Genomic Differentiation during Speciation-with-Gene-Flow: Comparing Geographic and Host-Related Variation in Divergent Life History Adaptation in Rhagoletis pomonella

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
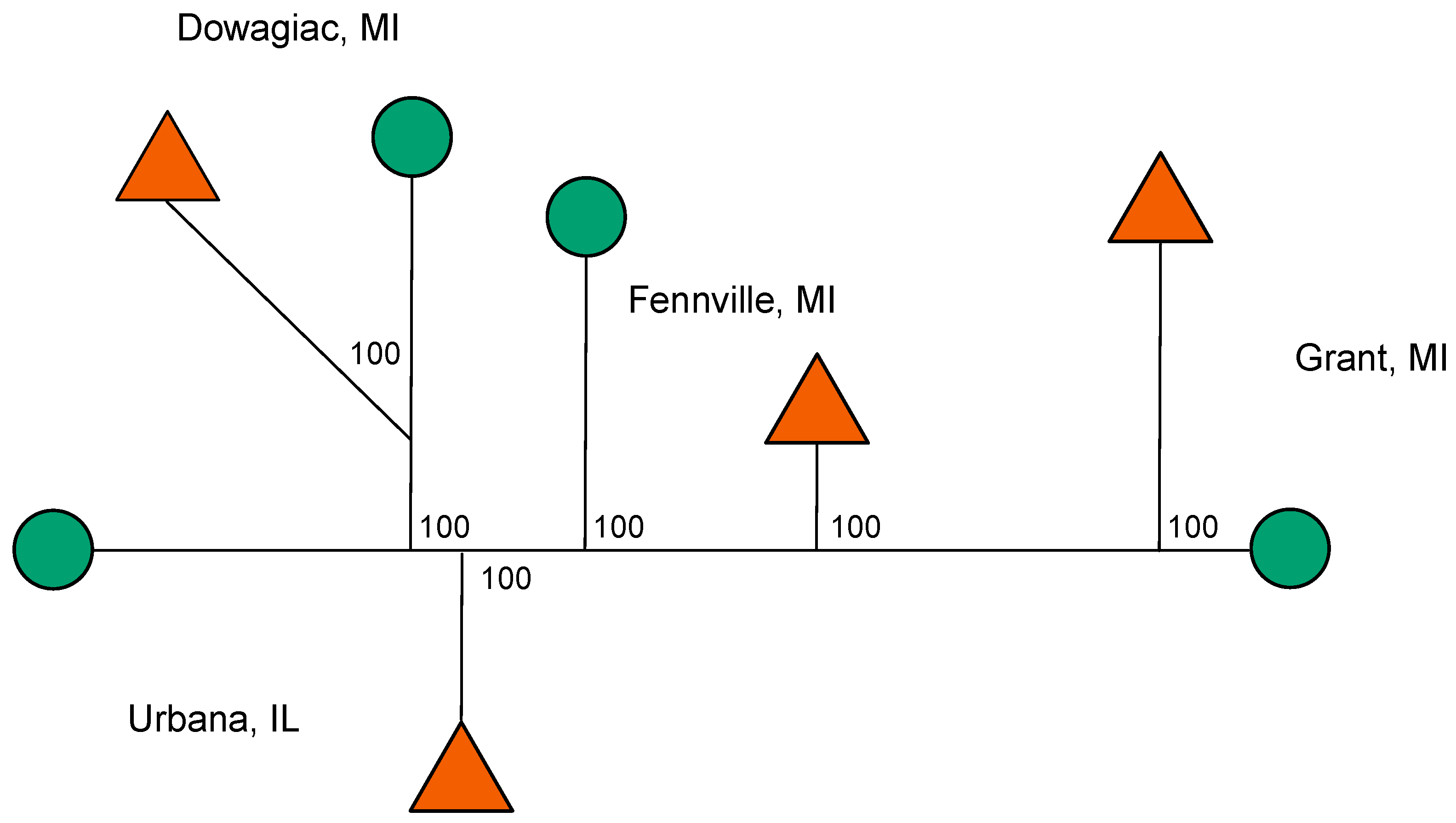
2.1. Geographic Survey
2.2. Eclosion Study
2.3. Prewinter Selection Experiments
2.4. Genotyping-by-Sequencing
2.5. Linkage Disequilibrium and Inversion Polymorphism
2.6. Tests for Genetic Response in Eclosion Time, Prewinter Selection Experiments, Geographic Variation, and Host-Related Differentiation
2.7. Tests for Genetic Associations of Eclosion Time, Prewinter Selection Experiments, Geographic Variation, and Host-Related Differentiation
2.8. Tests for Genetic Clustering among Populations
2.9. Analysis of Linkage Disequilibrium among Unlinked Loci
3. Results
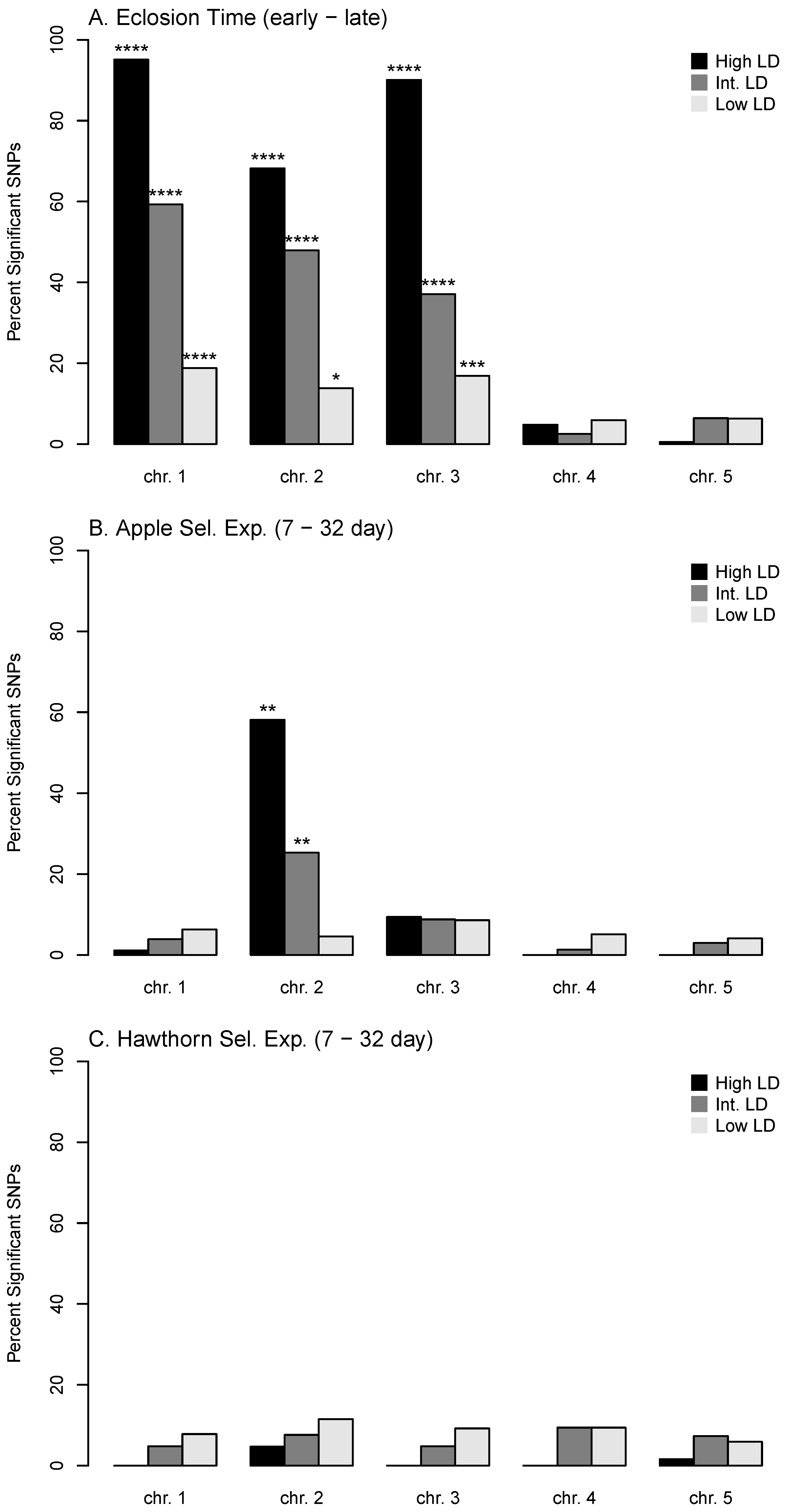
3.1. Genetic Response in the Apple Prewinter Selection Experiment
3.2. Association of Genetic Responses among Apple and Hawthorn Prewinter, and Eclosion Experiments
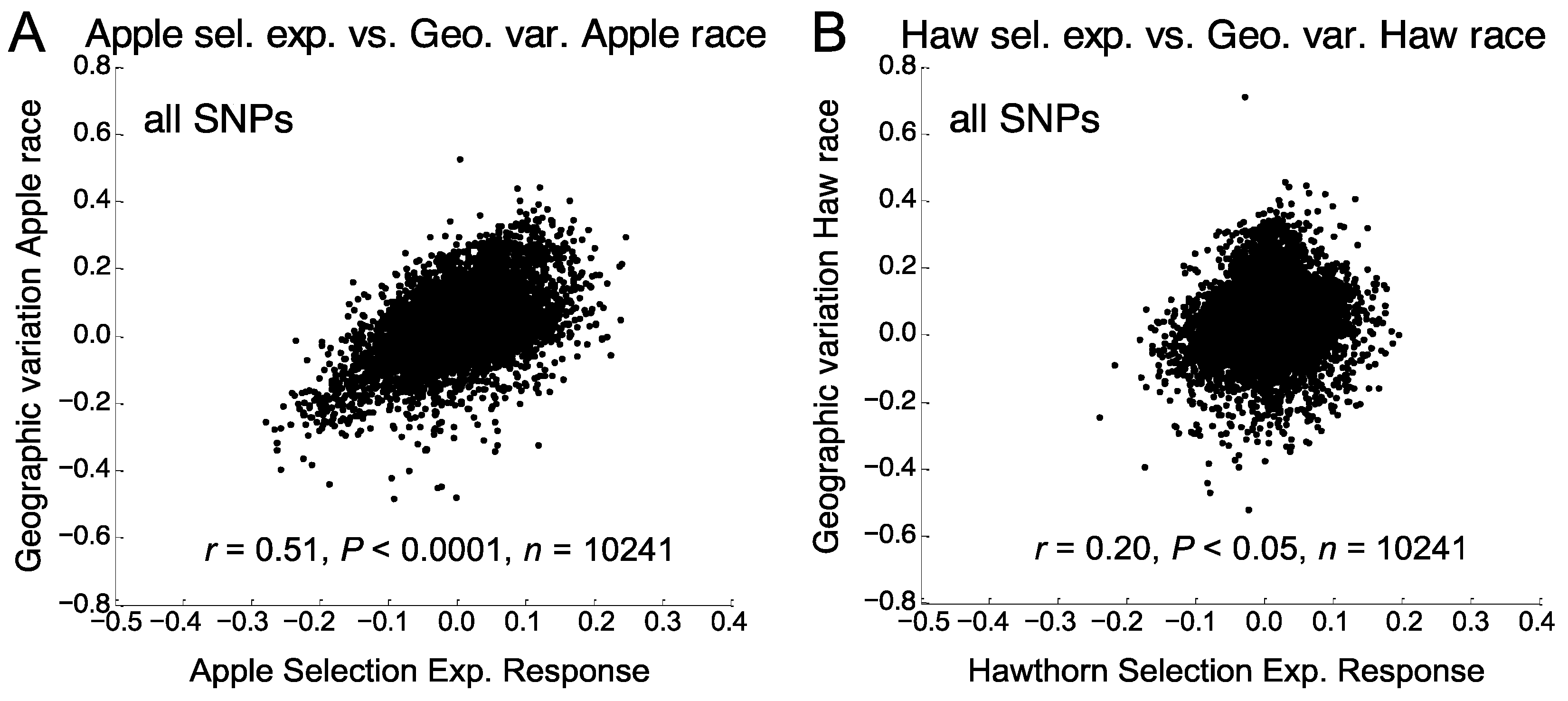
3.3. Relationship of Apple Prewinter Selection Experiment with Geographic Variation
3.4. Relationship of Hawthorn Prewinter Selection Experiments with Geographic Variation
3.5. Relationship of Eclosion Time and Prewinter Selection Experiments with Geographic Variation
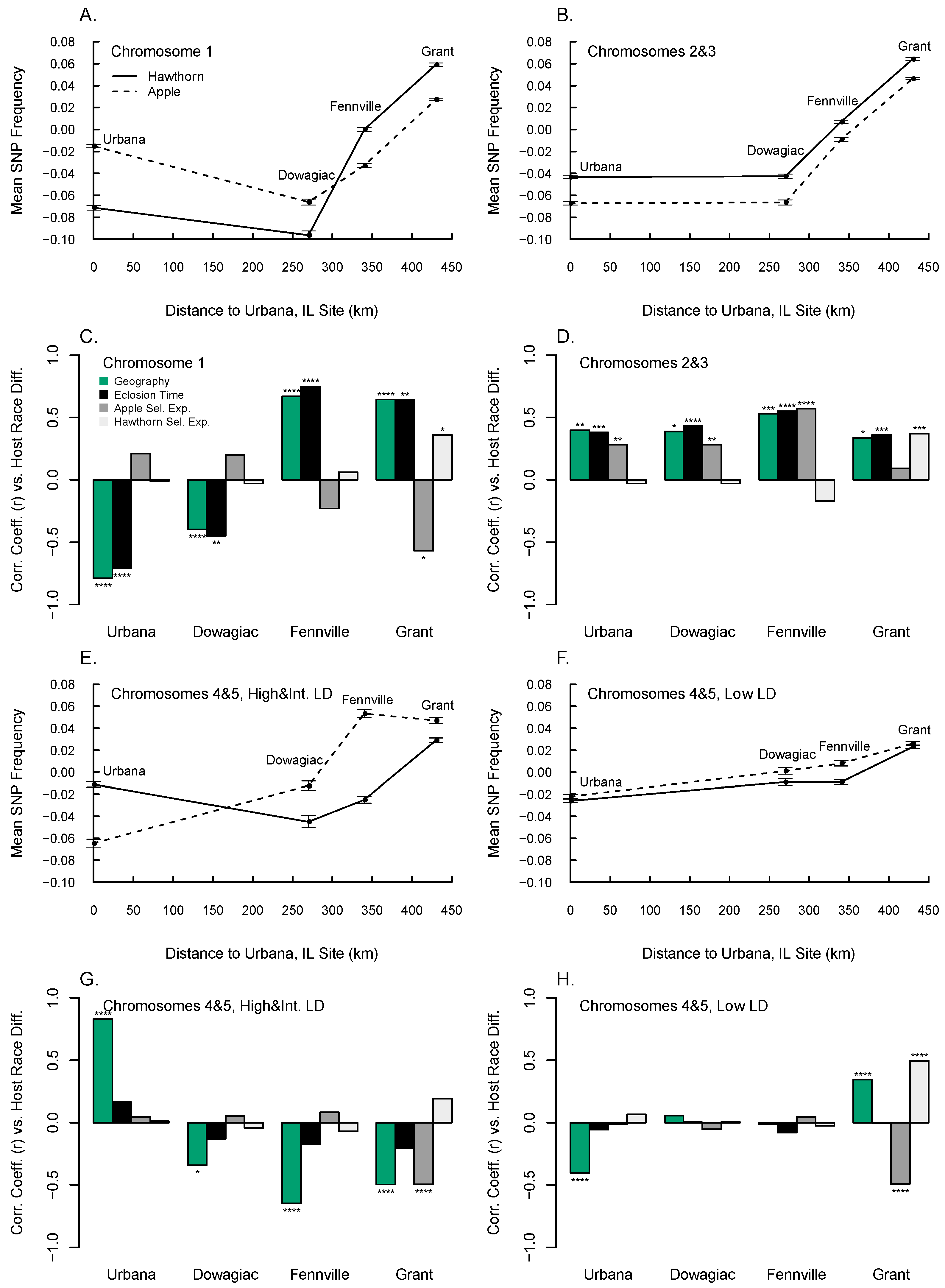
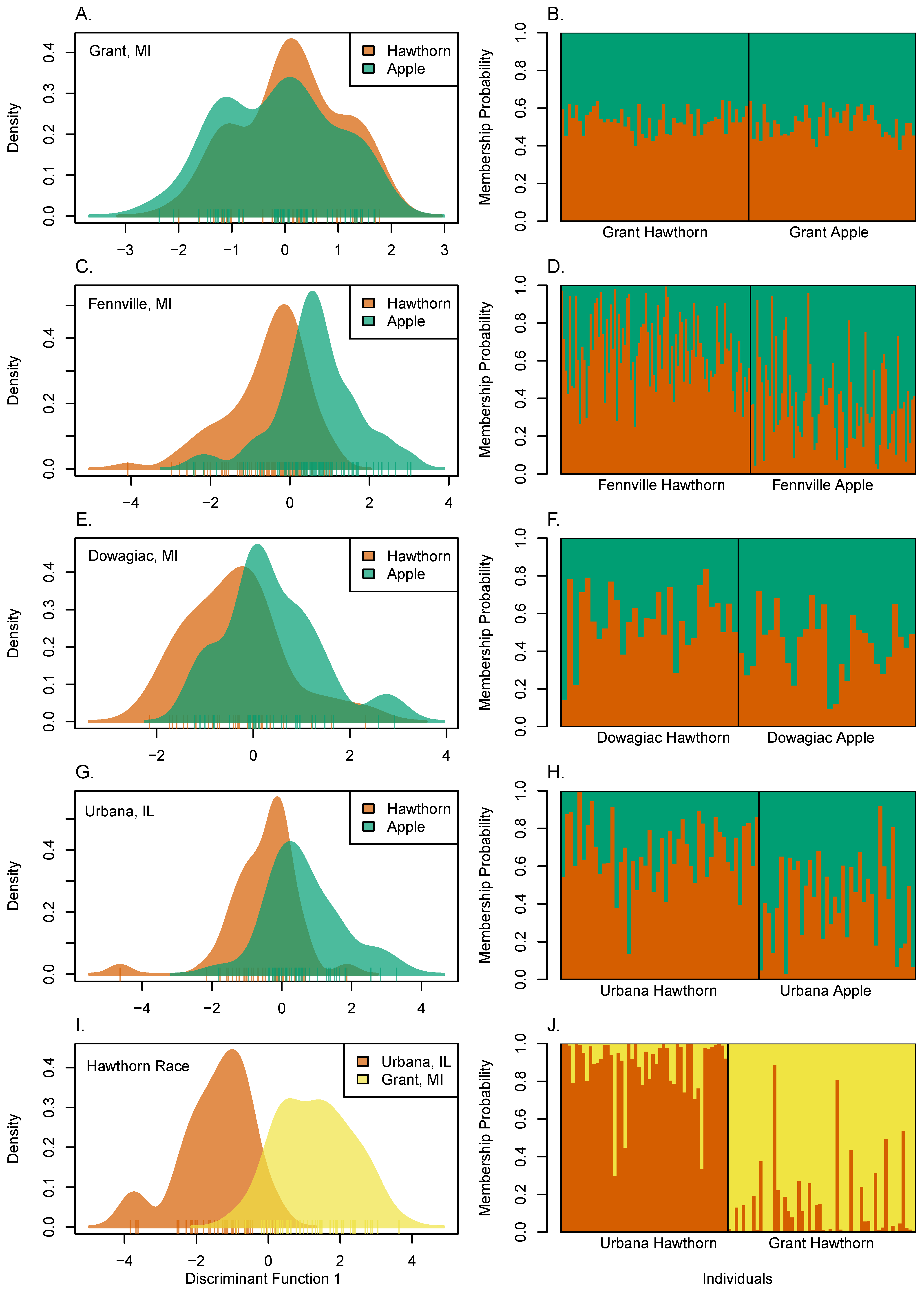
3.6. Diapause, Geographic, and Host Race Differentiation at Sympatric Sites
3.7. Local and Global Patterns of Genetic Differentiation
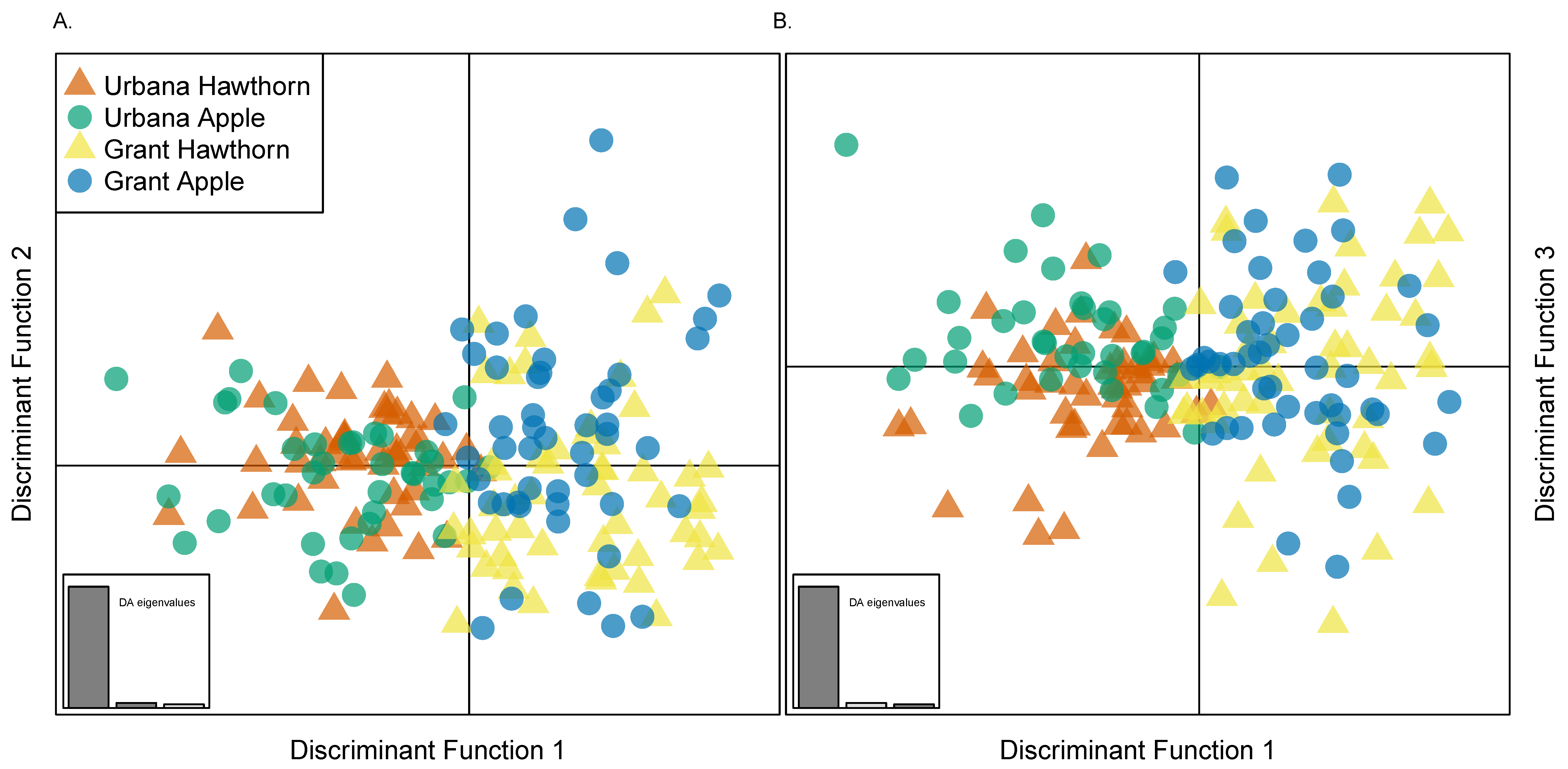
3.8. Genetic Clustering among Populations
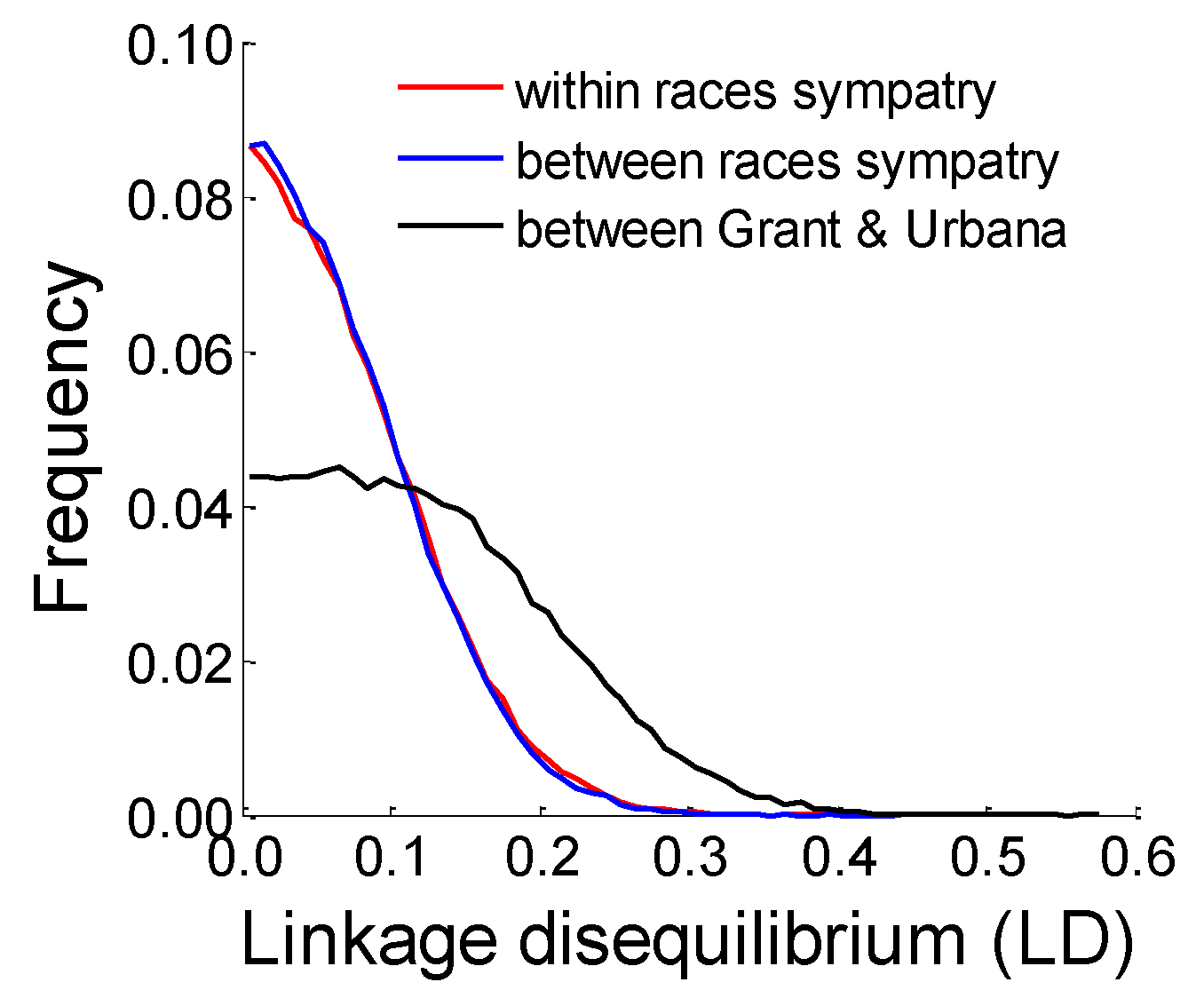
3.9. Analysis of Linkage Disequilibrium among Unlinked Loci
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Data Accessibility
References
