Cytogenetics in Arctica islandica (Bivalvia, Arctidae): the Longest Lived Non-Colonial Metazoan



Abstract
:1. Introduction
2. Materials and Methods
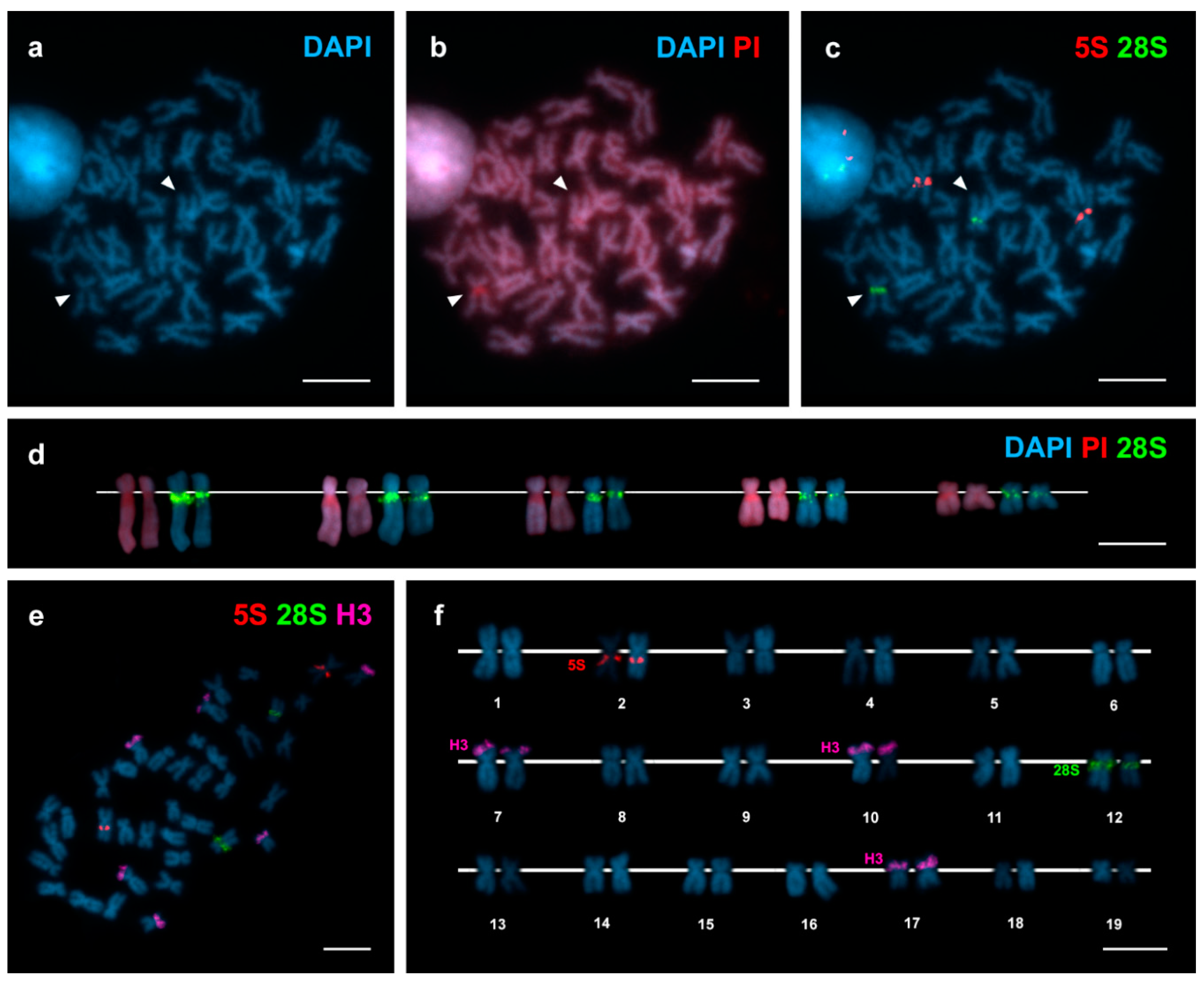
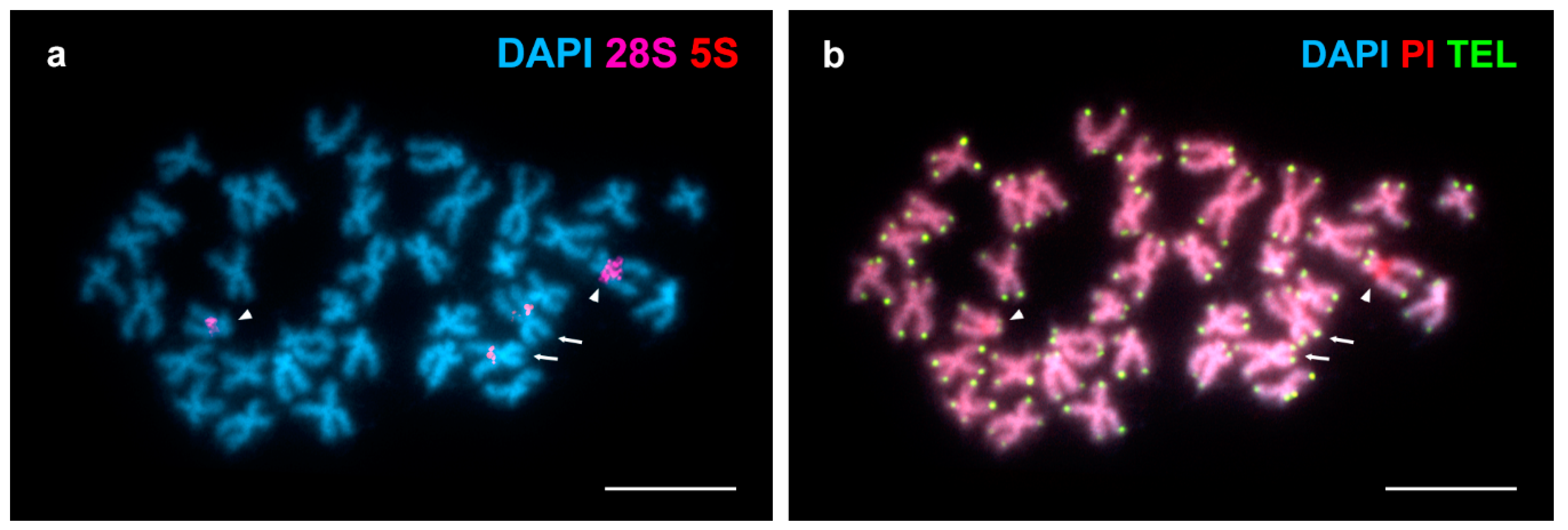
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Morton, B. The biology and functional morphology of Arctica islandica (Bivalvia: Arcticidae): A gerontophilic living fossil. Mar. Biol. Res. 2011, 7, 540–553. [Google Scholar] [CrossRef]
- Ridgway, I.D.; Richardson, C.A. Arctica islandica: The longest lived non colonial animal known to science. Rev. Fish Biol. Fish. 2011, 21, 297–310. [Google Scholar] [CrossRef]
- Hennen, D.R. How should we harvest an animal that can live for centuries? North Am. J. Fish. Manag. 2015, 35, 512–527. [Google Scholar] [CrossRef]
- Abele, D.; Strahl, J.; Brey, T.; Philipp, E.E.R. Imperceptible senescence: Ageing in the ocean quahog Arctica islandica. Free Radic. Res. 2008, 42, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipp, E.E.R.; Abele, D. Masters of longevity: Lessons from long-lived bivalves—A mini-review. Gerontology 2010, 56, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, M.; Wapstra, E.; Friesen, C. Ectothermic telomeres: It’s time they came in from the cold. Philos. Trans. R. Soc. B 2018, 373, 20160449. [Google Scholar] [CrossRef] [PubMed]
- Schöne, B.R. Arctica islandica (Bivalvia): A unique paleoenvironmental archive of the northern North Atlantic Ocean. Glob. Planet. Change 2013, 111, 199–225. [Google Scholar] [CrossRef]
- Dahlgren, T.G.; Weinberg, J.R.; Halanych, K.M. Phylogeography of the ocean quahog (Arctica islandica): Influences of paleoclimate on genetic diversity and species range. Mar. Biol. 2000, 137, 487–495. [Google Scholar] [CrossRef]
- Holmes, S.P.; Witbaard, R.; van der Meer, J. Phenotypic and genotypic population differentiation in the bivalve mollusc Arctica islandica: Results from RAPD analysis. Mar. Ecol. Prog. Ser. 2003, 254, 163–176. [Google Scholar] [CrossRef]
