C/EBPβ Promotes STAT3 Expression and Affects Cell Apoptosis and Proliferation in Porcine Ovarian Granulosa Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
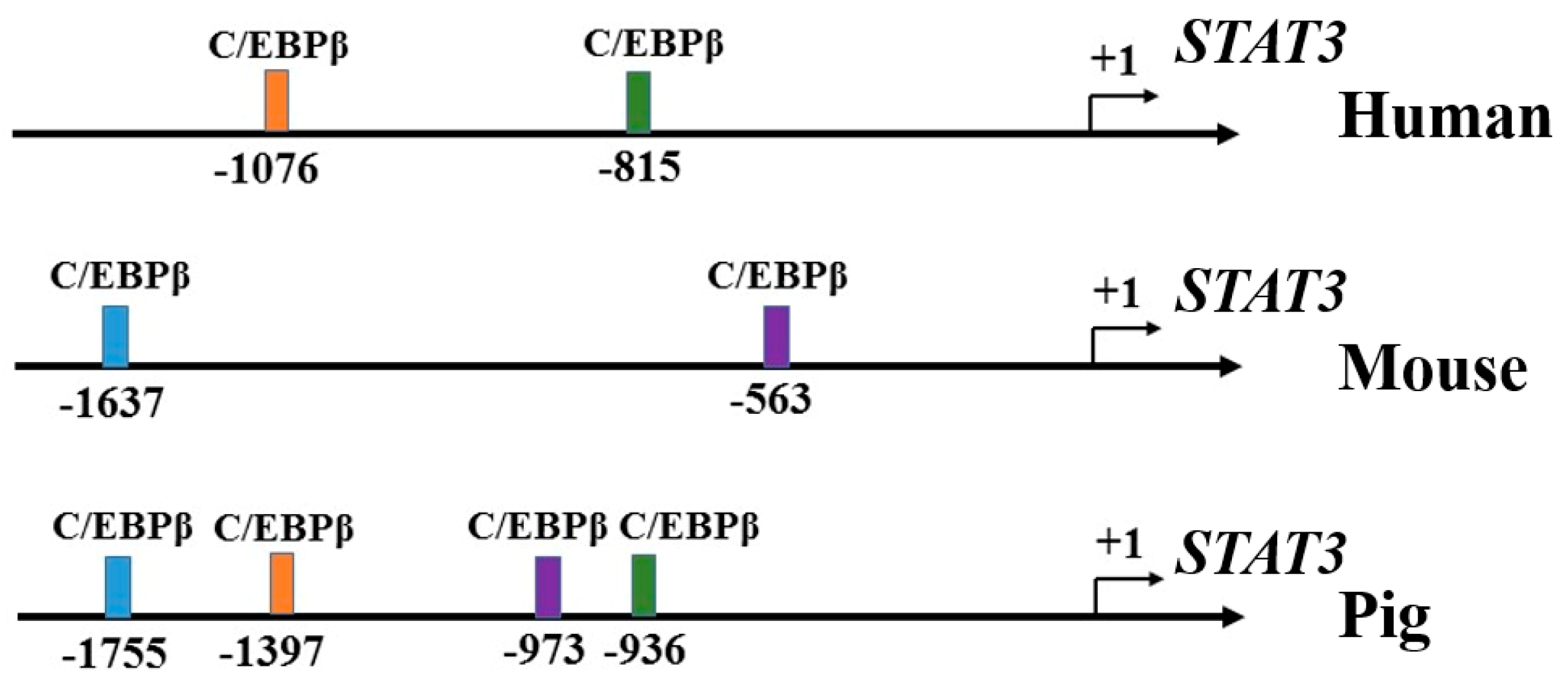
2.2. Prediction of Putative C/EBPβ Binding Sites on the Promoter of STAT3
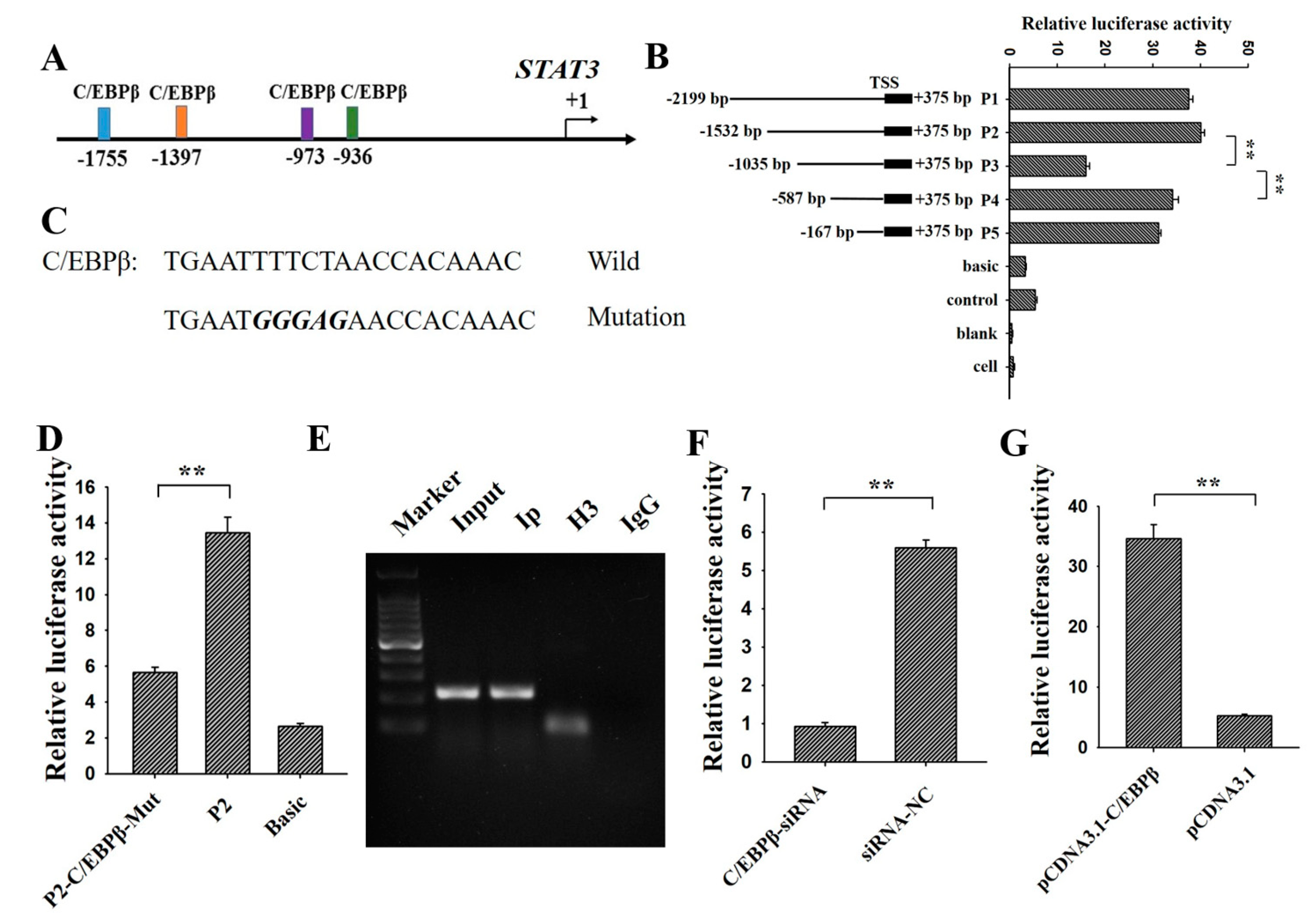
2.3. Construction of STAT3 5′ Deletion and Luciferase Assay
2.4. Culture of Porcine GCs In Vitro
2.5. Chromatin Immunoprecipitation Assay
2.6. Real-Time Quantitative PCR Analysis
2.7. Cell Proliferation and Apoptosis Assay
2.8. Western Blot Analysis
2.9. Data Analysis
3. Results
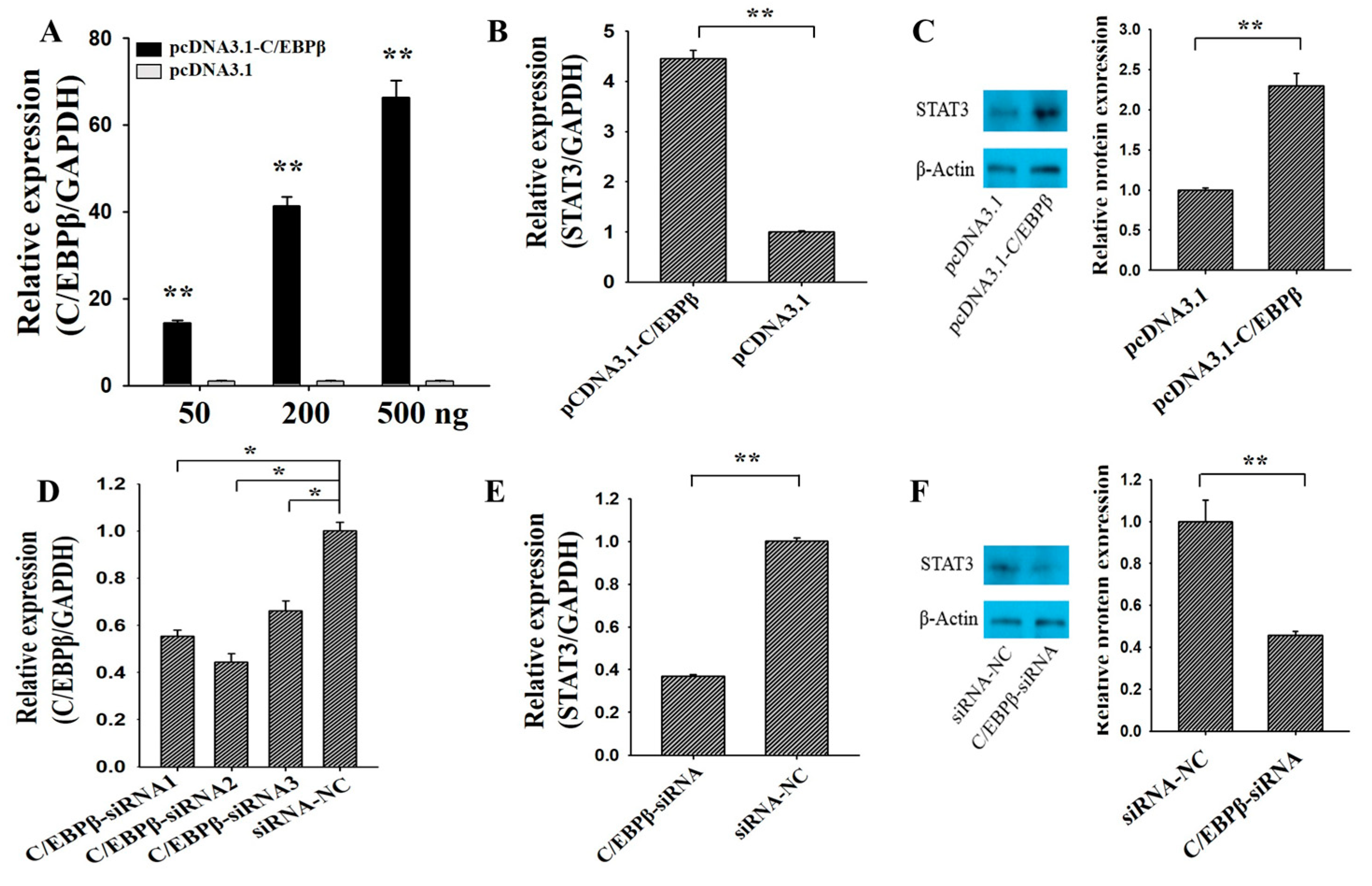
3.1. C/EBPβ Promotes the mRNA and Protein Level of STAT3
3.2. C/EBPβ Binding at −1397/−1387 Region of STAT3
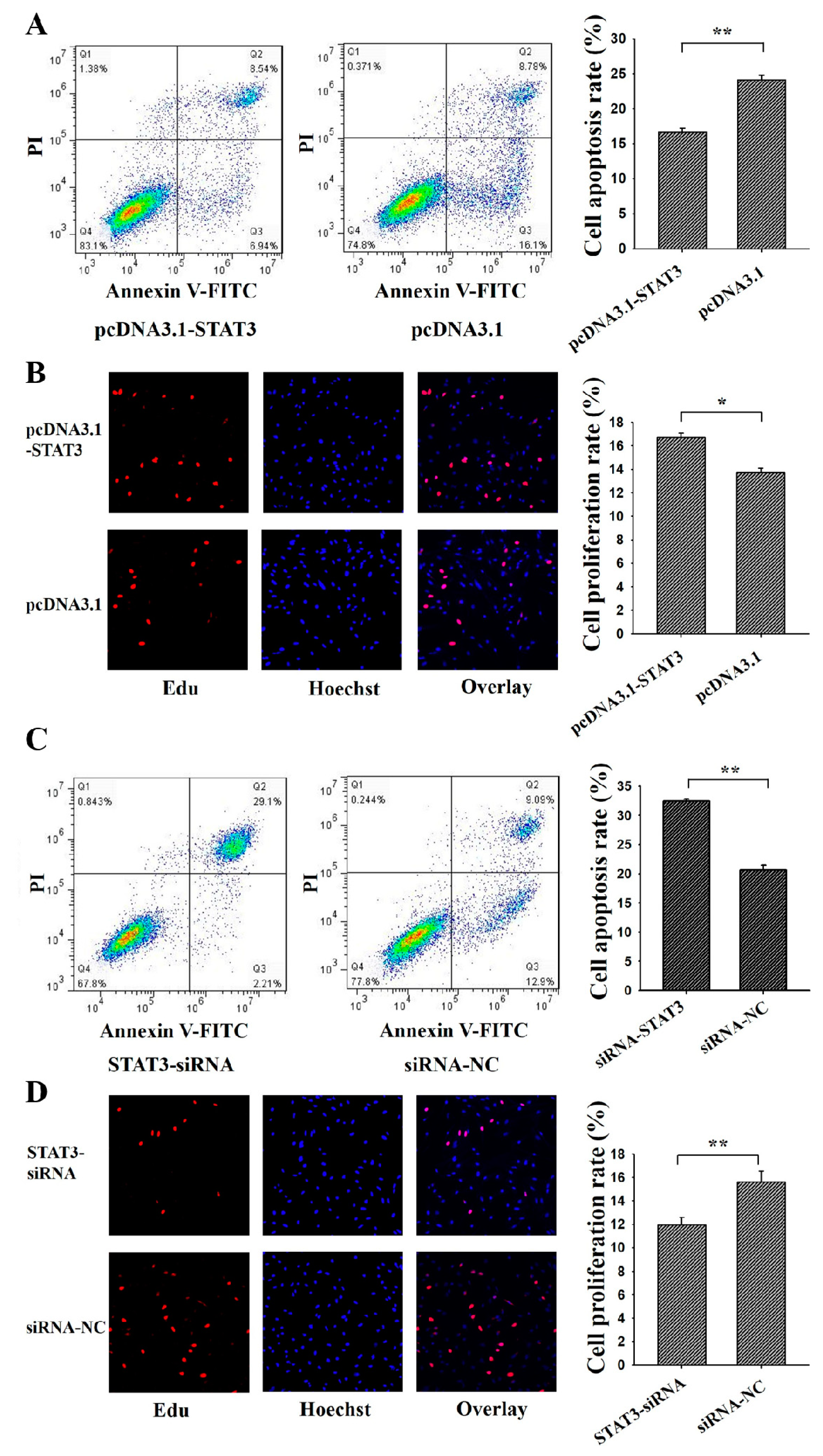
3.3. STAT3 Inhibits Apoptosis and Promotes Proliferation of Ovarian GCs
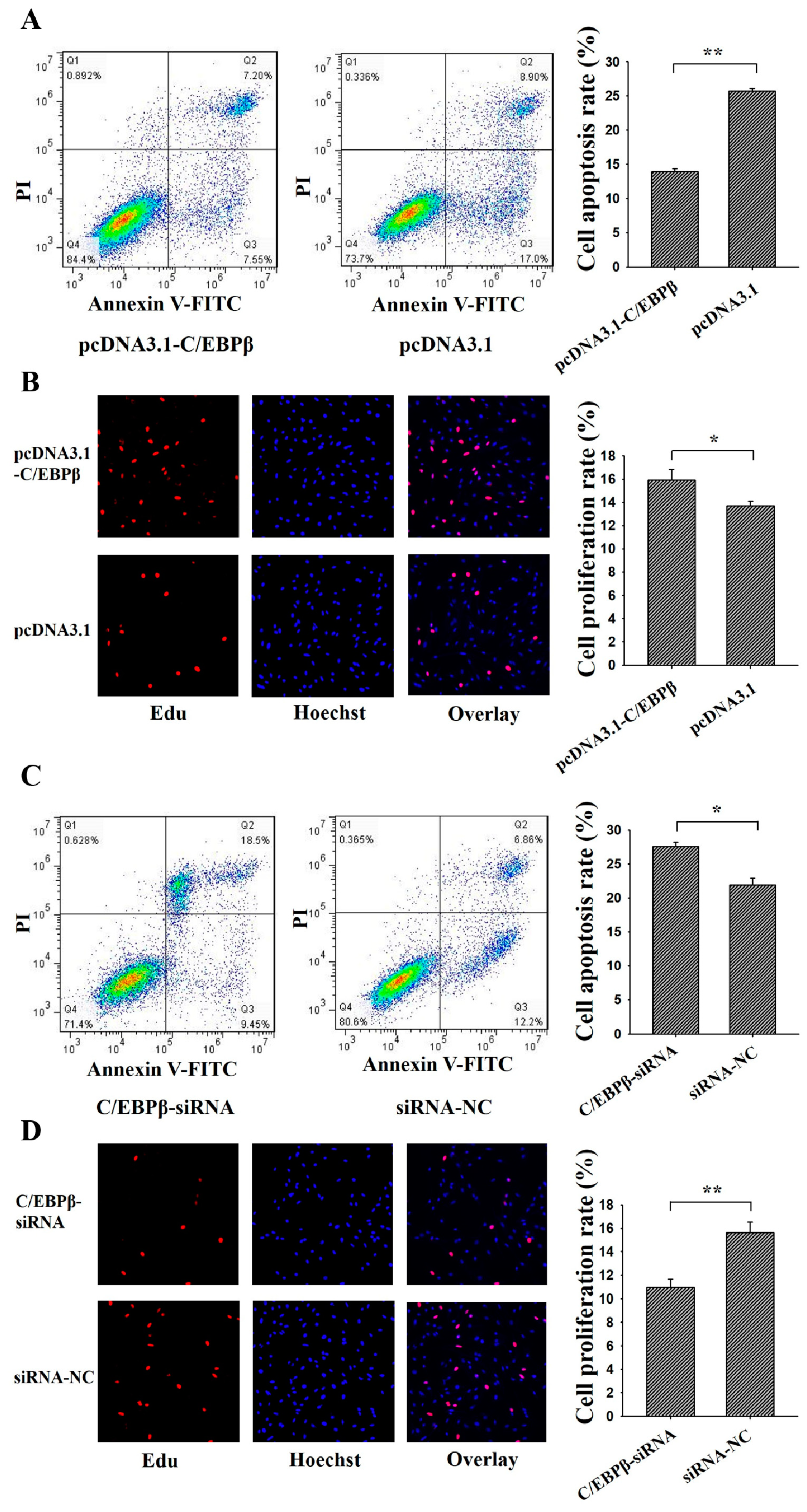
3.4. C/EBPβ Inhibits Apoptosis and Promotes Proliferation of Ovarian GCs
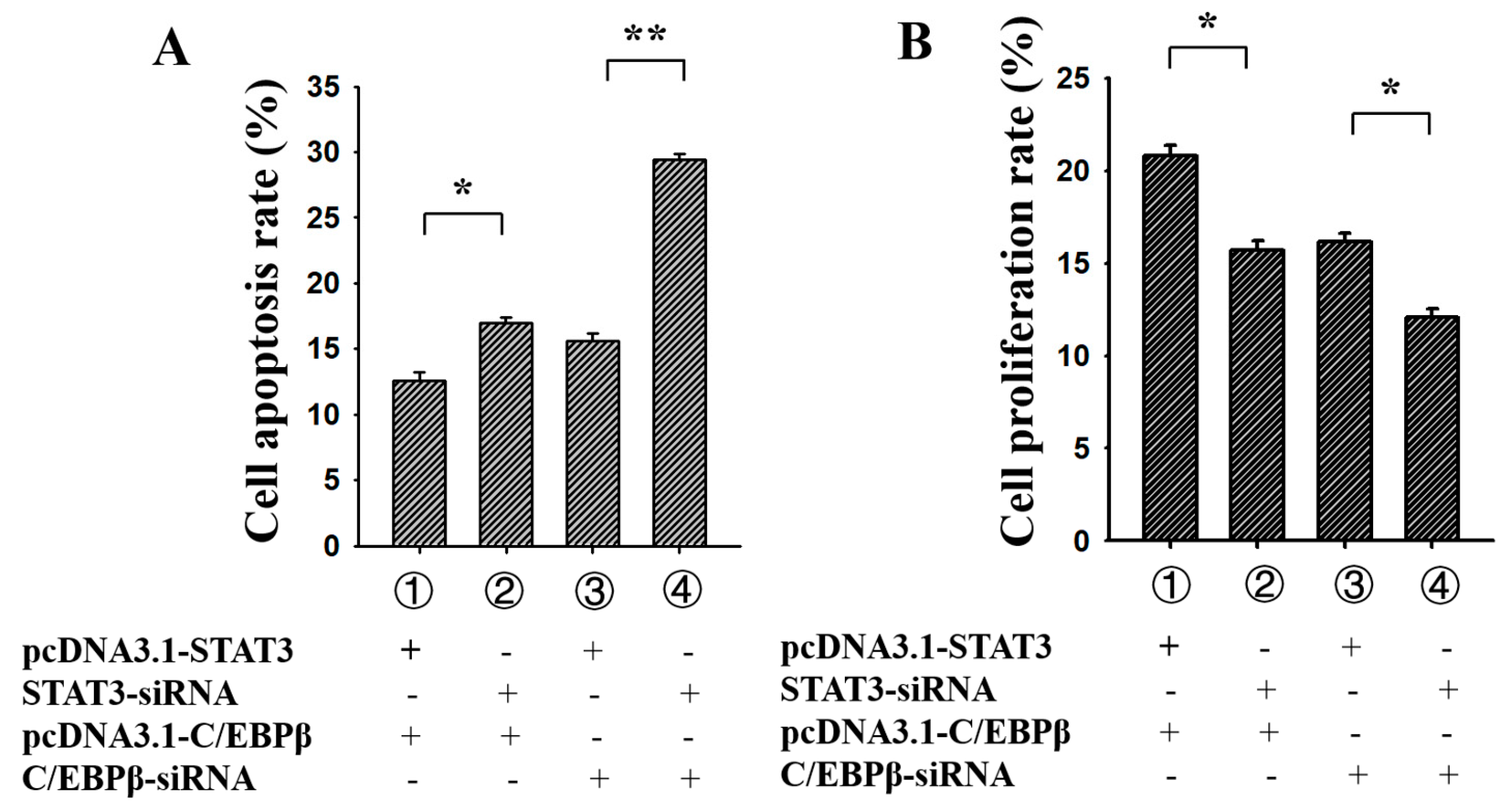
