Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata



Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Pipelines for Repeat Identification and Repeat Landscape Construction
2.3. Comparative Analysis Pipeline
2.4. Transcriptome Analysis
2.5. Experimental Validation of TEs
3. Results
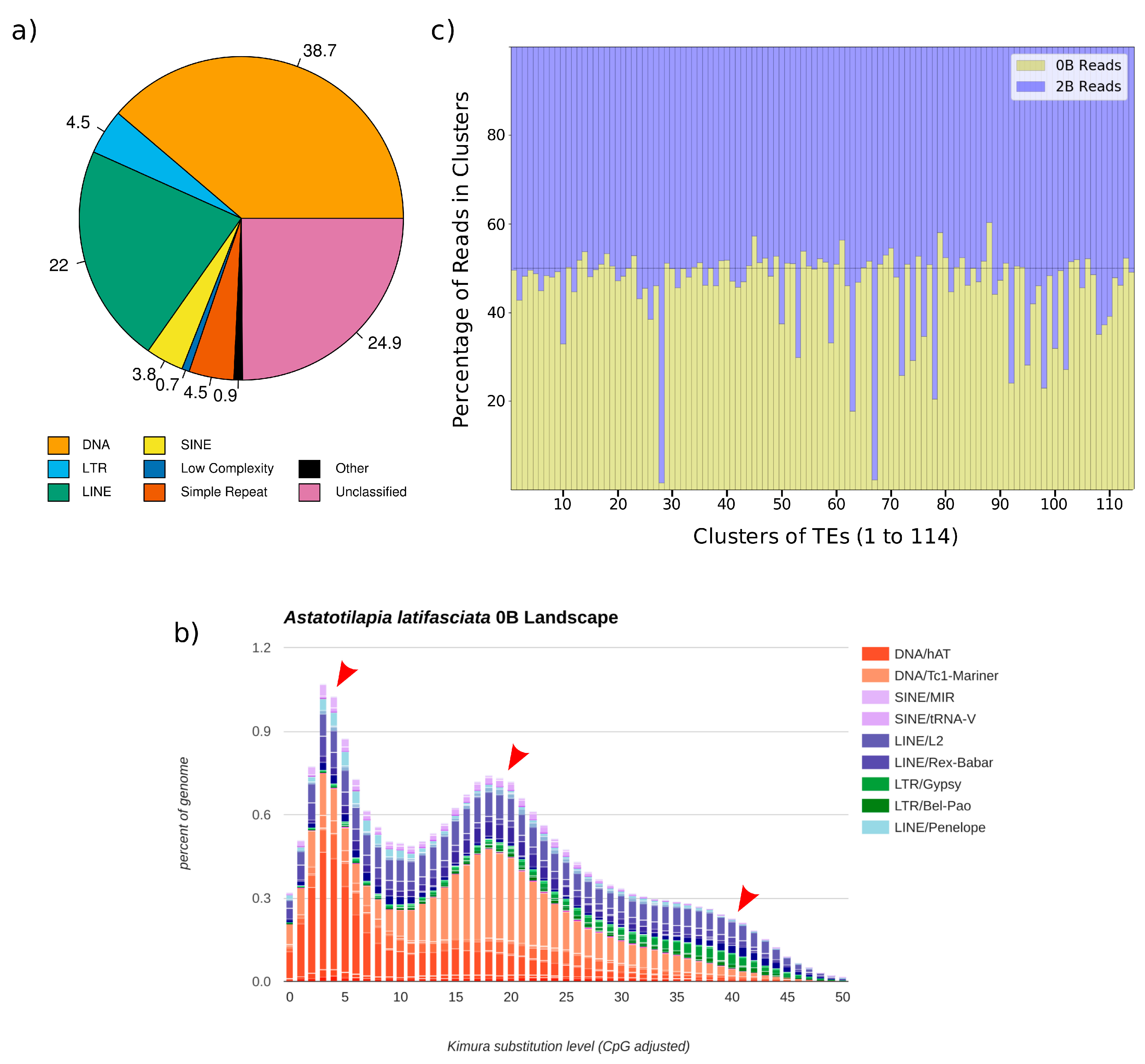
3.1. DNA Transposable Elements Are the Most Represented Class in the A. latifasciata Genome
3.2. Three Main Bursts of DNA Element Insertion during Evolution Explain Their Genomic Abundance
3.3. Accumulation of TEs in the B Chromosome of A. latifasciata
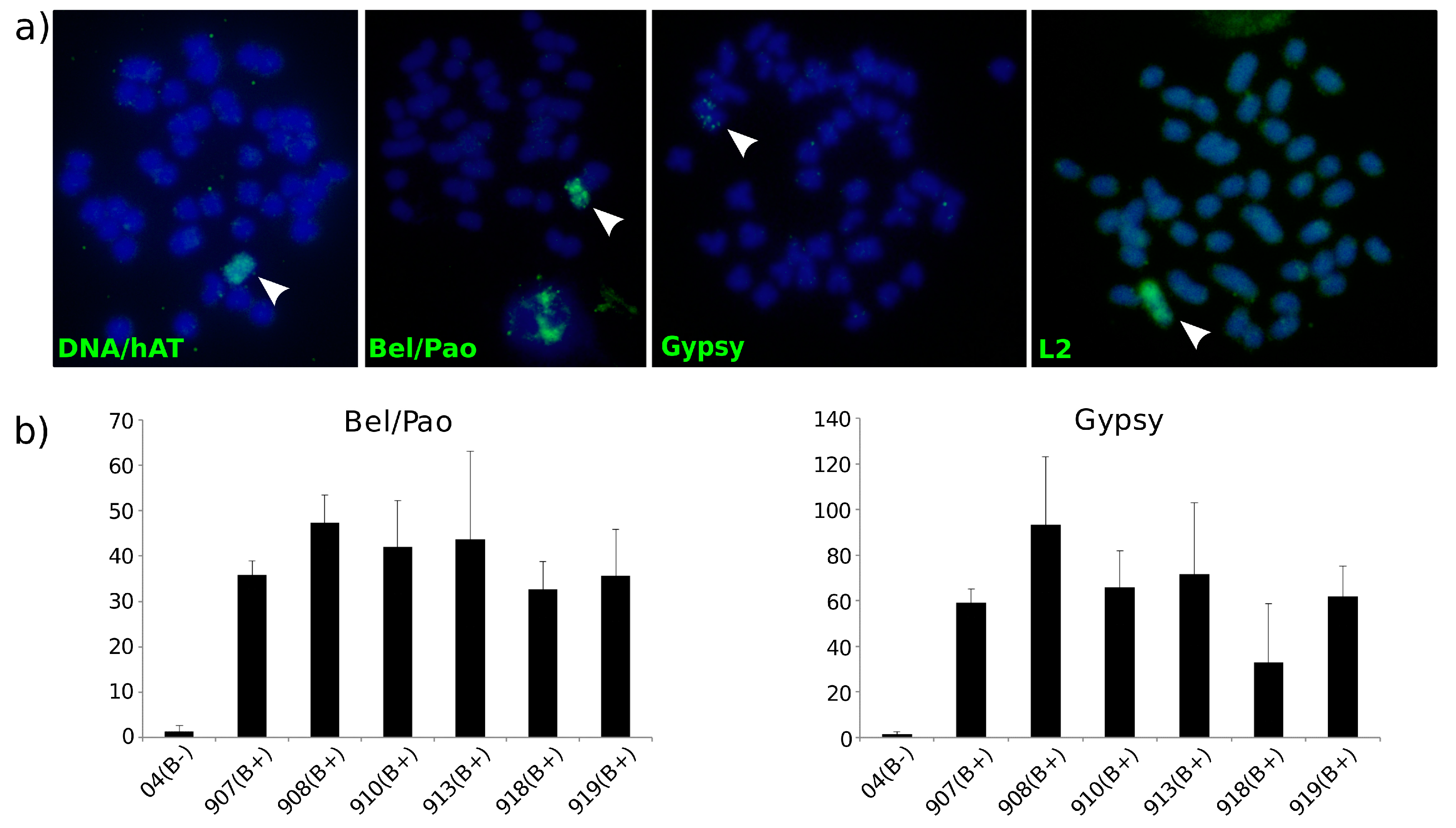
3.4. B Chromosome TE Accumulation Is Validated by Cytogenetics and Molecular Techniques
3.5. TE Transcription across Tissues and Higher Expression in B Chromosome Samples
4. Discussion
4.1. Genomic Organization of TEs in A. latifasciata
4.2. Organization and Transcription of TEs in the B Chromosome
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Camacho, J.P.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Kuroiwa, A.; Hirai, H.; Hirai, Y.; Matsuda, Y.; et al. B chromosomes have a functional effect on female sex determination in Lake Victoria cichlid fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef] [PubMed]
- Houben, A.; Banaei-Moghaddam, A. Evolution and biology of supernumerary B chromosomes. Cell. Mol. Life Sci. 2014, 71, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Ploskaya-Chaibi, M.; Voitovich, A.M.; Novitsky, R.V.; Bouhadad, R. B-chromosome and V-shaped spot asymmetry in the common frog (Rana temporaria L.) populations. Comptes Rendus Biol. 2015, 338, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Beukeboom, L.W. Bewildering Bs: An impression of the 1st B-Chromosome conference. Heredity 1994, 73, 328–336. [Google Scholar] [CrossRef]
- Carchilan, M.; Kumke, K.; Mikolajewski, S.; Houben, A. Rye B chromosomes are weakly transcribed and might alter the transcriptional activity of a chromosome sequences. Chromosoma 2009, 118, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Adnađević, T.; Jovanović, V.M.; Blagojević, J.; Budinski, I.; Cabrilo, B.; Bijelić-Čabrilo, O.; Vujošević, M. Possible influence of B Chromosomes on genes included in immune response and parasite burden in Apodemus flavicollis. PLoS ONE 2014, 9, e112260. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Nakajima, R.T.; Fantinatti, B.E.A.; Marques, D.F.; Almeida, R.O.; Simões, R.P.; Martins, C. B chromosomes: From cytogenetics to systems biology. Chromosoma 2017, 126, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Moghaddam, A.M.; Martis, M.M.; Macas, J.; Gundlach, H.; Himmelbach, A.; Altschmied, L.; Mayer, K.F.X.; Houben, A. Genes on B chromosomes: Old questions revisited with new tools. Biochim. Biophys. Acta 2015, 1849, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Conte, M.A.; Fantinatti, B.E.A.; Cabral-de-Mello, D.C.; Carvalho, R.F.; Vicari, M.R.; Kocher, T.D.; Martins, C. Origin and evolution of B chromosomes in the cichlid fish Astatotilapia latifasciata based on integrated genomic analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.M.A.; Pansonato-Alves, J.C.; Utsunomia, R.; Araya-Jaime, C.; Ruiz-Ruano, F.J.; Daniel, S.N.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Porto-Foresti, F.; et al. Delimiting the origin of a B chromosome by FISH mapping, chromosome painting and DNA sequence analysis in Astyanax paranae (Teleostei, Characiformes). PLoS ONE 2014, 9, e94896. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Sniegowski, P.; Stephan, W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 1994, 371, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A.; von Sternberg, R. Why repetitive DNA is essential to genome function. Biol. Rev. 2005, 80, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Rebollo, R.; Romanish, M.T.; Mager, D.L. Transposable elements: An abundant and natural source of regulatory sequences for host genes. Annu. Rev. Genet. 2012, 46, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Bueno, D.; Palacios-Gimenez, O.M.; Cabral-de-Mello, D.C. Chromosomal mapping of repetitive DNAs in the grasshopper Abracris flavolineata reveal possible ancestry of the B chromosome and H3 histone spreading. PLoS ONE 2013, 8, e66532. [Google Scholar] [CrossRef] [PubMed]