- Philipp, E.E.R.; Wessels, W.; Gruber, H.; Strahl, J.; Wagner, A.E.; Ernst, I.M.A.; Rimbach, G.; Kraemer, L.; Schreiber, S.; Abele, D.; et al. Gene expression and physiological changes of different populations of the long-lived bivalve Arctica islandica under low oxygen conditions. PLoS ONE 2012, 7, e44621. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.; Schaible, R.; Ridgway, I.D.; Chowd, T.T.; Helde, C.; Philipp, E.E.R. Telomere-independent ageing in the longest-lived non-colonial animal, Arctica islandica. Exp. Gerontol. 2014, 51, 38–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GOLD Genomes OnLine Database. Available online: https://gold.jgi.doe.gov/project?id=38667 (accessed on 12 February 2018).
- Glöckner, G.; Heinze, I.; Platzer, M.; Held, C.; Abele, D. The mitochondrial genome of Arctica islandica; Phylogeny and variation. PLoS ONE 2013, 8, e82857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dégletagne, C.; Abele, D.; Held, C. A distinct mitochondrial genome with DUI-like inheritance in the ocean quahog Arctica islandica. Mol. Biol. Evol. 2015, 33, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, J.; Nantón, A.; Ruiz-Ruano, F.J.; Camacho, J.P.M.; Méndez, J. First complete female mitochondrial genome in four bivalve species genus Donax and their phylogenetic relationships within the Veneroida order. PLoS ONE 2017, 12, e0184464. [Google Scholar] [CrossRef] [PubMed]
- Insua, A.; Freire, R.; Méndez, J. The 5S rDNA of the bivalve Cerastoderma edule: Nucleotide sequence of the repeat unit and chromosomal location relative to 18-28S rDNA. Genet. Sel. Evol. 1999, 31, 509–518. [Google Scholar] [CrossRef]
- Martínez, A.; Marinas, L.; González-Tizón, A.; Méndez, J. Cytogenetic characterization of Donax trunculus (Bivalvia: Donacidae) by means of karyotyping, fluorochrome banding and fluorescent in situ hybridization. J. Molluscan Stud. 2002, 68, 393–396. [Google Scholar] [CrossRef]
- Plohl, M.; Prats, E.; Martínez-Lage, A.; González-Tizón, A.; Méndez, J.; Cornudella, L. Telomeric localization of the vertebrate-type hexamer repeat (TTAGGG)n in the wedgeshell clam Donax trunculus and other marine invertebrate genomes. J. Biol. Chem. 2002, 277, 19839–19846. [Google Scholar] [CrossRef] [PubMed]
