Prenatal Diagnosis of Fragile X Syndrome in a Twin Pregnancy Complicated by a Complete Retraction

 , ,
, , 
Abstract
:1. Introduction
2. Methods
2.1. Ethics
2.2. Sample Processing
2.3. CGG Sizing
2.4. FMR1 Methylation Analysis
2.5. FMR1 mRNA Analysis
3. Results
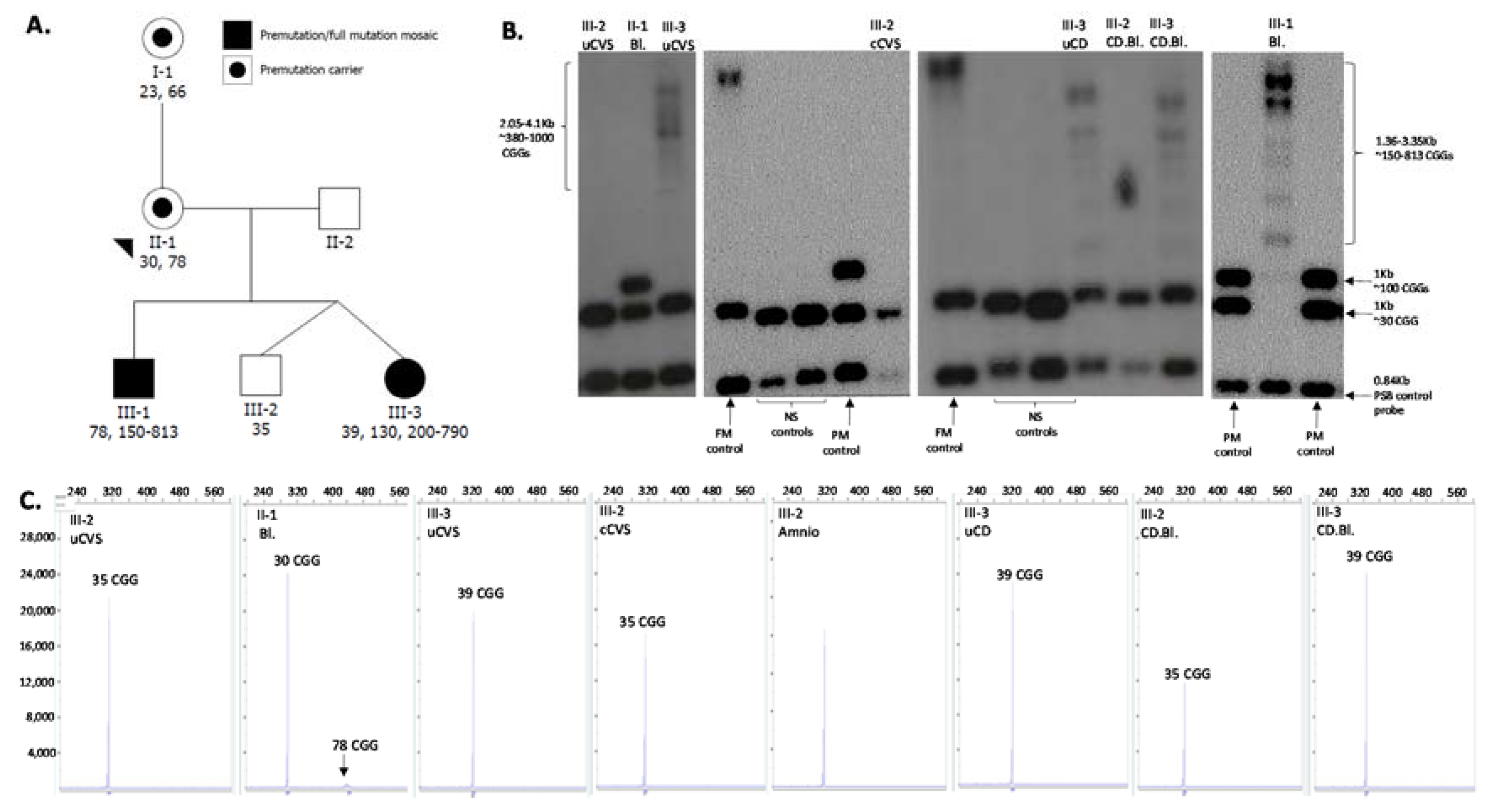
3.1. Clinical History
3.2. Initial Diagnostic Testing
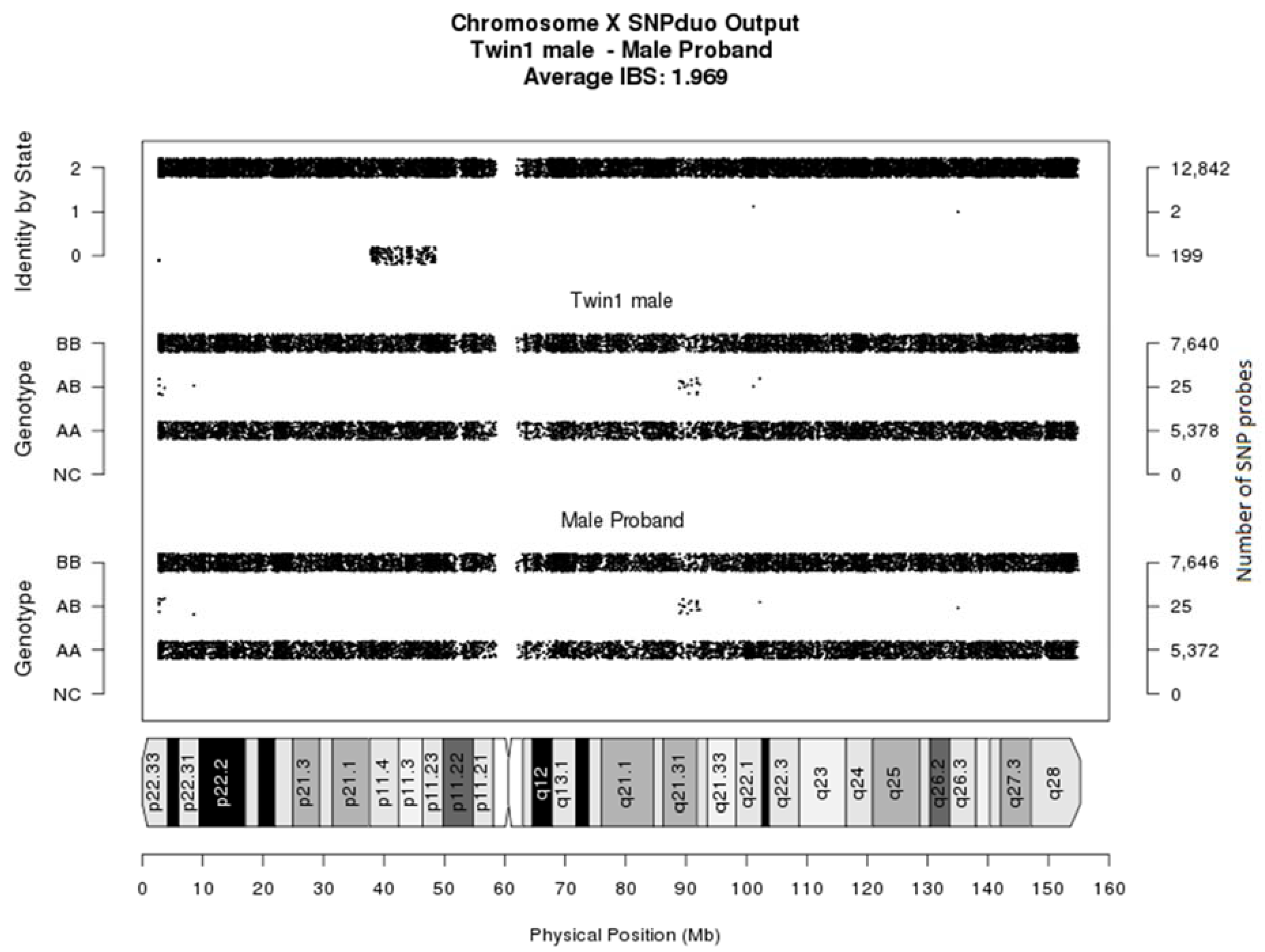
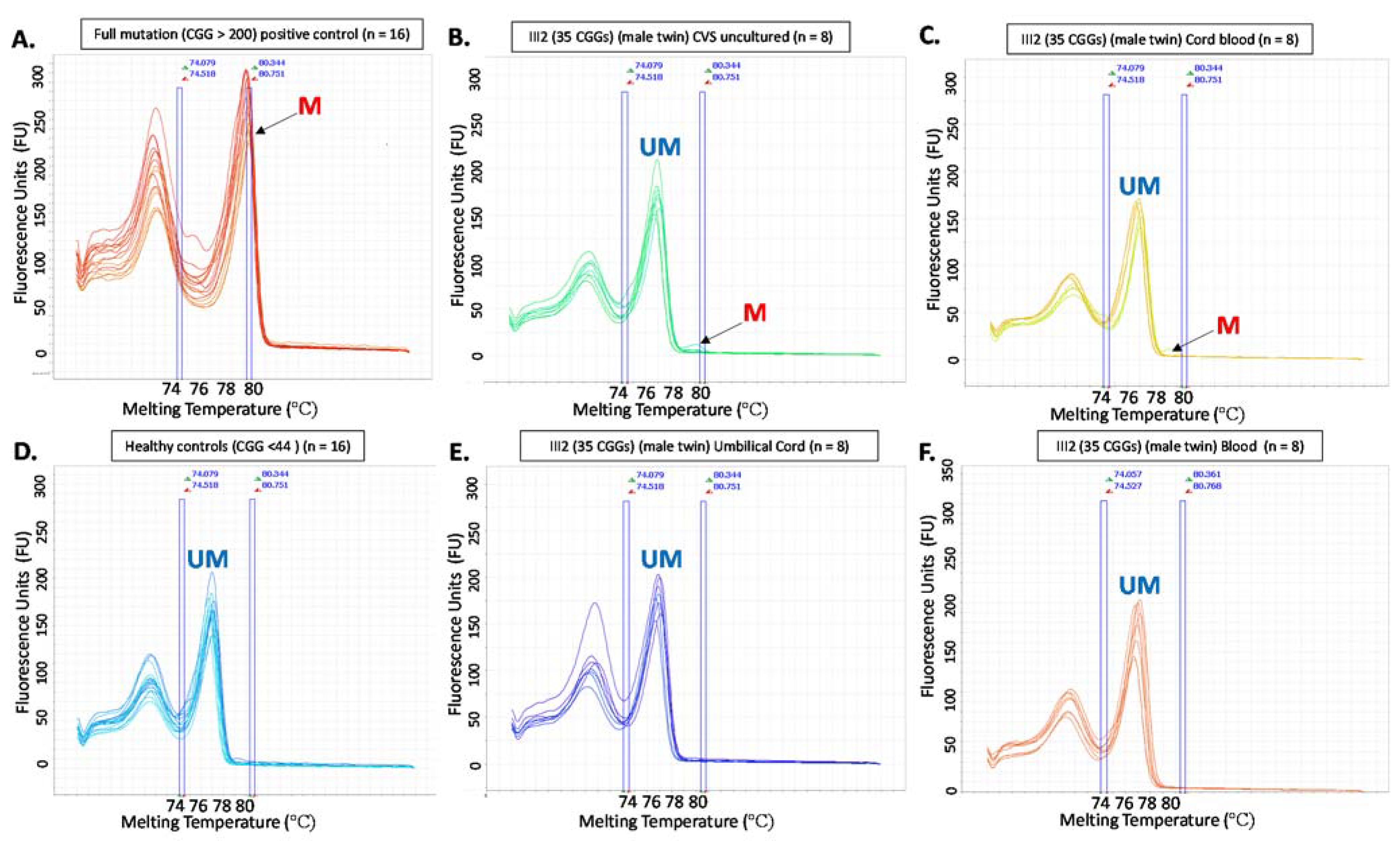
3.3. Follow-Up Prenatal Investigation
3.4. PostNatal Investigation
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hagerman, R.J.; Berry-Kravis, E.; Kaufmann, W.E.; Ono, M.Y.; Tartaglia, N.; Lachiewicz, A.; Kronk, R.; Delahunty, C.; Hessl, D.; Visootsak, J.; et al. Advances in the treatment of fragile X syndrome. Pediatrics 2009, 123, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Godler, D.E.; Tassone, F.; Loesch, D.Z.; Taylor, A.K.; Gehling, F.; Hagerman, R.J.; Burgess, T.; Ganesamoorthy, D.; Hennerich, D.; Gordon, L.; et al. Methylation of novel markers of fragile X alleles is inversely correlated with FMRP expression and FMR1 activation ratio. Hum. Mol. Genet. 2010, 19, 1618–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddalena, A.; Richards, C.S.; McGinniss, M.J.; Brothman, A.; Desnick, R.J.; Grier, R.E.; Hirsch, B.; Jacky, P.; McDowell, G.A.; Popovich, B.; et al. Technical standards and guidelines for fragile X: The first of a series of disease-specific supplements to the standards and guidelines for clinical genetics laboratories of the American college of medical genetics. Quality assurance subcommittee of the laboratory practice committee. Genet. Med. 2001, 3, 200–205. [Google Scholar] [PubMed]