- Teruel, M.; Cabrero, J.; Perfectti, F.; Camacho, J.P.M. B chromosome ancestry revealed by histone genes in the migratory locust. Chromosoma 2010, 119, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Cabral-de-Mello, D.C.; Moura, R.C.; Martins, C. Chromosomal mapping of repetitive DNAs in the beetle Dichotomius geminatus provides the first evidence for an association of 5S rRNA and histone H3 genes in insects, and repetitive DNA similarity between the B chromosome and A complement. Heredity 2010, 104, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Fantinatti, B.E.; Mazzuchelli, J.; Valente, G.T.; Cabral-de-Mello, D.C.; Martins, C. Genomic content and new insights on the origin of the B chromosome of the cichlid fish Astatotilapia latifasciata. Genetica 2011, 139, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Utsunomia, R.; de Andrade Silva, D.M.Z.; Ruiz-Ruano, F.J.; Araya-Jaime, C.; Pansonato-Alves, J.C.; Scacchetti, P.C.; Hashimoto, D.T.; Oliveira, C.; Trifonov, V.A.; Porto-Foresti, F.; et al. Uncovering the ancestry of B chromosomes in Moenkhausia sanctaefilomenae (Teleostei, Characidae). PLoS ONE 2016, 11, e0150573. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.M.Z.; Daniel, S.N.; Camacho, J.P.M.; Utsunomia, R.; Ruiz-Ruano, F.J.; Penitente, M.; Pansonato-Alves, J.C.; Hashimoto, D.T.; et al. Origin of B chromosomes in the genus Astyanax (Characiformes, Characidae) and the limits of chromosome painting. Mol. Genet. Genom. 2016, 291, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Martis, M.M.; Klemme, S.; Banaei-Moghaddam, A.M.; Blattner, F.R.; Macas, J.; Schmutzer, T.; Scholz, U.; Gundlach, H.; Wicker, T.; Šimková, H.; et al. Selfish supernumerary chromosome reveals its origin as a mosaic of host genome and organellar sequences. Proc. Natl. Acad. Sci. USA 2012, 109, 13343–13346. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.; Lamatsch, D.K.; Steinlein, C.; Engel, W.; Schartl, M.; Schmid, M. The giant B chromosome of the cyprinid fish Alburnus alburnus harbours a retrotransposon-derived repetitive DNA sequence. Chromosome Res. 2003, 11, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.C.; Riddle, N.C.; Cheng, Y.M.; Theuri, J.; Birchler, J.A. Localization and transcription of a retrotransposon-derived element on the maize B chromosome. Chromosome Res. 2007, 15, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Du, Y.; Zhao, X.; Jin, W. B chromosome contains active genes and impacts the transcription of A chromosomes in maize (Zea mays L.). BMC Plant Biol. 2016, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Gabriel, T.S.; Martis, M.M.; Gursinsky, T.; Schubert, V.; Vrána, J.; Doležel, J.; Grundlach, H.; Altschmied, L.; Scholz, U.; et al. Rye B chromosomes encode a functional Argonaute-like protein with in vitro slicer activities similar to its A chromosome paralog. New Phytol. 