- Harrison, R.G. Hybrid zones: Windows on evolutionary process. In Oxford Surveys in Evolutionary Biology; Futuyma, D., Antonovics, J., Eds.; Oxford University Press: New York, NY, USA, 1990; Volume 7, pp. 69–128. [Google Scholar]
- Harrison, R.G. (Ed.) Hybrid Zones and the Evolutionary Process; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Endler, J.A. Geographic Variation, Speciation, and Clines; Princeton University Press: Princeton, NJ, USA, 1977; Volume 10. [Google Scholar]
- Barton, N.H.; Hewitt, G.M. Analysis of hybrid zones. Annu. Rev. Ecol. Syst. 1985, 16, 113–148. [Google Scholar] [CrossRef]
- Szymura, J.M.; Barton, N.H. Genetic analysis of a hybrid zone between the fire-bellied toads, Bombina bombina and B. variegata, near Cracow in Southern Poland. Evolution 1986, 40, 1141–1159. [Google Scholar] [PubMed]
- Rieseberg, L.H.; Whitton, J.; Gardner, K. Hybrid zones and the genetic architecture of a barrier to gene flow between two sunflower species. Genetics 1999, 152, 713–727. [Google Scholar] [PubMed]
- Gompert, Z.; Buerkle, C.A. A powerful regression-based method for admixture mapping of isolation across the genome of hybrids. Mol. Ecol. 2009, 18, 1207–1224. [Google Scholar] [CrossRef] [PubMed]
- Gompert, Z.; Buerkle, C.A. Bayesian estimation of genomic clines. Mol. Ecol. 2011, 20, 2111–2127. [Google Scholar] [CrossRef] [PubMed]
- Payseur, B.A. Using differential introgression in hybrid zones to identify genomic regions involved in speciation. Mol. Ecol. Resour. 2010, 10, 806–820. [Google Scholar] [CrossRef] [PubMed]
- Nachman, M.W.; Payseur, B.A. Recombination rate variation and speciation: Theoretical predictions and empirical results from rabbits and mice. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, B.M. Alternative forms for genomic clines. Ecol. Evol. 2013, 3, 1951–1966. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.G.; Larson, E.L. Heterogeneous genome divergence, differential introgression, and the origin and structure of hybrid zones. Mol. Ecol. 2016, 25, 2454–2466. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.H. Gene flow past a cline. Heredity 1979, 43, 333–339. [Google Scholar] [CrossRef]
