M.C.; Hruszkewycz, A.; Vaghasia, A.; Yegnasubramanian, S.; Wheelan, S.J.; et al. MYC drives overexpression of telomerase RNA (hTR/TERC) in prostate cancer. J. Pathol. 2018, 244, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Funk, W.D.; Wang, S.S.; Weinrich, S.L.; Avilion, A.A.; Chiu, C.P.; Adams, R.R.; Chang, E.; Allsopp, R.C.; Yu, J.; et al. The RNA component of human telomerase. Science 1995, 269, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Kilchert, C.; Wittmann, S.; Vasiljeva, L. The regulation and functions of the nuclear RNA exosome complex. Nat. Rev. Mol. Cell Biol. 2016, 17, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Schmidt, J.C.; Goldfarb, K.C.; Cech, T.R.; Parker, R. Inhibition of telomerase RNA decay rescues telomerase deficiency caused by dyskerin or PARN defects. Nat. Struct. Mol. Biol. 2016, 23, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Christensen, R.; Park, N.H.; Sapp, P.; Kang, M.K.; Park, N.H. Elevated expression of hTERT is associated with dysplastic cell transformation during human oral carcinogenesis in situ. Clin. Cancer Res. 2001, 7, 3079–3086. [Google Scholar] [PubMed]
- Majumder, M.; House, R.; Palanisamy, N.; Qie, S.; Day, T.A.; Neskey, D.; Diehl, J.A.; Palanisamy, V. RNA-Binding Protein FXR1 Regulates p21 and TERC RNA to Bypass p53-Mediated Cellular Senescence in OSCC. PLoS Genet. 2016, 12. [Google Scholar] [CrossRef]
- Fernandez, E.; Mallette, F.A. The Rise of FXR1: Escaping Cellular Senescence in Head and Neck Squamous Cell Carcinoma. PLoS Genet. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Lacatena, R.M.; Cesareni, G. Base pairing of RNA I with its complementary sequence in the primer precursor inhibits ColE1 replication. Nature 1981, 294, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Rosikiewicz, W.; Makalowska, I. Biological functions of natural antisense transcripts. Acta Biochim. Pol. 2016, 63, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Coucoravas, C.; Dhanjal, S.; Henriksson, S.; Bohm, S.; Farnebo, M. Phosphorylation of the Cajal body protein WRAP53beta by ATM promotes its involvement in the DNA damage response. RNA Biol. 2017, 14, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.; Sletten, A.C.; Lee, J.; Pyles, K.D.; Matkovich, S.J.; Ory, D.S.; Schaffer, J.E. Nuclear export factor 3 regulates localization of small nucleolar RNAs. J. Biol. Chem. 2017, 292, 20228–20239. [Google Scholar] [CrossRef] [PubMed]
- Holley, C.L.; Li, M.W.; Scruggs, B.S.; Matkovich, S.J.; Ory, D.S.; Schaffer, J.E. Cytosolic accumulation of small nucleolar RNAs (snoRNAs) is dynamically regulated by NADPH oxidase. J. Biol. Chem. 2015, 290, 11741–11748. [Google Scholar] [CrossRef] [PubMed]
