Membrane Surface Features of Blastocystis Subtypes

{kind=link}
{kind=link}
Abstract
:1. Introduction
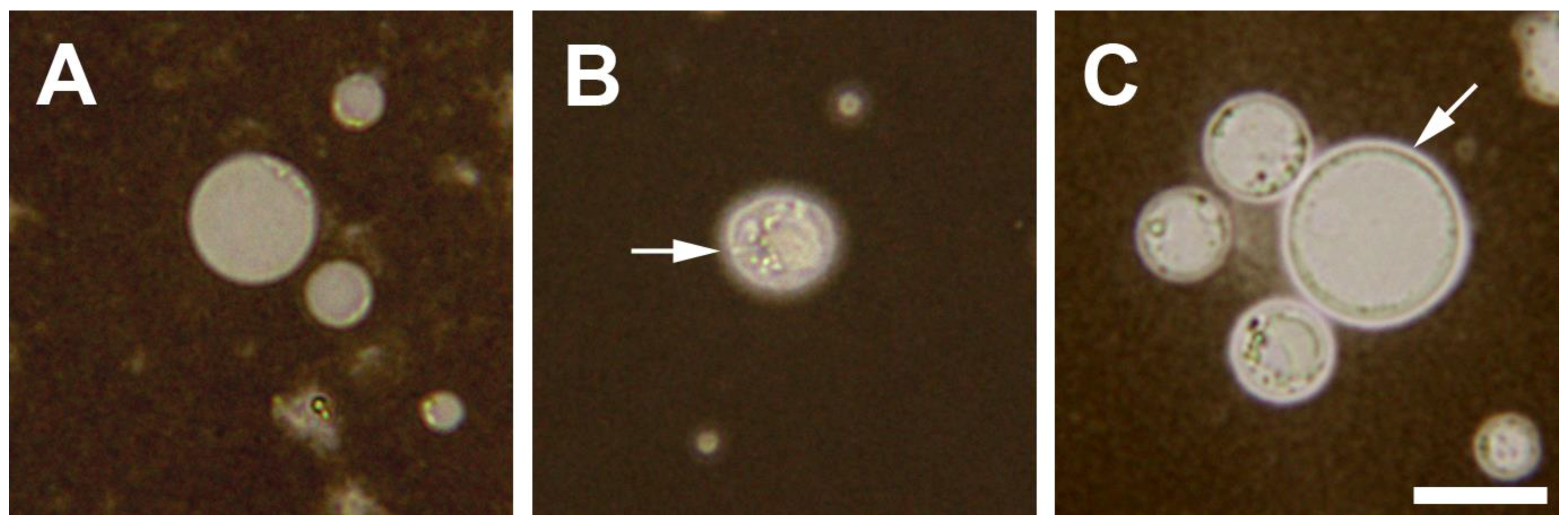
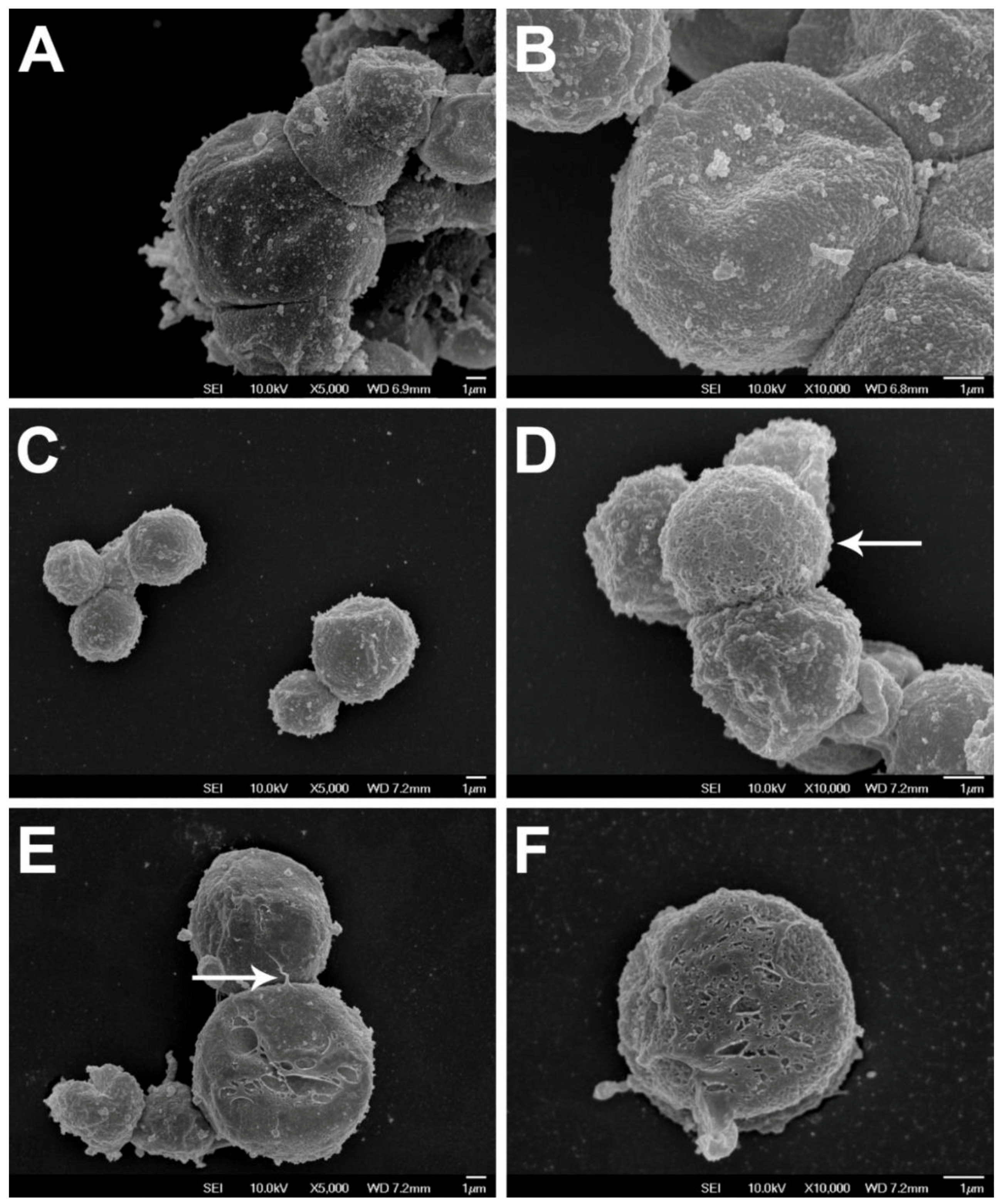
2. Blastocystis Surface Features
2.1. Surface Coat Structure
2.2. Surface Coat Variations
2.3. Role of Surface Coat
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clark, C.G.; van der Giezen, M.; Alfellani, M.A.; Stensvold, C.R. Chapter One—Recent Developments in Blastocystis Research. In Advances in Parasitology, 1st ed.; Rollinson, D., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 82, pp. 1–32. [Google Scholar]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.O.; Stensvold, C.R. Blastocystis in health and disease: Are we moving from a clinical to a public health perspective? J. Clin. Microbiol. 2016, 54, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Gentekaki, E.; Curtis, B.A.; Stairs, C.W.; Klimeš, V.; Eliáš, M.; Salas-Leiva, D.E.; Herman, E.K.; Eme, L.; Arias, M.C.; Henrissat, B.; et al. Extreme genome diversity in the hyper-prevalent parasitic eukaryote Blastocystis. PLoS Biol. 2017, 15, e2003769. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.; Stark, D.; Harkness, J.; Ellis, J. Update on the pathogenic potential and treatment options for Blastocystis sp. Gut Pathog. 2014, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Govind, S.K.; Khairul, A.A.; Smith, H.V. Multiple reproductive processes in Blastocystis. Trends Parasitol. 2002, 18, 528. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Suresh, G.K.; Tan, K.S.W.; Thompson, R.C.A.; Traub, R.J.; Viscogliosi, E.; Yoshikawa, H.; Clark, C.G. Terminology for Blastocystis subtypes—A consensus. Trends Parasitol. 2007, 23, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Yason, J.A.; Tan, K.S.W. Seeing the Whole Elephant: Imaging Flow Cytometry Reveals Extensive Morphological Diversity within Blastocystis Isolates. PLoS ONE 2015, 10, e0143974. [Google Scholar] [CrossRef] [PubMed]
- Mirza, H.; Wu, Z.; Kidwai, F.; Tan, K.S.W. A metronidazole-resistant isolate of Blastocystis spp. is susceptible to nitric oxide and downregulates intestinal epithelial inducible nitric oxide synthase by a novel parasite survival mechanism. Infect. Immun. 2011, 79, 5019–5026. [Google Scholar] [CrossRef] [PubMed]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.U.