2017, 213, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Poletto, A.B.; Ferreira, I.A.; Martins, C. The B chromosomes of the African cichlid fish Haplochromis obliquidens harbour 18S rRNA gene copies. BMC Genet. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, A.; Terai, Y.; Kobayashi, N.; Yoshida, K.; Suzuki, M.; Nakanishi, A.; Matsuda, Y.; Watanabe, M.; Okada, N. Construction of chromosome markers from the Lake Victoria cichlid Paralabidochromis chilotes and their application to comparative mapping. Cytogenet. Genome Res. 2014, 142, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Clark, F.E.; Conte, M.A.; Ferreira-Bravo, I.A.; Poletto, A.B.; Martins, C.; Kocher, T.D. Dynamic sequence evolution of a sex-associated B chromosome in Lake Malawi cichlid fish. J. Hered. 2017, 108, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Turner, G.F.; Seehausen, O.; Knight, M.E.; Allender, C.J.; Robinson, R.L. How many species of cichlid fishes are there in African lakes? Mol. Ecol. 2001, 10, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Kocher, T.D. Adaptive evolution and explosive speciation: The cichlid fish model. Nat. Rev. Genet. 2004, 5, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Seehausen, O. African cichlid fish: A model system in adaptive radiation research. Proc. Biol. Sci. 2006, 273, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Fantinatti, B.E.A.; Martins, C. Development of chromosomal markers based on next-generation sequencing: The B chromosome of the cichlid fish Astatotilapia latifasciata as a model. BMC Genet. 2016, 17, 119. [Google Scholar] [CrossRef] [PubMed]
- Jehangir, M. Genome Assembly of the Cichlid Fish Astatotilapia latifasciata with Focus in Population Genomics of B Chromosome Polymorphism. Available online: http://hdl.handle.net/11449/151740 (accessed on 12 March 2018).
- Smit, A.; Hubley, R. RepeatModeler Open-1.0 2015. Available online: http://www.repeatmasker.org/RepeatModeler/ (accessed on 18 December 2015).