3.5. C/EBPβ Enhanced the Antiapoptotic and Pro-Proliferation Effects of STAT3 in Ovarian GCs
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shirasawa, H.; Terada, Y. In vitro maturation of human immature oocytes for fertility preservation and research material. Reprod. Med. Biol. 2017, 16, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, M.; Prasad, S.; Tripathi, A.; Pandey, A.N.; Ali, I.; Singh, A.K.; Shrivastav, T.G.; Chaube, S.K. Apoptosis in mammalian oocytes: A review. Apoptosis 2015, 20, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Soder, O. Ontogeny of gonadal sex steroids. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.M.; Jones, P.L.; Ingram, B.A. Age-related changes in ovarian characteristics, plasma sex steroids and fertility during pubertal development in captive female murray cod Maccullochella peelii peelii. Comp. Biochem. Phys. 2008, 150, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.A.; Ewen, M.; Mackie, K.; Staines, M.E.; McEvoy, T.G.; Sinclair, K.D. Effect of ammonium chloride on the growth and metabolism of bovine ovarian granulosa cells and the development of ovine oocytes matured in the presence of bovine granulosa cells previously exposed to ammonium chloride. Anim. Reprod. Sci. 2004, 84, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Kubo, N.; Cayo-Colca, I.S.; Miyano, T. Effect of estradiol-17β during in vitro growth culture on the growth, maturation, cumulus expansion and development of porcine oocytes from early antral follicles. Anim. Sci. J. 2015, 86, 251–259. [Google Scholar] [CrossRef] [PubMed]
- McGee, E.A.; Hsueh, A.J. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.G. Oocyte maturation and ovum quality in pigs. Rev. Reprod. 2000, 5, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cesaro, M.P.; Dos Santos, J.T.; Ferst, J.G.; Nobrega, J.E., Jr.; Rosa, P.; Rovani, M.T.; Ilha, G.F.; Bohrer, R.C.; Ferreira, R.; Gasperin, B.G.; et al. Natriuretic peptide system regulation in granulosa cells during follicle deviation and ovulation in cattle. Reprod. Domest. Anim. 2018, 53, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, M.T.; Mlotkowska, P.; Tanski, D.; Lepiarczyk, E.; Oklinski, M.K.; Nielsen, S.; Skowronska, A. Pituitary gonadotropins, prolactin and growth hormone differentially regulate AQP1 expression in the porcine ovarian follicular cells. Int. J. Mol. Sci. 2018, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular growth and atresia in mammalian ovaries: Regulation by survival and death of granulosa cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Hamm, M.L.; Bhat, G.K.; Thompson, W.E.; Mann, D.R. Folliculogenesis is impaired and granulosa cell apoptosis is increased in leptin-deficient mice. Biol. Reprod. 2004, 71, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.E.; Upton, R.M.O.; McLaughlin, E.A.; Sutherland, J.M. Phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) and Janus kinase/signal transducer and activator of transcription (JAK/STAT) follicular signalling is conserved in the mare ovary. Reprod. Fertil. Dev. 2018, 30, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Nakajima, T.; Hayashi, S.; Chambon, P.; Watanabe, H.; Iguchi, T.; Sato, T. Effects of diethylstilbestrol on programmed oocyte death and induction of polyovular follicles in neonatal mouse ovaries. Biol. Reprod. 2009, 81, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Craig, J.; Dyce, P.W.; Li, J. Cloning of porcine signal transducer and activator of transcription 3 cDNA and its expression in reproductive tissues. Reproduction 2006, 132, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, A.G.; Bahr, J.M. Role of FSH and epidermal growth factor (EGF) in the initiation of steroidogenesis in granulosa cells associated with follicular selection in chicken ovaries. Reproduction 2003, 125, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, D.; Zhang, S.; Xing, Y.; Gao, Y.; Wu, J. MicroRNA-125a-5p induces mouse granulosa cell apoptosis by targeting signal transducer and activator of transcription 3. Menopause 2016, 23, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Robker, R.L.; Watson, L.N.; Robertson, S.A.; Dunning, K.R.; McLaughlin, E.A.; Russell, D.L. Identification of sites of STAT3 action in the female reproductive tract through conditional gene deletion. PLoS ONE 2014, 9, e101182. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Liu, Z.; Shimada, M.; Sterneck, E.; Johnson, P.F.; Hedrick, S.M.; Richards, J.S. MAPK3/1 (ERK1/2) in ovarian granulosa cells are essential for female fertility. Science 2009, 324, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Sterneck, E.; Tessarollo, L.; Johnson, P.F. An essential role for C/EBPbeta in female reproduction. Genes Dev. 1997, 11, 2153–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirois, J.; Richards, J.S. Transcriptional regulation of the rat prostaglandin endoperoxide synthase 2 gene in granulosa cells. Evidence for the role of a cis-acting C/EBP beta promoter element. J. Biol. Chem. 1993, 268, 21931–21938. [Google Scholar] [PubMed]
- Signal Transducer and Activator of Transcription 3 Homo sapiens (Human). Available online: https://www.ncbi.nlm.nih.gov/gene/6774 (accessed on 28 May 2018).