- Petrović, V.; Pérez-García, C.; Pasantes, J.J.; Šatović, E.; Prats, E.; Plohl, M. A GC-rich satellite DNA and karyology of the bivalve mollusk Donax trunculus: A dominance of GC-rich heterochromatin. Cytogenet. Genome Res. 2009, 124, 63–71. [Google Scholar] [CrossRef] [PubMed]
- García-Souto, D.; Pérez-García, C.; Pasantes, J.J. Are pericentric inversions reorganizing wedge shell genomes? Genes 2017, 8, 370. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, X. Chromosomal mapping of the vertebrate telomeric sequence (TTAGGG)n in four bivalve molluscs by fluorescence in situ hybridization. J. Shellfish Res. 2001, 20, 1187–1190. [Google Scholar]
- Wang, Y.; Guo, X. Chromosomal mapping of the major ribosomal RNA genes in the dwarf surfclam (Mulinia lateralis Say). J. Shellfish Res. 2008, 27, 307–311. [Google Scholar] [CrossRef]
- García-Souto, D.; Pérez-García, C.; Kendall, J.; Pasantes, J.J. Molecular cytogenetics in trough shells (Mactridae, Bivalvia): Divergent GC-rich heterochromatin content. Genes 2016, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- García-Souto, D.; Mravinac, B.; Šatović, E.; Plohl, M.; Morán, P.; Pasantes, J.J. Methylation profile of a satellite DNA constituting the intercalary G+C-rich heterochromatin of the cut trough shell Spisula subtruncata (Bivalvia, Mactridae). Sci. Rep. 2017, 7, 6930. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tajes, J.; Martínez-Lage, A.; Freire, R.; Guerra, A.; Méndez, J.; González-Tizón, A.M. Genome sizes and karyotypes in the razor clams Ensis arcuatus (Jeffreys, 1985) and E. siliqua (Linnaeus, 1758). Cah. Biol. Mar. 2008, 49, 79–85. [Google Scholar] [CrossRef]
- González-Tizón, A.; Rojo, V.; Vierna, J.; Jensen, K.T.; Egea, E.; Martínez-Lage, A. Cytogenetic characterisation of the razor shells Ensis directus (Conrad, 1843) and E. minor (Chenu, 1843) (Mollusca: Bivalvia). Helgol. Mar. Res. 2013, 67, 73–82. [Google Scholar] [CrossRef]
- González-Tizón, A.M.; Martínez-Lage, A.; Rego, I.; Ausió, J.; Méndez, J. DNA content, karyotypes, and chromosomal location of 18S-5.8S28S¨C ribosomal loci in some species of bivalve molluscs from the pacific Canadian coast. Genome 2000, 43, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, N.; Pasantes, J.J. Surface spreading of synaptonemal