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Flórez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; López, M.C.; García, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Mirza, H.; Tan, K.S.W. Intra-subtype variation in enteroadhesion accounts for differences in epithelial barrier disruption and is associated with metronidazole resistance in Blastocystis subtype-7. PLoS Negl. Trop. Dis. 2014, 8, e2885. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Mirza, H.; Teo, J.D.W.; Tan, K.S.W. Strain-dependent induction of human enterocyte apoptosis by Blastocystis disrupts epithelial barrier and ZO-1 organization in a caspase 3- and 9-dependent manner. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Mirza, H.; Teo, J.D.W.; Upcroft, J.; Tan, K.S.W. A Rapid, High-throughput viability assay for Blastocystis spp. reveals metronidazole resistance and extensive subtype-dependent variations in drug susceptibilities. Antimicrob. Agents Chemother. 2011, 55, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Yason, J.A.; Ajjampur, S.S.R.; Tan, K.S.W. Blastocystis isolate B exhibits multiple modes of resistance against antimicrobial peptide LL-37. Infect. Immun. 2016, 84, 2220–2232. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.K.; Zierdt, C.H. Ultrastructure of Blastocystis hominis. Z. Parasitenkd. 1973, 42, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Yamada, M.; Yoshida, Y. Light-microscopical appearance and ultrastructure of Blastocystis hominis, an intestinal parasite of man. Zentralbl. Bakteriol. Mikrobiol. Hyg. A 1987, 264, 379–385. [Google Scholar] [CrossRef]
- Dunn, L.A.; Boreham, P.F.L.; Stenzel, D.J. Ultrastructural variation of Blastocystis hominis stocks in culture. Int. J. Parasitol. 1989, 19, 43–56. [Google Scholar] [CrossRef]
- Stenzel, D.J.; Boreham, P.F.L.; McDougall, R. Ultrastructure of Blastocystis hominis in human stool samples. Int. J. Parasitol. 1991, 21, 807–812. [Google Scholar] [CrossRef]
- Cassidy, M.F.; Stenzel, D.J.; Boreham, P.F.L. Electron microscopy of surface structures of Blastocystis sp. from different hosts. Parasitol. Res. 1994, 80, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Kuwayama, N.; Enose, Y. Histochemical detection of carbohydrates of Blastocystis hominis. J. Eukaryot. Microbiol. 1995, 42, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Lanuza, M.D.; Carbajal, J.A.; Borrás, R. Identification of surface coat carbohydrates in Blastocystis hominis by lectin probes. Int. J. Parasitol. 1996, 26, 527–532. [Google Scholar] [CrossRef]