- Barton, N.H. Multilocus clines. Evolution 1983, 37, 454–471. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, B. The flow of genes through a genetic barrier. In Evolution: Essays in Honor of John Maynard Smith; Greenwood, P., Slatkin, M., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 31–42. [Google Scholar]
- Barton, N.H.; Bengtsson, B.O. The barrier to genetic exchange between hybridising populations. Heredity 1986, 57, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.H.; de Cara, M.A.R. The evolution of strong reproductive isolation. Evolution 2009, 63, 1171–1190. [Google Scholar] [CrossRef] [PubMed]
- Smadja, C.M.; Butlin, R.K. A framework for comparing processes of speciation in the presence of gene flow. Mol. Ecol. 2011, 20, 5123–5140. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.L.; Mullen, S.P. Genes versus phenotypes in the study of speciation. Genetica 2011, 139, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Bierne, N.; Welch, J.; Loire, E.; Bonhomme, F.; David, P. The coupling hypothesis: Why genome scans may fail to map local adaptation genes. Mol. Ecol. 2011, 20, 2044–2072. [Google Scholar] [CrossRef] [PubMed]
- Bierne, N.; Gagnaire, P.; David, P. The geography of introgression in a patchy environment and the thorn in the side of ecological speciation. Curr. Zool. 2013, 59, 72–86. [Google Scholar] [CrossRef]
- Gompert, Z.; Lucas, L.K.; Buerkle, C.A.; Forister, M.L.; Fordyce, J.A.; Nice, C.C. Admixture and the organization of genetic diversity in a butterfly species complex revealed through common and rare genetic variants. Mol. Ecol. 2014, 23, 4555–4573. [Google Scholar] [CrossRef] [PubMed]