- Rodriguez-Revenga, L.; Madrigal, I.; Pagonabarraga, J.; Xuncla, M.; Badenas, C.; Kulisevsky, J.; Gomez, B.; Mila, M. Penetrance of FMR1 premutation associated pathologies in fragile X syndrome families. Eur. J. Hum. Genet. 2009, 17, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Warren, S.T. Understanding the molecular basis of fragile X syndrome. Hum. Mol. Genet. 2000, 9, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolin, S.L.; Brown, W.T.; Glicksman, A.; Houck, G.E., Jr.; Gargano, A.D.; Sullivan, A.; Biancalana, V.; Brondum-Nielsen, K.; Hjalgrim, H.; Holinski-Feder, E.; et al. Expansion of the fragile X CGG repeat in females with premutation or intermediate alleles. Am. J. Hum. Genet. 2003, 72, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Biancalana, V.; Glaeser, D.; McQuaid, S.; Steinbach, P. EMQN best practice guidelines for the molecular genetic testing and reporting of fragile x syndrome and other fragile X-associated disorders. Eur. J. Hum. Genet. 2015, 23, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Khaniani, M.S.; Kalitsis, P.; Burgess, T.; Slater, H.R. An improved diagnostic PCR assay for identification of cryptic heterozygosity for CGG triplet repeat alleles in the fragile X gene (FMR1). Mol. Cytogenet. 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Francis, D.; Burgess, T.; Mitchell, J.; Slater, H. Identification of small FRAXA premutations. Mol. Diagn. 