complexes in the clam Dosinia exoleta (Mollusca, Bivalvia). Chromosome Res. 2005, 13, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, X. Chromosomal mapping of major ribosomal rRNA genes in the hard clam (Mercenaria mercenaria) using fluorescent hybridization. Mar. Biol. 2007, 150, 1183–1189. [Google Scholar] [CrossRef]
- Carrilho, J.; Pérez-García, C.; Leitão, A.; Malheiro, I.; Pasantes, J.J. Cytogenetic characterization and mapping of rDNAs, core histone genes and telomeric sequences in Venerupis aurea and Tapes rhomboides (Bivalvia: Veneridae). Genetica 2011, 139, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, N.; Pérez-García, C.; Morán, P.; Pasantes, J.J. Genetic and cytological evidence of hybridization between native Ruditapes decussatus and introduced Ruditapes philippinarum (Mollusca, Bivalvia, Veneridae) in NW Spain. Aquaculture 2011, 311, 123–128. [Google Scholar] [CrossRef]
- Pérez-García, C.; Hurtado, N.S.; Morán, P.; Pasantes, J.J. Evolutionary dynamics of rDNA clusters in chromosomes of five clam species belonging to the family Veneridae (Mollusca, Bivalvia). BioMed Res. Int. 2014, 2014, 754012. [Google Scholar] [CrossRef] [PubMed]
- García-Souto, D.; Pérez-García, C.; Morán, P.; Pasantes, J.J. Divergent evolutionary behavior of H3 histone gene and rDNA clusters in venerid clams. Mol. Cytogenet. 2015, 8, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Souto, D.; Qarkaxhija, V.; Pasantes, J.J. Resolving the taxonomic status of Chamelea gallina and C. striatula (Veneridae, Bivalvia): A combined molecular cytogenetic and phylogenetic approach. BioMed Res. Int. 2017, 2017, 7638790. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tajes, J.; González-Tizón, A.; Martínez-Lage, A.; Méndez, J. Cytogenetics of the razor clam Solen marginatus (Mollusca. Bivalvia: Solenidae). Cytogenet. Genome Res. 2003, 101, 43–46. [Google Scholar] [CrossRef]
- García-Souto, D.; Rios, G.; Pasantes, J.J. Karyotype differentiation in tellin shells (Bivalvia: Tellinidae). BMC Genet. 2017, 18, 66. [Google Scholar] [CrossRef] [PubMed]
- Thalasa Tradition. Available online: http://www.thalassa-tradition.fr (accessed on 15 January 2018).