- Signal Transducer and Activator of Transcription 3Mus musculus (House Mouse). Available online: https://www.ncbi.nlm.nih.gov/gene/20848 (accessed on 28 May 2018).
- Signal Transducer and Activator of Transcription 3 Sus scrofa (Pig). Available online: https://www.ncbi.nlm.nih.gov/gene/733648 (accessed on 28 May 2018).
- Tsunoda, T.; Takagi, T. Estimating transcription factor bindability on DNA. Bioinformatics 1999, 15, 622–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biobase. Available online: http://gene-regulation.com/pub/programs/alibaba2/index.html (accessed on 28 May 2018).
- Mathelier, A.; Fornes, O.; Arenillas, D.J.; Chen, C.Y.; Denay, G.; Lee, J.; Shi, W.; Shyr, C.; Tan, G.; Worsley-Hunt, R.; et al. JASPAR 2016: A major expansion and update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2016, 44, D110–D115. [Google Scholar] [CrossRef] [PubMed]
- Research. Available online: http://alggen.lsi.upc.es/cgi-bin/promo_v3/promo/promoinit.cgi?dirDB=TF_8.3 (accessed on 28 May 2018).
- Lai, T.C.; Li, H.F.; Li, Y.S.; Hung, P.Y.; Shyu, M.K.; Hu, M.C. Proximal GATA-binding sites are essential for human HSD3B1 gene transcription in the placenta. Sci. Rep. 2017, 7, 4271. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Du, X.; Zhou, J.; Pan, Z.; Liu, H.; Li, Q. MicroRNA-26b functions as a proapoptotic factor in porcine follicular Granulosa cells by targeting Sma-and Mad-related protein 4. Biol. Reprod. 2014, 91, 146. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.J.; Feng, L.J.; Wang, X.; Du, J.; Chen, Y.; Yang, W.; Zhou, C.Y.; Cheng, L.; Shen, Y.J.; Fang, S.Y.; et al. Mesencephalic astrocyte-derived neurotrophic factor is involved in inflammation by negatively regulating the NF-kB pathway. Sci. Rep. 2015, 5, 8133. [Google Scholar] [CrossRef] [PubMed]
- Silverman, E.; Eimerl, S.; Orly, J. CCAAT enhancer-binding protein beta and GATA-4 binding regions within the promoter of the steroidogenic acute regulatory protein (StAR) gene are required for transcription in rat ovarian cells. J. Biol. Chem. 1999, 274, 17987–17996. [Google Scholar] [CrossRef] [PubMed]
- Li, M.G.; Ding, G.L.; Chen, X.J.; Lu, X.P.; Dong, L.J.; Dong, M.Y.; Yang, X.F.; Lu, X.E.; Huang, H.F. Association of serum and follicular fluid leptin concentrations with granulosa cell phosphorylated signal transducer and activator of transcription 3 expression in fertile patients with polycystic ovarian syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 4771–4776. [Google Scholar] [CrossRef] [PubMed]
- Imai, F.; Kishi, H.; Nakao, K.; Nishimura, T.; Minegishi, T. IL-6 up-regulates the expression of rat LH receptors during granulosa cell differentiation. Endocrinology 2014, 155, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Ndiaye, K.; Castonguay, A.; Benoit, G.; Silversides, D.W.; Lussier, J.G. Differential regulation of Janus kinase 3 (JAK3) in bovine preovulatory follicles and identification of JAK3 interacting proteins in granulosa cells. J. Ovarian Res. 2016, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Gasperin, B.G.; Rovani, M.T.; Ferreira, R.; Ilha, G.F.; Bordignon, V.; Goncalves, P.B.; Duggavathi, R. Functional status of STAT3 and MAPK3/1 signaling pathways in granulosa cells during bovine follicular deviation. Theriogenology 2015, 83, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Ilha, G.F.; Rovani, M.T.; Gasperin, B.G.; Antoniazzi, A.Q.; Goncalves, P.B.; Bordignon, V.; Duggavathi, R. Lack of FSH support enhances LIF-STAT3 signaling in granulosa cells of atretic follicles in cattle. Reproduction 2015, 150, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Kou, X.; Zhang, Y.; Zhang, X.; Cheng, G.; Jia, T. Leptin siRNA promotes ovarian granulosa cell apoptosis and affects steroidogenesis by increasing NPY2 receptor expression. Gene 2017, 633, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Liu, Z.L.; Johnson, P.F.; Richards, J.S. CCAAT/enhancer-binding proteins (C/EBP)-alpha and -beta are essential for ovulation, luteinization, and the expression of key target genes. Mol. Endocrinol. 