2000, 5, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Godler, D.E.; Slater, H.R.; Bui, Q.M.; Storey, E.; Ono, M.Y.; Gehling, F.; Inaba, Y.; Francis, D.; Hopper, J.L.; Kinsella, G.; et al. Fragile X mental retardation 1 (FMR1) intron 1 methylation in blood predicts verbal cognitive impairment in female carriers of expanded FMR1 alleles: Evidence from a pilot study. Clin. Chem. 2012, 58, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Inaba, Y.; Schwartz, C.E.; Bui, Q.M.; Li, X.; Skinner, C.; Field, M.; Wotton, T.; Hagerman, R.J.; Francis, D.; Amor, D.J.; et al. Early detection of fragile X syndrome: Applications of a novel approach for improved quantitative methylation analysis in venous blood and newborn blood spots. Clin. Chem. 2014, 60, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Loesch, D.Z.; Godler, D.E.; Evans, A.; Bui, Q.M.; Gehling, F.; Kotschet, K.E.; Trost, N.; Storey, E.; Stimpson, P.; Kinsella, G.; et al. Evidence for the toxicity of bidirectional transcripts and mitochondrial dysfunction in blood associated with small CGG expansions in the FMR1 gene in patients with parkinsonism. Genet. Med. 2011, 13, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Kraan, C.M.; Cornish, K.M.; Bui, Q.M.; Li, X.; Slater, H.R.; Godler, D.E. β-glucuronidase use as a single internal control gene may confound analysis in FMR1 mRNA toxicity studies. PLoS ONE 2018, 13, e0192151. [Google Scholar] [CrossRef] [PubMed]
- Arpone, M.; Baker, E.K.; Bretherton, L.; Bui, M.; Li, X.; Whitaker, S.; Dissanayake, C.; Cohen, J.; Hickerton, C.; Rogers, C.; et al. Intragenic DNA methylation in buccal epithelial cells and intellectual functioning in a paediatric cohort of males with fragile X. Sci. Rep. 2018, 8, 3644. [Google Scholar] [CrossRef] [PubMed]