- Gompert, Z.; Egan, S.P.; Barrett, R.D.H.; Feder, J.L.; Nosil, P. Multilocus approaches for the measurement of selection on correlated genetic loci. Mol. Ecol. 2017, 26, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Gompert, Z.; Comeault, A.A.; Farkas, T.E.; Feder, J.L.; Parchman, T.L.; Buerkle, C.A.; Nosil, P. Experimental evidence for ecological selection on genome variation in the wild. Ecol. Lett. 2014, 17, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Soria-Carrasco, V.; Gompert, Z.; Comeault, A.A.; Farkas, T.E.; Parchman, T.L.; Johnston, J.S.; Buerkle, C.A.; Feder, J.L.; Bast, J.; Schwander, T.; et al. Stick insect genomes reveal natural selection’s role in parallel speciation. Science 2014, 344, 738–742. [Google Scholar] [CrossRef] [PubMed]
- De Villemereuil, P.; Gaggiotti, O.E.; Mouterde, M.; Till-Bottraud, I. Common garden experiments in the genomic era: New perspectives and opportunities. Heredity 2016, 116, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Villoutreix, R.; de Carvalho, C.F.; Farkas, T.E.; Soria-Carrasco, V.; Feder, J.L.; Crespi, B.J.; Gompert, Z. Natural selection and the predictability of evolution in Timema stick insects. Science 2018, 359, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Hybrid zones-natural laboratories for evolutionary studies. Trends Ecol. Evol. 1988, 3, 158–167. [Google Scholar] [CrossRef]
- Ragland, G.J.; Doellman, M.M.; Meyers, P.J.; Hood, G.R.; Egan, S.P.; Powell, T.H.Q.; Hahn, D.A.; Nosil, P.; Feder, J.L. A test of genomic modularity among life-history adaptations promoting speciation with gene flow. Mol. Ecol. 2017, 26, 3926–3942. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.P.; Ragland, G.J.; Assour, L.; Powell, T.H.Q.; Hood, G.R.; Emrich, S.; Nosil, P.; Feder, J.L. Experimental evidence of genome-wide impact of ecological selection during early stages of speciation-with-gene-flow. Ecol. Lett. 2015, 18, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Bush, G. The taxonomy, cytology, and evolution of the genus Rhagoletis in North America (Diptera, Tephritidae). Bull. Mus. Comp. Zool. Harvard Univ. 1966, 134, 431–562. [Google Scholar]