- Guha-Niyogi, A.; Sullivan, D.R.; Turco, S.J. Glycoconjugate structures of parasitic protozoa. Glycobiology 2001, 11, 45R–59R. [Google Scholar] [CrossRef] [PubMed]
- McConville, M.J. The surface glycoconjugates of parasitic protozoa: Potential targets for new drugs. Aust. N. Z. J. Med. 1995, 25, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Inge, P.M.; Edson, C.M.; Farthing, M.J. Attachment of Giardia lamblia to rat intestinal epithelial cells. Gut 1988, 29, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Sher, A. Evasion of innate immunity by parasitic protozoa. Nat. Immunol. 2002, 3, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.S.W.; Ng, G.C.; Quek, E.; Howe, J.; Ramachandran, N.P.; Yap, E.H.; Singh, M. Blastocystis hominis: A simplified, high-efficiency method for clonal growth on solid agar. Exp. Parasitol. 2000, 96, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.W.; Singh, M.; Ho, L.C.; Howe, J.; Moe, K.T.; Chen, X.Q.; Ng, G.C.; Yap, E.H. Survival of Blastocystis hominis clones after exposure to a cytotoxic monoclonal antibody. Int. J. Parasitol. 1997, 27, 947–954. [Google Scholar] [CrossRef]
- Wu, B.; Yin, J.; Texier, C.; Roussel, M.; Tan, K.S.W. Blastocystis legumain is localized on the cell surface, and specific inhibition of its activity implicates a pro-survival role for the enzyme. J. Biol. Chem. 2010, 285, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Gentekaki, E.; Curtis, B.; Archibald, J.M.; Roger, A.J. Lateral gene transfer in the adaptation of the anaerobic parasite Blastocystis to the Gut. Curr. Biol. 2017, 27, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Zaman, V.; Howe, J.; Ng, M.; Goh, T.K. Scanning electron microscopy of the surface coat of Blastocystis hominis. Parasitol. Res. 1999, 85, 974–976. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yason, J.A.; Tan, K.S.W. Membrane Surface Features of Blastocystis Subtypes. Genes 2018, 9, 417. https://doi.org/10.3390/genes9080417
Yason JA, Tan KSW. Membrane Surface Features of Blastocystis Subtypes. Genes. 2018; 9(8):417. https://doi.org/10.3390/genes9080417
Chicago/Turabian StyleYason, John Anthony, and Kevin Shyong Wei Tan. 2018. "Membrane Surface Features of Blastocystis Subtypes" Genes 9, no. 8: 417. https://doi.org/10.3390/genes9080417
APA StyleYason, J. A., & Tan, K. S. W. (2018). Membrane Surface Features of Blastocystis Subtypes. Genes, 9(8), 417. https://doi.org/10.3390/genes9080417