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.; Hubley, R.; Green, P. RepeatMasker Open-4.0 2013. Available online: http://www.repeatmasker.org/ (accessed on 5 July 2015).
- Novak, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A Galaxy-based web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef] [PubMed]
- Novak, P.; (Institute of Plant Molecular Biology, České Budějovice, Czech Republic); Macas, J.; (Institute of Plant Molecular Biology, České Budějovice, Czech Republic). Personal Communication, 2014.
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Fastqc 2012. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 January 2014).
- Hannon FASTX-Toolkit 2010. Available online: http://hannonlab.cshl.edu/fastx_toolkit/index.html (accessed on 1 August 2014).
- Staton, E. Pairfq 2014. Available online: https://github.com/sestaton/Pairfq (accessed 15 April 2014).
- Garcia-Alcalde, F.; Okonechnikov, K.; Carbonell, J.; Cruz, L.M.; Gotz, S.; Tarazona, S.; Dopazo, J.; Meyer, T.F.; Conesa, A. Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 2012, 28, 2678–2679. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R. BEDTools: The swiss-army tool for genome feature analysis. In Current Protocols in Bioinformatics; Wiley Online Library: Hoboken, NJ, USA, 2014; ISBN 0471250937. [Google Scholar]
- Marques, D.F. Functional analysis of B chromosome presence using cichlid Astatotilapia latifasciata as model. Available online: http://hdl.handle.net/11449/141953 (accessed on 12 March 2018).
- Criscione, S.W.; Zhang, Y.; Thompson, W.; Sedivy, J.M.; Neretti, N. Transcriptional landscape of repetitive elements in normal and cancer human cells. BMC Genom. 2014, 15, 583. [Google Scholar] [CrossRef] [PubMed]
- Criscione, S. RepEnrich Tutorial. Available online: https://github.com/nerettilab/RepEnrich (accessed on 17 April 2016).
- Gene Link Oligo Explorer 1.2 2005. Available online: http://www.genelink.com/tools/gl-oe.asp (accessed on 14 November 2014).
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.; Takahashi, C.; Moreira-Filho, O. Citotaxonomic consideration on Hoplias lacerdae (Pisces, Erythrinidae). Braz. J. Genet. 1978, 1, 103–120. [Google Scholar]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef] [PubMed]
- Bel, Y.; Ferré, J.; Escriche, B. Quantitative real-time PCR with SYBR Green detection to assess gene duplication in insects: Study of gene dosage in Drosophila melanogaster (Diptera) and in Ostrinia nubilalis (Lepidoptera). BMC Res. Notes 2011, 4, 84. [Google Scholar] [CrossRef] [PubMed]
- Chalopin, D.; Fan, S.; Simakov, O.; Meyer, A.; Schartl, M.; Volff, J.N. Evolutionary active transposable elements in the genome of the coelacanth. J. Exp. Zool. Part B Mol. Dev. Evol. 2014, 322, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.N. Comparative analysis of transposable elements highlights mobilome diversity and evolution in vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Sotero-Caio, C.G.; Platt, R.N.; Suh, A.; Ray, D.A. Evolution and Diversity of Transposable Elements in Vertebrate Genomes. Genome Biol. Evol. 2017, 9, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Brawand, D.; Wagner, C.E.; Li, Y.I.; Malinsky, M.; Keller, I.; Fan, S.; Simakov, O.; Ng, A.Y.; Lim, Z.W.; Bezault, E.; et al. The genomic substrate for adaptive radiation in African cichlid fish. Nature 2014, 513, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belyayev, A. Bursts of transposable elements as an evolutionary driving force. J. Evol. Biol. 2014, 27, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Hedges, D.J.; Deininger, P.L. Inviting instability: Transposable elements, double-strand breaks, and the maintenance of genome integrity. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2007, 616, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.; Jiang, N.; Wessler, S.R. Plant transposable elements: Where genetics meets genomics. Nat. Rev. Genet. 2002, 3, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Klemme, S.; Banaei-Moghaddam, A.M.; Macas, J.; Wicker, T.; Novák, P.; Houben, A. High-copy sequences reveal distinct evolution of the rye B chromosome. New Phytol. 2013, 199, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Kaessmann, H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010, 20, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Maestre, J.; Tchénio, T.; Dhellin, O.; Heidmann, T. mRNA retroposition in human cells: Processed pseudogene formation. EMBO J. 1995, 14, 6333–6338. [Google Scholar] [PubMed]
- Moran, J.V.; DeBerardinis, R.J.; Kazazian, H.H. Exon shuffling by L1 retrotransposition. Science 1999, 283, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.C.; Tingey, S.V.; Morgante, M. Abundance, distribution, and transcriptional activity of repetitive elements in the maize genome. Genome Res. 2001, 11, 1660–1676. [Google Scholar] [CrossRef] [PubMed]
- González, L.G.; Deyholos, M.K. Identification, characterization and distribution of transposable elements in the flax (Linum usitatissimum L.) genome. BMC Genom. 2012, 13, 644. [Google Scholar] [CrossRef] [PubMed]
- Vicient, C.M. Transcriptional activity of transposable elements in maize. BMC Genom. 2010, 11, 601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houben, A. B chromosomes—A matter of chromosome drive. Front. Plant Sci. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Cabrero, J.; Camacho, J.P.M.; López-León, M.D. Gypsy, RTE and Mariner transposable elements populate Eyprepocnemis plorans genome. Genetica 2012, 140, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Bugno-Poniewierska, M.; Solek, P.; Wronski, M.; Potocki, L.; Jezewska-Witkowska, G.; Wnuk, M. Genome organization and DNA methylation patterns of B chromosomes in the red fox and Chinese raccoon dogs. Hereditas 2014, 151, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Ramos, É.; Cardoso, A.L.; Brown, J.; Marques, D.F.; Fantinatti, B.E.A.; Cabral-de-Mello, D.C.; Oliveira, R.A.; O’Neill, R.J.; Martins, C. The repetitive DNA element BncDNA, enriched in the B chromosome of the cichlid fish Astatotilapia latifasciata, transcribes a potentially noncoding RNA. Chromosoma 2017, 126, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; MacManes, M.D. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.D.; Watanabe, C.K. GMAP: A genomic mapping and alignment program for mRNA and EST sequences. Bioinformatics 2005, 21, 1859–1875. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cluster | 0B Reads | 2B Reads | Annotation | Repeat Class |
|---|---|---|---|---|
| 28 | 359 | 23,389 | Bel/Pao | Retro/LTR |
| 63 | 2939 | 13,563 | DNA/hAT | DNA |
| 67 | 351 | 15,603 | Gypsy | Retro/LTR |
| 74 | 4575 | 11,117 | L2 | Retro/LINE |
| Element | Class | Family | F1-0B/M1-0B | F2-1B/M1-0B | M2-1B/M1-0B | M3-1B/M1-0B | M4-2B/M1-0B |
|---|---|---|---|---|---|---|---|
| Gypsy-188_DR-I | LTR | Gypsy | 0.00 | 357.90 | 478.75 | 505.95 | 977.98 |
| AlRepB-26 | LINE | L2 | 2.07 | 454.47 | 438.12 | 454.37 | 976.85 |