- Bush, G.L. Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera, Tephritidae). Evolution 1969, 23, 237. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.L.; Chilcote, C.A.; Bush, G.L. Genetic differentiation between sympatric host races of the apple maggot fly Rhagoletis pomonella. Nature 1988, 336, 61–64. [Google Scholar] [CrossRef]
- Feder, J.L.; Hunt, T.; Bush, G. The effects of climate, host-plant phenology and host fidelity on the genetics of apple and hawthorn infesting races of Rhagoletis pomonella. Entomol. Exp. Appl. 1993, 69, 117–135. [Google Scholar] [CrossRef]
- Feder, J.L.; Opp, S.B.; Wlazlo, B.; Reynolds, K.; Go, W.; Spisak, S. Host fidelity is an effective premating barrier between sympatric races of the apple maggot fly. Proc. Natl. Acad. Sci. USA 1994, 91, 7990–7994. [Google Scholar] [CrossRef] [PubMed]
- Reissig, W.H.; Smith, D.C. Bionomics of Rhagoletis pomonella in Crataegus. Ann. Entomol. Soc. Am. 1978, 71, 155–159. [Google Scholar] [CrossRef]
- Noor, M.A.F.; Bennett, S.M. Islands of speciation or mirages in the desert? Examining the role of restricted recombination in maintaining species. Heredity 2009, 103, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, T.E.; Hahn, M.W. Reanalysis suggests that genomic islands of speciation are due to reduced diversity, not reduced gene flow. Mol. Ecol. 2014, 23. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.; Tauber, C.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef] [PubMed]
- Dopman, E.B.; Robbins, P.S.; Seaman, A. Components of reproductive isolation between North American pheromone strains of the European corn borer. Evolution 2009, 64, 881–902. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilization. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.A.; Denlinger, D.L. Energetics of insect diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.W.; Chapman, P.J. Bionomics of the Apple Maggot in Eastern New York; Cornell University: Ithaca, NY, USA, 1973. [Google Scholar]