- Falaleeva, M.; Stamm, S. Processing of snoRNAs as a new source of regulatory non-coding RNAs: snoRNA fragments form a new class of functional RNAs. Bioessays 2013, 35, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.C.; Brown, D.D.; Brown, J.W. The small nucleolar ribonucleoprotein (snoRNP) database. RNA 2010, 16, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Kishore, S.; Gruber, A.R.; Jedlinski, D.J.; Syed, A.P.; Jorjani, H.; Zavolan, M. Insights into snoRNA biogenesis and processing from PAR-CLIP of snoRNA core proteins and small RNA sequencing. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.E.; Ku, J.; Honda, T.K.; Yu, V.; Kuo, S.Z.; Zheng, H.; Xuan, Y.; Saad, M.A.; Hinton, A.; Brumund, K.T.; et al. Transcriptome sequencing uncovers novel long noncoding and small nucleolar RNAs dysregulated in head and neck squamous cell carcinoma. RNA 2015, 21, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- Davanian, H.; Balasiddaiah, A.; Heymann, R.; Sundstrom, M.; Redenstrom, P.; Silfverberg, M.; Brodin, D.; Sallberg, M.; Lindskog, S.; Kruger Weiner, C.; et al. Ameloblastoma RNA profiling uncovers a distinct non-coding RNA signature. Oncotarget 2017, 8, 4530–4542. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Petronacci, C.; Perez-Sayans, M.; Padin-Iruegas, M.E.; Marichalar-Mendia, X.; Gallas-Torreira, M.; Garcia Garcia, A. Differential expression of snoRNAs in oral squamous cell carcinomas: New potential diagnostic markers. J. Enzyme Inhib. Med. Chem. 2018, 33, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Kunzmann, A.; Brennicke, A.; Marchfelder, A. 5′ end maturation and RNA editing have to precede tRNA 3′ processing in plant mitochondria. Proc. Natl. Acad. Sci. USA 1998, 95, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Betat, H.; Long, Y.; Jackman, J.E.; Mörl, M. From End to End: tRNA Editing at 5′- and 3′-Terminal Positions. Int. J. Mol. Sci. 2014, 15, 23975–23998. [Google Scholar] [CrossRef] [PubMed]
- Pederson, T. Regulatory RNAs derived from transfer RNA? RNA 2010, 16, 1865–1869. [Google Scholar] [CrossRef] [PubMed]
- Diebel, K.W.; Zhou, K.; Clarke, A.B.; Bemis, L.T. Beyond the Ribosome: Extra-translational Functions of tRNA Fragments. Biomark. Insights 2016, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Lin, M.; Rockowitz, S.; Lachman, H.M.; Zheng, D. Characterization of human pseudogene-derived non-coding RNAs for functional potential. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Spizzo, R.; Almeida, M.I.; Colombatti, A.; Calin, G.A. Long non-coding RNAs and cancer: A new frontier of translational research? Oncogene 2012, 31, 4577–4587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tutar, Y. Pseudogenes. Comp. Funct. Genom. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Perreault, J.; Noel, J.F.; Briere, F.; Cousineau, B.; Lucier, J.F.; Perreault, J.P.; Boire, G. Retropseudogenes derived from the human Ro/SS-A autoantigen-associated hY RNAs. Nucleic Acids Res. 2005, 33, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Hendrix, D.; Levine, M.; Haley, B. A distinct class of small RNAs arises from pre-miRNA-proximal regions in a simple chordate. Nat. Struct. Mol. Biol. 2009, 16, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, S.; Biasiolo, M.; Bisognin, A. MicroRNA-offset RNAs (moRNAs): By-product spectators or functional players? Trends Mol. Med. 2011, 17, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Fassan, M.; Dall’Olmo, L.; Galasso, M.; Braconi, C.; Pizzi, M.; Realdon, S.; Volinia, S.; Valeri, N.; Gasparini, P.; Baffa, R.; et al. Transcribed ultraconserved noncoding RNAs (T-UCR) are involved in Barrett’s esophagus carcinogenesis. Oncotarget 2014, 5, 7162–7171. [Google Scholar] [CrossRef] [PubMed]
- Boscolo-Rizzo, P.; Da Mosto, M.C.; Rampazzo, E.; Giunco, S.; Del Mistro, A.; Menegaldo, A.; Baboci, L.; Mantovani, M.; Tirelli, G.; De Rossi, A. Telomeres and telomerase in head and neck squamous cell carcinoma: From pathogenesis to clinical implications. Cancer Metastasis Rev. 2016, 35, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Gulei, D.; Irimie, A.I.; Cojocneanu-Petric, R.; Schultze, J.L.; Berindan-Neagoe, I. Exosomes-Small Players, Big Sound. Bioconjug. Chem. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, Y.; Duan, J.; Ma, Y.; Shen, Z.; Wei, L.; Cui, X.; Zhang, J.; Xie, Y.; Liu, J. Quantitative proteomic analysis of exosome protein content changes induced by hepatitis B virus in Huh-7 cells using SILAC labeling and LC-MS/MS. J. Proteome Res. 2014, 13, 5391–5402. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Odenthal, M.; Fries, J.W.U. Exosomes as miRNA Carriers: Formation–Function–Future. Int. J. Mol. Sci. 2016, 17, 2028. [Google Scholar] [CrossRef] [PubMed]
- Shahabipour, F.; Banach, M.; Sahebkar, A. Exosomes as nanocarriers for siRNA delivery: Paradigms and challenges. Arch. Med. Sci. 2016, 12, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Van Balkom, B.W.; Eisele, A.S.; Pegtel, D.M.; Bervoets, S.; Verhaar, M.C. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J. Extracell. Vesicles 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortty, S.K.; Prakash, A.; Nechooshtan, G.; Hearn, S.; Gingeras, T.R. Extracellular vesicle-mediated transfer of processed and functional RNY5 RNA. RNA 2015, 21, 1966–1979. [Google Scholar] [CrossRef] [PubMed]