| BEL32-I_DR | LTR | Pao | 11.24 | 276.16 | 324.04 | 343.45 | 731.48 |
| Gypsy-23_GA-I | LTR | Gypsy | 0.00 | 311.83 | 336.77 | 360.23 | 709.62 |
| Mariner-N13_DR | DNA | TcMar-Tc1 | 17.81 | 276.37 | 294.65 | 312.57 | 648.31 |
| AlRepB-738 | DNA | hAT-Ac | 23.67 | 320.79 | 356.62 | 382.23 | 631.88 |
| BEL32-LTR_DR | LTR | Pao | 5.46 | 215.99 | 278.12 | 309.23 | 593.38 |
| Maui | LINE | L2 | 777.82 | 916.34 | 626.59 | 665.77 | 312.38 |
| AlRepC-927 | DNA | hAT-Ac | 253.98 | 250.69 | 96.81 | 92.91 | 0.00 |
| AlRepB-157 | Satellite | Satellite | 545.89 | 537.41 | 39.69 | 23.75 | 44.40 |
| TZSAT | Satellite | Satellite | 181.04 | 177.25 | 4.73 | 4.52 | 6.86 |
| Element | Superfamily | Class | Fold-Change | B Expansion |
|---|---|---|---|---|
| Brain | ||||
| AlRepD-1119 | L2 | LINE | 1.4829 | 22/88/115/126/187 |
| CR1-28_HM | CR1 | LINE | 3.2887 | No |
| REX1-3_XT | Rex-Babar | LINE | 1.7297 | No |
| GYPSY2-I_CB | Gypsy | LTR | 5.2102 | No |
| AlRepC-299 | Unknown | Unknown | 1.5748 | 9/16/4/6/0 |
| I-6_AAe | I | LINE | 3.6624 | No |
| L1-6_DR | L1 | LINE | −3.8609 | 2/0/0/0/2 |
| BovBa-1_EF | RTE-BovB | LINE | 4.1560 | No |
| AlRepD-1964 | Unknown | Unknown | 1.1420 | 24/117/121/120/160 |
| Gypsy-34-I_DR | Gypsy | LTR | 4.5448 | No |
| AlRepD-520 | Unknown | Unknown | 1.8113 | 47/30/6/2/0 |
| Mariner-1_SP | TcMar-Fot1 | DNA | 4.0150 | 0/0/2/5/12 |
| ERV1-3N-EC_I-int | ERV1 | LTR | 2.4732 | No |
| Gypsy-17-I_DR | Gypsy | LTR | −1.7877 | 2/2/2/4/16 |
| Gypsy52-I_DR | Gypsy | LTR | 5.4846 | No |
| Tc1-2Eso | TcMar-Tc1 | DNA | 2.4478 | No |
| Rex1-52_DR | Rex-Babar | LINE | −3.1220 | No |
| Helitron-2_DR | Helitron | RC | −1.2385 | No |
| RMER17C-int | ERVK | LTR | 2.6684 | No |
| Gypsy-20-I_DR | Gypsy | LTR | −2.1144 | 0/0/2/0/0 |
| Muscle | ||||
| AlRepC-299 | Unknown | Unknown | 7.5228 | 9/16/4/6/0 |
| AlRepB-358 | hAT-Ac | DNA | 3.1883 | 207/209/146/139/11 |
| Mariner-1_SP | TcMar-Fot1 | DNA | 11.0098 | 0/0/2/5/12 |
| Gonad Male | ||||
| AlRepD-4130 | hAT-Ac | DNA | 2.7431 | 37/87/51/56/31 |
| HEROTn | R2-Hero | LINE | −6.4246 | No |
| 7SLRNA | srpRNA | srpRNA | 3.4512 | No |
| P-27_HM | P | DNA | −2.5097 | No |
| AgaP15 | P | DNA | −5.8314 | No |
| Charlie16 | hAT-Charlie | DNA | −5.7227 | No |
| AlRepE-134 | DNA | DNA | −2.9391 | 6/3/3/5/3 |
| ERV-4_CPB-I | ERV1 | LTR | −2.9007 | No |
| Gypsy-10_GA-LTR | Gypsy | LTR | −2.6009 | No |
| AlRepE-2243 | Unknown | Unknown | 3.6990 | 5/2/2/0/2 |
| CR1-20_CQ | CR1 | LINE | −2.9297 | No |
| Dada-1_ON | Dada | DNA | −3.1755 | No |
| Copia3-I_XT | Copia | LTR | −5.6443 | No |
| AlRepD-3555 | Unknown | Unknown | −4.5855 | No |
| Gonad Female | ||||
| AlRepD-1636 | Unknown | Unknown | 5.5199 | 23/94/96/104/111 |
| hAT-27_LCh | DNA | DNA | 3.0980 | No |
| AlRepD-4141 | Unknown | Unknown | 6.7706 | 2/0/0/0/2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coan, R.L.B.; Martins, C. Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata. Genes 2018, 9, 269. https://doi.org/10.3390/genes9060269
Coan RLB, Martins C. Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata. Genes. 2018; 9(6):269. https://doi.org/10.3390/genes9060269
Chicago/Turabian StyleCoan, Rafael L. B., and Cesar Martins. 2018. "Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata" Genes 9, no. 6: 269. https://doi.org/10.3390/genes9060269
APA StyleCoan, R. L. B., & Martins, C. (2018). Landscape of Transposable Elements Focusing on the B Chromosome of the Cichlid Fish Astatotilapia latifasciata. Genes, 9(6), 269. https://doi.org/10.3390/genes9060269