- Feder, J.L.; Roethele, J.B.; Wlazlo, B.; Berlocher, S.H. Selective maintenance of allozyme differences among sympatric host races of the apple maggot fly. Proc. Natl. Acad. Sci. USA 1997, 94, 11417–11421. [Google Scholar] [CrossRef] [PubMed]
- Filchak, K.E.; Roethele, J.B.; Feder, J.L. Natural selection and sympatric divergence in the apple maggot Rhagoletis pomonella. Nature 2000, 407, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Prokopy, R.J. Influence of photoperiod, temperature, and food on initiation of diapause in the apple maggot. Can. Entomol. 1968, 100, 318–329. [Google Scholar] [CrossRef]
- Lyons-Sobaski, S.; Berlocher, S.H. Life history phenology differences between southern and northern populations of the apple maggot fly, Rhagoletis pomonella. Entomol. Exp. Appl. 2009, 130, 149–159. [Google Scholar] [CrossRef]
- Feder, J.L.; Berlocher, S.H.; Roethele, J.B.; Dambroski, H.; Smith, J.J.; Perry, W.L.; Gavrilovic, V.; Filchak, K.E.; Rull, J.; Aluja, M. Allopatric genetic origins for sympatric host-plant shifts and race formation in Rhagoletis. Proc. Natl. Acad. Sci. USA 2003, 100, 10314–10319. [Google Scholar] [CrossRef] [PubMed]
- Dambroski, H.R.; Feder, J.L. Host plant and latitude-related diapause variation in Rhagoletis pomonella: A test for multifaceted life history adaptation on different stages of diapause development. J. Evol. Biol. 2007, 20, 2101–2112. [Google Scholar] [CrossRef] [PubMed]
- Doellman, M.; Egan, S.; Ragland, G.; Meyers, P.; Hood, G.; Powell, T.; Lazorchak, P.; Hahn, D.; Berlocher, S.; Nosil, P.; et al. Divergent natural selection on eclosion time affects genome wide patterns of geographic and host related differentiation for apple and hawthorn races of Rhagoletis pomonella flies. (manuscript in preparation).