- Tosar, J.P.; Gambaro, F.; Sanguinetti, J.; Bonilla, B.; Witwer, K.W.; Cayota, A. Assessment of small RNA sorting into different extracellular fractions revealed by high-throughput sequencing of breast cell lines. Nucleic Acids Res. 2015, 43, 5601–5616. [Google Scholar] [CrossRef] [PubMed]
- Chiba, M.; Kimura, M.; Asari, S. Exosomes secreted from human colorectal cancer cell lines contain mRNAs, microRNAs and natural antisense RNAs, that can transfer into the human hepatoma HepG2 and lung cancer A549 cell lines. Oncol. Rep. 2012, 28, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Zlotogorski-Hurvitz, A.; Dayan, D.; Chaushu, G.; Korvala, J.; Salo, T.; Sormunen, R.; Vered, M. Human Saliva-Derived Exosomes: Comparing Methods of Isolation. J. Histochem. Cytochem. 2015, 63, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Principe, S.; Hui, A.B.; Bruce, J.; Sinha, A.; Liu, F.F.; Kislinger, T. Tumor-derived exosomes and microvesicles in head and neck cancer: Implications for tumor biology and biomarker discovery. Proteomics 2013, 13, 1608–1623. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
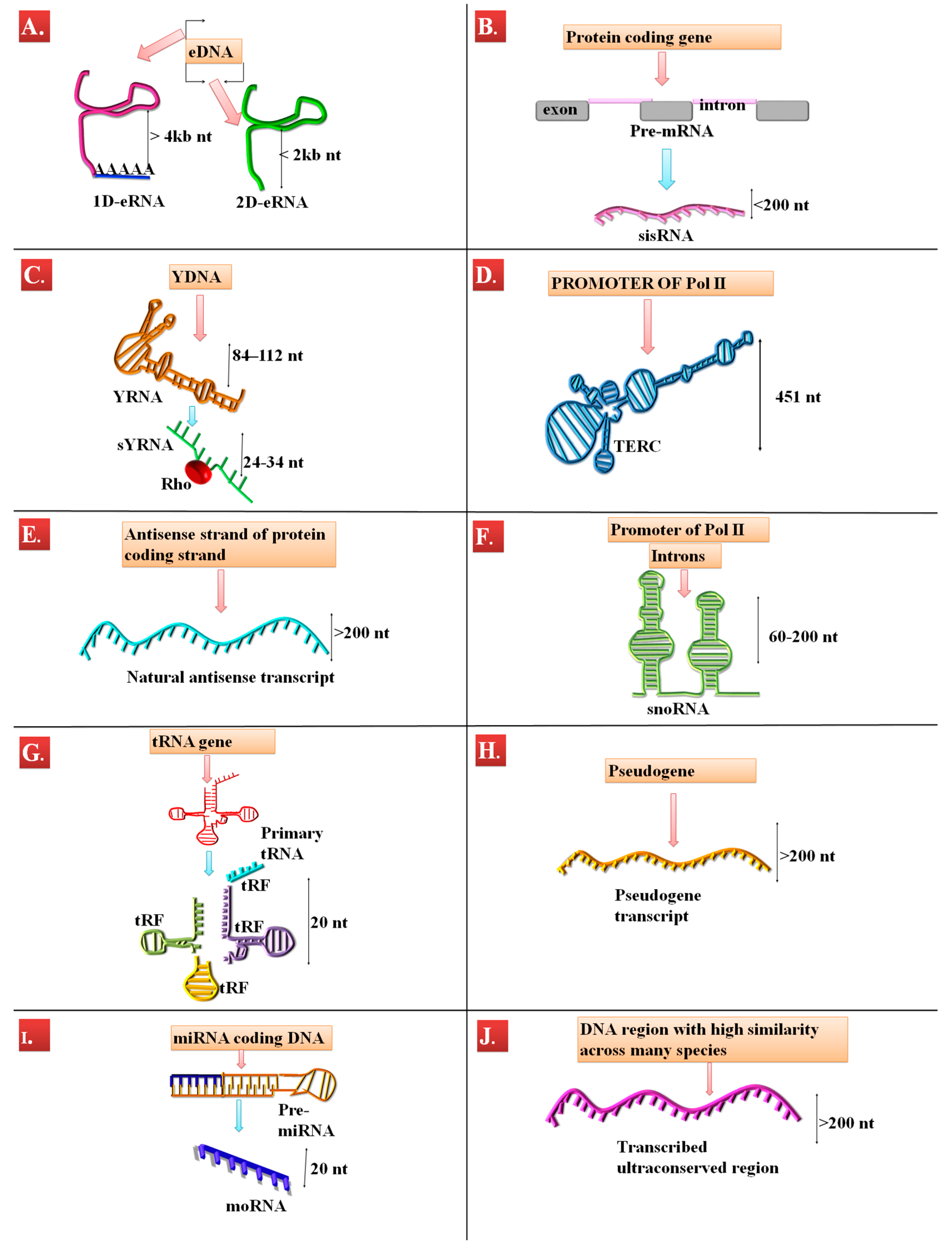
| ncRNA Type | Length (nt) | DNA Region | Mechanism of Action | Reference |
|---|---|---|---|---|
| eRNA | 800–2000 | enhancer DNA | histone methylation | [67,68,69] |
| sisRNA | intronic length | protein-coding genes | alternative splicing, transcription stimulation, antibody isotype class switch | [70,71] |
| YRNA | 84–112 | yDNA | misfolded RNA degradation, DNA replication | [50,72,73] |
| sYRNA | ~24 to ~34 | yDNA | interaction with TLR | [74] |
| TERC/hTR | 451 | Pol II promotor region | telomere maintenance | [54,75] |
| NAT | depending of the coding gene length | antisense strand of protein-coding genes | mRNA inhibition | [76] |
| snoRNA | 60-200 C/D snoRNAs, 120–250 for H/ACA snoRNAs | introns, promoter region of Pol II | rRNA processing, splicing | [77] |
| tRF | 20 | tRNA coding genes | Unknown | [78] |
| Pseudogenestranscrips | gene lenght | Pseudogenes | translation repression, miRNA sponge, multiple miRNAs, siRNA origin | [62,79] |
| moRNAs | 20 | miRNA-coding genes | Unknown | [80] |
| T-UCR | >200 | Ultra-conserved regions of the DNA | miRNA sponging | [81] |
| Type of ncRNA | Name/Code | Expression | Determination method | Clinical/In Vitro/In Vivo | Observation | Reference |
|---|---|---|---|---|---|---|
| syRNAs | RNY1, RNY4P17 | Up | Deep sequencing with IlluminaHiSeq 2000 | Clinical blood samples | Altered expression level in head and neck cancer | [104] |
| RNY4P1, RNY4P5 RNY4P8, RNY4P11 Y_RNA.725, Y_RNA.122 RNY4P20, RNY4P25 RNY4P27, Y_RNA.182 Y_RNA.257, RNY4P18 Y_RNA.44, RNY4P6 Y_RNA.668, Y_RNA.7 Y_RNA.796, Y_RNA.292 Y_RNA.662 | Down | |||||