2011, 25, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.A.; Liu, Z.L.; Mullany, L.K.; Fan, C.M.; Richards, J.S. Growth arrest specific-1 (GAS1) is a C/EBP target gene that functions in ovulation and corpus luteum formation in mice. Biol. Reprod. 2016, 94. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Ju, Y.F.; Imamichi, Y.; Osaki, T.; Yazawa, T.; Kawabe, S.; Ishikane, S.; Matsumura, T.; Kanno, M.; Kamiki, Y.; et al. C/EBPβ (CCAAT/enhancer-binding protein β) mediates progesterone production through transcriptional regulation in co-operation with SF-1 (steroidogenic factor-1). Biochem. J. 2014, 460, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Lind, A.K.; Dahm-Kahler, P.; Brannstrom, M.; Carletti, M.Z.; Christenson, L.K.; Curry, T.E., Jr.; Jo, M. RUNX2 transcription factor regulates gene expression in luteinizing granulosa cells of rat ovaries. Mol. Endocrinol. 2010, 24, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Son, D.S.; Terranova, P.F.; Roby, K.F. Interaction of adenosine 3′,5′-cyclic monophosphate and tumor necrosis factor-alpha on serum amyloid A3 expression in mouse granulosa cells: dependence on CCAAT-enhancing binding protein-beta isoform. Endocrinology 2010, 151, 3407–3419. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.K.; Johnson, P.F.; McAllister, J.M.; Strauss, J.F. CCAAT/enhancer-binding proteins regulate expression of the human steroidogenic acute regulatory protein (StAR) gene. J. Biol. Chem. 1999, 274, 26591–26598. [Google Scholar] [CrossRef] [PubMed]
- LaVoie, H.A.; Singh, D.; Hui, Y.Y. Concerted regulation of the porcine steroidogenic acute regulatory protein gene promoter activity by follicle-stimulating hormone and insulin-like growth factor I in granulosa cells involves GATA-4 and CCAAT/enhancer binding protein beta. Endocrinology 2004, 145, 3122–3134. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence * | Product (bp) | Accession Number |
|---|---|---|---|
| P1 (−2199/+375) | F: CGACGCGT TCCTCAACCCACCAAGAAAG | 2575 | NM_001044580 |
| R: CCCTCGAG CTCCCGGTCTCTTCGTATCC | |||
| P2 (−1532/+375) | F: CGACGCGT CTCCAAGTCATTGATTTTCT | 1908 | NM_001044580 |
| R: ditto | |||
| P3 (−1035/+375) | F: CGACGCGT TACTAAACAAACACAATAAA | 1410 | NM_001044580 |
| R: ditto | |||
| P4 (−587/+375) | F: CGACGCGT CTGAGGTTCAAAGCAGGCGG | 963 | NM_001044580 |
| R: ditto | |||
| P5 (−167/+375) | F: CGACGCGT CTCTCCTCATTGGCAAGTGG | 543 | NM_001044580 |
| R: ditto | |||
| qRT-PCR-STAT3 | F: GGGCTTTATCAGTAAGGAGA R: GGAATGTCAGGGTAGAGGTA | 276 | NM_001044580 |
| qRT-PCR -C/EBPβ | F: CGGACTGCAAGCGGAAGGAGGA R: GGCTGGACGACGAGGATGTGGA | 153 | NM_001199889 |
| qRT-PCR-GAPDH | F: TCCCGCCAACATCAAAT R: CACGCCCATCACAAACAT | 201 | NM_001206359 |
| ChIP-STAT3 | F: ATAGCTATCCTTGGGGAGG R: AAGGGCCTGTTATCTCAC | 150 | NM_001044580 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, X.; Zhou, X.; He, Y.; Zhong, Y.; Zhang, A.; Zhang, Z.; Zhang, H.; Li, J. C/EBPβ Promotes STAT3 Expression and Affects Cell Apoptosis and Proliferation in Porcine Ovarian Granulosa Cells. Genes 2018, 9, 295. https://doi.org/10.3390/genes9060295
Yuan X, Zhou X, He Y, Zhong Y, Zhang A, Zhang Z, Zhang H, Li J. C/EBPβ Promotes STAT3 Expression and Affects Cell Apoptosis and Proliferation in Porcine Ovarian Granulosa Cells. Genes. 2018; 9(6):295. https://doi.org/10.3390/genes9060295
Chicago/Turabian StyleYuan, Xiaolong, Xiaofeng Zhou, Yingting He, Yuyi Zhong, Ailing Zhang, Zhe Zhang, Hao Zhang, and Jiaqi Li. 2018. "C/EBPβ Promotes STAT3 Expression and Affects Cell Apoptosis and Proliferation in Porcine Ovarian Granulosa Cells" Genes 9, no. 6: 295. https://doi.org/10.3390/genes9060295
APA StyleYuan, X., Zhou, X., He, Y., Zhong, Y., Zhang, A., Zhang, Z., Zhang, H., & Li, J. (2018). C/EBPβ Promotes STAT3 Expression and Affects Cell Apoptosis and Proliferation in Porcine Ovarian Granulosa Cells. Genes, 9(6), 295. https://doi.org/10.3390/genes9060295