- Nolin, S.L.; Glicksman, A.; Houck, G.E., Jr.; Brown, W.T.; Dobkin, C.S. Mosaicism in fragile X affected males. Am. J. Med. Genet. 1994, 51, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Aliaga, S.M.; Slater, H.R.; Francis, D.; Du Sart, D.; Li, X.; Amor, D.J.; Alliende, A.M.; Santa Maria, L.; Faundes, V.; Morales, P.; et al. Identification of males with cryptic fragile X alleles by methylation-specific quantitative melt analysis. Clin. Chem. 2016, 62, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Manor, E.; Jabareen, A.; Magal, N.; Kofman, A.; Hagerman, R.J.; Tassone, F. Prenatal diagnosis of fragile X: Can a full mutation allele in the FMR1 gene contract to a normal size? Front. Genet. 2017, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, M.P.; Cohen, M.; Vnencak-Jones, C.L. Maternal FMR1 premutation allele expansion and contraction in fraternal twins. Am. J. Med. Genet. A 2013, 161, 2620–2625. [Google Scholar]
- Ferreira, S.I.; Pires, L.M.; Ferrao, J.; Sa, J.; Serra, A.; Carreira, I.M. Mosaicism for FMR1 gene full mutation and intermediate allele in a female foetus: A postzygotic retraction event. Gene 2013, 527, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Stark, Z.; Francis, D.; Gaffney, L.; Greenberg, J.; Hills, L.; Li, X.; Godler, D.E.; Slater, H.R. Prenatal diagnosis of fragile X syndrome complicated by full mutation retraction. Am. J. Med. Genet. A 2015, 167, 2485–2487. [Google Scholar] [CrossRef] [PubMed]