| TERC/hTR | Up | qRT-PCR | in vitro UMSCC74A, UMSCC74B/TCGA | cell senescence inhibited | [111] | |
| Up | Gene copy number—FISH | OSCC—tissue samples vs. normal tissue | installment of oral lesions | [135] | ||
| Up | RT-PCR | oral cancer tissue vs. normal tissue | associated with higher grade dysplasia or carcinoma | [110] | ||
| snoRNAs | SNORD35B | Down | RNA-Seq for lncRNA, (TCGA)/RT-PCR | in vitro normal cell lines OKF4 and OKF6 vs. oral cancer cell lines UMSCC-10B, UMSCC-22B, HN-1, HN-12, and HN-30 | associated with poor prognostic | [121] |
| mediate the pattern of 28S rRNA | ||||||
| 4qI-4,14qII-22,ACA17,U84, mgh18S-121, U18A, U8, 14qII-12, U28 ENSG00000263442, ENSG00000264591, ENSG00000265325, ENSG00000265607, ENSG00000266646, ENSG00000266755, | Down | Microarray | OSCC tissue vs. keratinized gums preserved at −80 °C | Potential biomarkers, prognostic value | [123] | |
| LINC340, SNORD116-25, SNORA11, SNORA21, SNORA47 and SNORA65 | Up | microarray, RT-PCR | Formalin fixed and paraffin embedded (FFPE) samples of ameloblastoma clinical | better diagnostic tool through biomarkers and potential therapeutic targets | [122] | |
| NATs | WRAP53 | Up | qRT-PCR, Western-blot | ESCC cell lines EC109, EC9706, KYSE150, and KYSE180 and tissue from 134 oesophageal cancer patients | higher expression of this transcript is progression of oesophageal cancer | [76] |
| Pseudogenes transcript | PTENP1 | Down | HN4, HN6, HN13Cal27, HN30 cell lines | proliferation, invasion and migration capabilitiesand decreased survival rate | [37] | |
| T-UCRs | uc.214+, uc.328+, uc.329+, uc.356+, | Down | Microarray, RT-PCR, immunohistochemistry | oesophageal cancer | cancerous transformation | [134] |
| uc.202-, uc.223-uc.269- | Up | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irimie, A.I.; Zimta, A.-A.; Ciocan, C.; Mehterov, N.; Dudea, D.; Braicu, C.; Berindan-Neagoe, I. The Unforeseen Non-Coding RNAs in Head and Neck Cancer. Genes 2018, 9, 134. https://doi.org/10.3390/genes9030134
Irimie AI, Zimta A-A, Ciocan C, Mehterov N, Dudea D, Braicu C, Berindan-Neagoe I. The Unforeseen Non-Coding RNAs in Head and Neck Cancer. Genes. 2018; 9(3):134. https://doi.org/10.3390/genes9030134
Chicago/Turabian StyleIrimie, Alexandra Iulia, Alina-Andreea Zimta, Cristina Ciocan, Nikolay Mehterov, Diana Dudea, Cornelia Braicu, and Ioana Berindan-Neagoe. 2018. "The Unforeseen Non-Coding RNAs in Head and Neck Cancer" Genes 9, no. 3: 134. https://doi.org/10.3390/genes9030134
APA StyleIrimie, A. I., Zimta, A. -A., Ciocan, C., Mehterov, N., Dudea, D., Braicu, C., & Berindan-Neagoe, I. (2018). The Unforeseen Non-Coding RNAs in Head and Neck Cancer. Genes, 9(3), 134. https://doi.org/10.3390/genes9030134