- Estación de Ciencias Mariñas de Toralla (ECIMAT). Available online: https://ecimat.uvigo.es/en/ (accessed on 15 January 2018).
- Méndez, J.; Pasantes, J.J.; Martínez-Expósito, M.J. Banding pattern of mussel (Mytilus galloprovincialis) chromosomes induced by 2 × SSC/Giemsa-stain treatment. Mar. Biol. 1990, 106, 375–377. [Google Scholar] [CrossRef]
- Martínez-Expósito, M.J.; Pasantes, J.J.; Méndez, J. Proliferation kinetics of mussel (Mytilus galloprovincialis) gill cells. Mar. Biol. 1994, 120, 41–45. [Google Scholar] [CrossRef]
- Pérez-García, C.; Cambeiro, J.M.; Morán, P.; Pasantes, J.J. Chromosomal mapping of rDNAs, core histone genes and telomeric sequences in Perumytilus purpuratus (Bivalvia: Mytilidae). J. Exp. Mar. Biol. Ecol. 2010, 395, 199–205. [Google Scholar] [CrossRef]
- Pérez-García, C.; Guerra-Varela, J.; Morán, P.; Pasantes, J.J. Chromosomal mapping of rRNA genes, core histone genes and telomeric sequences in Brachidontes puniceus and Brachidontes rodriguezi (Bivalvia: Mytilidae). BMC Genet. 2010, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, C.; Morán, P.; Pasantes, J.J. Cytogenetic characterization of the invasive mussel species Xenostrobus securis Lmk. (Bivalvia: Mytilidae). Genome 2011, 54, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, C.; Morán, P.; Pasantes, J.J. Karyotypic diversification in Mytilus mussels (Bivalvia: Mytilidae) inferred from chromosomal mapping of rRNA and histone gene clusters. BMC Genet. 2014, 15, 84. [Google Scholar] [CrossRef] [PubMed]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Nakamura, H.K. A review of molluscan cytogenetic information based on the CISMOCH-computerized system for molluscan chromosomes. Bivalvia, Polyplacophora and Cephalopoda. Venus (Jap. J. Malacol.) 1985, 44, 193–225. [Google Scholar]
- Thiriot-Quiévreux, C. Advances in cytogenetics of aquatic organisms. In Genetics and evolution of aquatic organisms; Beaumont, A.R., Ed.; Chapman and Hall: London, UK, 1994; pp. 369–388. ISBN 9780412493706. [Google Scholar]
- Thiriot-Quiévreux, C. Review of the literature on bivalve cytogenetics in the last ten years. Cah. Biol. Mar. 2002, 43, 17–26. [Google Scholar]
- Leitão, A.; Chaves, R. Banding for chromosomal identification in bivalves: A 20-year history. In Aquaculture 1 Dynamic Biochemistry, Process Biotechnology and Molecular Biology 2; Russo, R., Ed.; Global Science Books: Ikenobe, Japan, 2008; pp. 44–49. Available online: http://www.globalsciencebooks.info/Online/GSBOnline/OnlineDBPBMB_2_SI1.html (accessed on 10 February 2018).