- Feder, J.L.; Xie, X.; Rull, J.; Velez, S.; Forbes, A.; Leung, B.; Dambroski, H.; Filchak, K.E.; Aluja, M. Mayr, Dobzhansky, and Bush and the complexities of sympatric speciation in Rhagoletis. Proc. Natl. Acad. Sci. USA 2005, 102, 6573–6580. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Rull, J.; Michel, A.P.; Velez, S.; Forbes, A.A.; Lobo, N.F.; Aluja, M.; Feder, J.L. Hawthorn-infesting populations of Rhagoletis pomonella in Mexico and speciation mode plurality. Evolution 2007, 61, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.P.; Rull, J.; Aluja, M.; Feder, J.L. The genetic structure of hawthorn-infesting Rhagoletis pomonella populations in Mexico: Implications for sympatric host race formation. Mol. Ecol. 2007, 16, 2867–2878. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Michel, A.P.; Schwarz, D.; Rull, J.; Velez, S.; Forbes, A.A.; Aluja, M.; Feder, J.L. Radiation and divergence in the Rhagoletis pomonella species complex: Inferences from DNA sequence data. J. Evol. Biol. 2008, 21, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J. A species definition for the modern synthesis. Trends Ecol. Evol. 1995, 10, 294–299. [Google Scholar] [CrossRef]
- Riesch, R.; Muschick, M.; Lindtke, D.; Villoutreix, R.; Comeault, A.A.; Farkas, T.E.; Lucek, K.; Hellen, E.; Soria-Carrasco, V.; Dennis, S.R.; et al. Transitions between phases of genomic differentiation during stick-insect speciation. Nat. Ecol. Evol. 2017, 1, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Nosil, P.; Feder, J.L.; Flaxman, S.M.; Gompert, Z. Tipping points in the dynamics of speciation. Nat. Ecol. Evol. 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.L.; Egan, S.P.; Nosil, P. The genomics of speciation-with-gene-flow. Trends Genet. 2012, 28, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Butlin, R.K.; Smadja, C.M. Coupling, reinforcement, and speciation. Am. Nat. 2017, 191. [Google Scholar] [CrossRef] [PubMed]
- Schilling, M.; Mullen, S.; Kronforst, M.; Safran, R.; Nosil, P.; Feder, J.; Gompert, Z.; Flaxman, S. Transitions from single- to multi-locus processes during speciation. Genes 2018. [Google Scholar] [CrossRef]
- Flaxman, S.M.; Feder, J.L.; Nosil, P. Genetic hitchhiking and the dynamic buildup of genomic divergence during speciation with gene flow. Evolution 2013, 67, 2577–2591. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.L.; Nosil, P.; Wacholder, A.C.; Egan, S.P.; Berlocher, S.H.; Flaxman, S.M. Genome-wide congealing and rapid transitions across the speciation continuum during speciation with gene flow. J. Hered. 2014, 105, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, S.M.; Wacholder, A.C.; Feder, J.L.; Nosil, P. Theoretical models of the influence of genomic architecture on the dynamics of speciation. Mol. Ecol. 2014, 23, 4074–4088. [Google Scholar] [CrossRef] [PubMed]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Feder, J.L.; Roethele, J.B.; Filchak, K.; Niedbalski, J.; Romero-Severson, J. Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella. Genetics 2003, 163, 939–953. [Google Scholar] [PubMed]
- Michel, A.P.; Sim, S.; Powell, T.H.Q.; Taylor, M.S.; Nosil, P.; Feder, J.L. Widespread genomic divergence during sympatric speciation. Proc. Natl. Acad. Sci. USA 2010, 107, 9724–9729. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S. Inferences about linkage disequilibrium. Biometrics 1979, 35, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, P.; Knight, C.G.; Sarma, D.K.; Hlaing, T.; Prakash, A.; Maung Maung, Y.N.; Somboon, P.; Mahanta, J.; Walton, C. Linkage disequilibrium network analysis (LDna) gives a global view of chromosomal inversions, local adaptation and geographic structure. Mol. Ecol. Resour. 2015, 15, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org (accessed on 10 January 2018).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Brooks, J.C.; Grünwald, N.J. Novel R tools for analysis of genome-wide population genetic data with emphasis on clonality. Front. Genet. 2015, 6, 208. [Google Scholar] [CrossRef] [PubMed]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K.; Paradis, E.; Claude, J.; Strimmer, K.; Schliep, K.; Revell, L.; Faith, D.; Isaac, N.; et al. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Ragland, G.J.; Fuller, J.; Feder, J.L.; Hahn, D.A. Biphasic metabolic rate trajectory of pupal diapause termination and post-diapause development in a tephritid fly. J. Insect Physiol. 2009, 55, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Berlocher, S.H.; Feder, J.L. Sympatric speciation in phytophagous insects: Moving beyond controversy? Annu. Rev. Entomol. 2002, 47, 773–815. [Google Scholar] [CrossRef] [PubMed]