- Grasso, M.; Faravelli, F.; Lo Nigro, C.; Chiurazzi, P.; Sperandeo, M.P.; Argusti, A.; Pomponi, M.G.; Lecora, M.; Sebastio, G.F.; Perroni, L.; et al. Mosaicism for the full mutation and a microdeletion involving the CGG repeat and flanking sequences in the FMR1 gene in eight fragile X patients. Am. J. Med. Genet. 1999, 85, 311–316. [Google Scholar] [CrossRef]
- Gerhardt, J.; Zaninovic, N.; Zhan, Q.; Madireddy, A.; Nolin, S.L.; Ersalesi, N.; Yan, Z.; Rosenwaks, Z.; Schildkraut, C.L. Cis-acting DNA sequence at a replication origin promotes repeat expansion to fragile X full mutation. J. Cell Biol. 2014, 206, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.T.; Aliaga, S.M.; Arpone, M.; Francis, D.; Li, X.; Chong, B.; Slater, H.R.; Rogers, C.; Bretherton, L.; Hunter, M.; et al. Partially methylated alleles, microdeletion, and tissue mosaicism in a fragile X male with tremor and ataxia at 30 years of age: A case report. Am. J. Med. Genet. A 2016, 170, 3327–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| ID | Sample type | CGG size | MS-QMA (MR) | EpiTYPER (MR) CpG10-12 FREE2 | FMR1 mRNA |
|---|---|---|---|---|---|
| III-1 (older brother) | Blood | 78; 150–813 | 0.55 (<0.03; n = 23) | 0.58 (<0.07; n = 17) | 0.98# |
| III-1 (older brother) | Buccal | 78; 150–813 | 0.54 (<0.03; n = 23) | 0.69 (<0.07; n = 17) | - |
| II-1 (PM mother) | Blood | 30; 70 | 0.21 (<0.37; n = 88) | - | - |
| II-1 (PM mother) | Buccal | 30; 70 | 0.19 (<0.36; n = 26) | - | - |
| II-1 (PM mother) | Saliva | 30; 70 | 0.2 (<0.4; n = 33) | - | - |
| III-2 (male twin) | Uncultured CVS | 35 | 0.01* (<0.03; n = 7) | - | - |
| III-2 (male twin) | Amnio | 35 | - | - | - |
| III-2 (male twin) | Cord blood | 35 | 0.01*(<0.03; n = 23) | - | - |
| III-2 (male twin) | Umbilical cord | 35 | 0.01 | - | - |