- Arias-Pérez, A.; Insua, A.; Freire, R.; Méndez, J.; Fernández-Tajes, J.M. Genetic studies in commercially important species of Veneridae. In Clam Fisheries and Aquaculture; Da Costa González, F., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 73–105. ISBN 9781622575183. [Google Scholar]
- Skuza, L.; Labecka, A.M.; Domagala, J. Cytogenetic and morphological characterization of Corbicula fluminalis (O. F. Müller, 1774) (Bivalvia: Veneroida: Corbiculidae): Taxonomic status assessment of a freshwater clam. Folia Biol. (Kraków) 2009, 57, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Petkevičiūtė, R.; Stunzėnas, V.; Stanevičiūtė, G. Comments on species divergence in the genus Sphaerium (Bivalvia) and phylogenetic affinities of Sphaerium nucleus and S. corneum var. mamillanum based on karyotypes and sequences of 16S and ITS1 rDNA. PLoS ONE 2018, 13, e0191427. [Google Scholar]
- García-Souto, D.; Sumner-Hempel, A.; Fervenza, S.; Pérez-García, C.; Torreiro, A.; González-Romero, R.; Eirín-López, J.M.; Morán, P.; Pasantes, J.J. Detection of invasive and cryptic species in marine mussels (Bivalvia, Mytilidae): A chromosomal perspective. J. Nat. Conserv. 2017, 39, 58–67. [Google Scholar] [CrossRef]
- Lansdorp, P.M.; Verwoerd, N.P.; van de Rijke, F.M.; Dragowska, V.; Little, M.T.; Dirks, R.W.; Raap, A.K.; Tanke, H.J. Heterogeneity in telomere length of human chromosomes. Hum. Mol. Genet. 1996, 5, 685–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemann, M.T.; Strong, M.A.; Hao, L.Y.; Greider, C.W. The shortest telomere, not average telomere length, is critical for cell viability and chromosome stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef]
- Vera, E.; Blasco, M.A. Beyond average: Potential for measurement of short telomeres. Aging (Albany NY) 2012, 4, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Ingles, E.D.; Deakin, J.E. Telomeres, species differences, and unusual telomeres in vertebrates: Presenting challenges and opportunities to understanding telomere dynamics. AIMS Genet. 2016, 3, 1–24. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Pair | Relative Length | Centromeric Index | Type * | ||
|---|---|---|---|---|---|
| Mean | SE | Mean | SE | ||
| 1 | 6.86 | 0.17 | 41.35 | 0.45 | m |
| 2 | 6.12 | 0.10 | 36.34 | 0.74 | sm |
| 3 | 6.05 | 0.10 | 42.20 | 0.67 | m |
| 4 | 5.97 | 0.13 | 30.39 | 4.01 | sm |
| 5 | 5.80 | 0.12 | 32.31 | 3.01 | sm |
| 6 | 5.49 | 0.08 | 29.51 | 1.97 | sm |
| 7 | 5.41 | 0.12 | 33.94 | 1.72 | sm |
| 8 | 5.38 | 0.09 | 34.37 | 1.74 | sm |
| 9 | 5.37 | 0.08 | 34.61 | 2.23 | sm |
| 10 | 5.22 | 0.11 | 41.76 | 0.91 | m |
| 11 | 5.21 | 0.04 | 35.47 | 1.34 | sm |
| 12 | 5.15 | 0.11 | 30.07 | 1.11 | sm |
| 13 | 5.08 | 0.06 | 33.39 | 1.68 | sm |
| 14 | 4.94 | 0.07 | 32.09 | 1.32 | sm |
| 15 | 4.92 | 0.10 | 29.50 | 1.33 | sm |
| 16 | 4.79 | 0.06 | 34.37 | 1.69 | sm |
| 17 | 4.58 | 0.05 | 37.10 | 1.81 | sm/m |
| 18 | 3.94 | 0.07 | 33.50 | 1.01 | sm |
| 19 | 3.71 | 0.11 | 43.41 | 0.89 | m |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Souto, D.; Pasantes, J.J. Cytogenetics in Arctica islandica (Bivalvia, Arctidae): the Longest Lived Non-Colonial Metazoan. Genes 2018, 9, 299. https://doi.org/10.3390/genes9060299
García-Souto D, Pasantes JJ. Cytogenetics in Arctica islandica (Bivalvia, Arctidae): the Longest Lived Non-Colonial Metazoan. Genes. 2018; 9(6):299. https://doi.org/10.3390/genes9060299
Chicago/Turabian StyleGarcía-Souto, Daniel, and Juan J. Pasantes. 2018. "Cytogenetics in Arctica islandica (Bivalvia, Arctidae): the Longest Lived Non-Colonial Metazoan" Genes 9, no. 6: 299. https://doi.org/10.3390/genes9060299
APA StyleGarcía-Souto, D., & Pasantes, J. J. (2018). Cytogenetics in Arctica islandica (Bivalvia, Arctidae): the Longest Lived Non-Colonial Metazoan. Genes, 9(6), 299. https://doi.org/10.3390/genes9060299