- Barrett, R.D.H.; Schluter, D. Adaptation from standing genetic variation. Trends Ecol. Evol. 2008, 23, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.J.; Barton, N.H.; Good, J.M. Genomics of hybridization and its evolutionary consequences. Mol. Ecol. 2016, 25, 2325–2332. [Google Scholar] [CrossRef] [PubMed]
- Payseur, B.A.; Rieseberg, L.H. A genomic perspective on hybridization and speciation. Mol. Ecol. 2016, 25, 2337–2360. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apple Race | chr 1 | chr 2 | chr 3 | chr 4 | chr 5 | chr 1–5 |
| Map SNP | 10 A | 39 EA | 43 EA | 29 A | 9 | 24 EA |
| High LD | 7 EA | 57 EA | 86 EA | 79 A | 7 A | 33 EA |
| Int. LD | 11 A | 39 EA | 35 EA | 40 | 11 | 24 EA |
| Low LD | 10 A | 15 EA | 15 A | 13 A | 8 | 11 A |
| Haw Race | chr 1 | chr 2 | chr 3 | chr 4 | chr 5 | chr 1–5 |
| Map SNP | 54 E | 50 EA | 48 EA | 12 | 15 | 37 EAH |
| High LD | 92 E | 82 EA | 84 EH | 5 | 17 | 58 EA |
| Int. LD | 46 E | 47 EA | 42 EAH | 12 | 14 | 35 EA |
| Low LD | 16 E | 21 EA | 21 H | 15 H | 16 H | 17 EH |
| Hawthorn Race | ||||||
| Predictor | R2 | Coeff. | SE | t | F4,4239 | p |
| Eclosion Exp. | 0.629 | 0.013 | 49.5 | <0.0001 | ||
| Hawthorn Sel. Exp. | 0.413 | 0.023 | 17.7 | <0.0001 | ||
| Apple Sel. Exp. | 0.258 | 0.016 | 16.6 | <0.0001 | ||
| LD | 0.036 | 0.005 | 6.8 | <0.0001 | ||
| Total | 0.540 | 1248 | <0.0001 | |||
| Apple Race | ||||||
| Predictor | R2 | Coeff. | SE | t | F3,4240 | p |
| Eclosion Exp. | 0.185 | 0.014 | 13.4 | <0.0001 | ||
| Apple Sel. Exp. | 0.698 | 0.017 | 40.9 | <0.0001 | ||
| LD | 0.046 | 0.006 | 8.2 | <0.0001 | ||
| Total | 0.3824 | 876.8 | <0.0001 |
| Grant, MI | ||||||
| Predictor | R2 | Coeff. | SE | t | F4,4239 | p |
| Eclosion Exp. | 0.236 | 0.008 | 31.3 | <0.0001 | ||
| Hawthorn Sel. Exp. | 0.394 | 0.014 | 28.5 | <0.0001 | ||
| Apple Sel. Exp. | −0.154 | 0.009 | −16.7 | <0.0001 | ||
| LD | 0.008 | 0.003 | 2.5 | 0.012 | ||
| Total | 0.348 | 565.8 | <0.0001 | |||
| Fennville, MI | ||||||
| Predictor | R2 | Coeff. | SE | t | F4,4239 | p |
| Eclosion Exp. | 0.371 | 0.012 | 31.6 | <0.0001 | ||
| Hawthorn Sel. Exp. | −0.061 | 0.022 | −2.8 | 0.0048 | ||
| Apple Sel. Exp. | 0.050 | 0.014 | 3.5 | 0.0005 | ||
| LD | 0.021 | 0.005 | 4.3 | <0.0001 | ||
| Total | 0.288 | 429.4 | <0.0001 | |||
| Dowagiac, MI | ||||||
| Predictor | R2 | Coeff. | SE | t | F2,4241 | p |
| Eclosion Exp. | −0.034 | 0.012 | −2.8 | 0.0051 | ||
| Apple Sel. Exp. | 0.239 | 0.017 | 13.9 | <0.0001 | ||
| Total | 0.043 | 96.1 | <0.0001 | |||
| Urbana, IL | ||||||
| Predictor | R2 | Coeff. | SE | t | F3,4240 | p |
| Eclosion Exp. | −0.217 | 0.013 | −16.1 | <0.0001 | ||
| Apple Sel. Exp. | 0.281 | 0.017 | 17.0 | <0.0001 | ||
| LD | 0.020 | 0.005 | 3.6 | <0.0001 | ||
| Total | 0.097 | 153.5 | 0.0004 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doellman, M.M.; Ragland, G.J.; Hood, G.R.; Meyers, P.J.; Egan, S.P.; Powell, T.H.Q.; Lazorchak, P.; Glover, M.M.; Tait, C.; Schuler, H.; et al. Genomic Differentiation during Speciation-with-Gene-Flow: Comparing Geographic and Host-Related Variation in Divergent Life History Adaptation in Rhagoletis pomonella. Genes 2018, 9, 262. https://doi.org/10.3390/genes9050262
Doellman MM, Ragland GJ, Hood GR, Meyers PJ, Egan SP, Powell THQ, Lazorchak P, Glover MM, Tait C, Schuler H, et al. Genomic Differentiation during Speciation-with-Gene-Flow: Comparing Geographic and Host-Related Variation in Divergent Life History Adaptation in Rhagoletis pomonella. Genes. 2018; 9(5):262. https://doi.org/10.3390/genes9050262
Chicago/Turabian StyleDoellman, Meredith M., Gregory J. Ragland, Glen R. Hood, Peter J. Meyers, Scott P. Egan, Thomas H. Q. Powell, Peter Lazorchak, Mary M. Glover, Cheyenne Tait, Hannes Schuler, and et al. 2018. "Genomic Differentiation during Speciation-with-Gene-Flow: Comparing Geographic and Host-Related Variation in Divergent Life History Adaptation in Rhagoletis pomonella" Genes 9, no. 5: 262. https://doi.org/10.3390/genes9050262
APA StyleDoellman, M. M., Ragland, G. J., Hood, G. R., Meyers, P. J., Egan, S. P., Powell, T. H. Q., Lazorchak, P., Glover, M. M., Tait, C., Schuler, H., Hahn, D. A., Berlocher, S. H., Smith, J. J., Nosil, P., & Feder, J. L. (2018). Genomic Differentiation during Speciation-with-Gene-Flow: Comparing Geographic and Host-Related Variation in Divergent Life History Adaptation in Rhagoletis pomonella. Genes, 9(5), 262. https://doi.org/10.3390/genes9050262