| III-2 (male twin) | Blood | 35 | 0.01 (<0.03; n = 23) | - | 1.16# |
| III-3 (female twin) | Uncultured CVS | 39; 380–1000 | 0.269 | - | - |
| III-3 (female twin) | Umbilical cord | 39; 130; 200–790 | - | 0.35 ( <0.43; n = 154)** | - |
| III-3 (female twin) | Cord blood | 39; 130; 200–790 | 0.27 (<0.37; n = 88) | 0.3 (<0.435; n = 154)** | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prawer, Y.; Hunter, M.; Cronin, S.; Ling, L.; Aliaga Vera, S.; Fahey, M.; Gelfand, N.; Oertel, R.; Bartlett, E.; Francis, D.; et al. Prenatal Diagnosis of Fragile X Syndrome in a Twin Pregnancy Complicated by a Complete Retraction. Genes 2018, 9, 287. https://doi.org/10.3390/genes9060287
Prawer Y, Hunter M, Cronin S, Ling L, Aliaga Vera S, Fahey M, Gelfand N, Oertel R, Bartlett E, Francis D, et al. Prenatal Diagnosis of Fragile X Syndrome in a Twin Pregnancy Complicated by a Complete Retraction. Genes. 2018; 9(6):287. https://doi.org/10.3390/genes9060287
Chicago/Turabian StylePrawer, Yael, Matthew Hunter, Sara Cronin, Ling Ling, Solange Aliaga Vera, Michael Fahey, Nikki Gelfand, Ralph Oertel, Essra Bartlett, David Francis, and et al. 2018. "Prenatal Diagnosis of Fragile X Syndrome in a Twin Pregnancy Complicated by a Complete Retraction" Genes 9, no. 6: 287. https://doi.org/10.3390/genes9060287
APA StylePrawer, Y., Hunter, M., Cronin, S., Ling, L., Aliaga Vera, S., Fahey, M., Gelfand, N., Oertel, R., Bartlett, E., Francis, D., & Godler, D. (2018). Prenatal Diagnosis of Fragile X Syndrome in a Twin Pregnancy Complicated by a Complete Retraction. Genes, 9(6), 287. https://doi.org/10.3390/genes9060287