Aerosol Microbiome over the Mediterranean Sea Diversity and Abundance

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Methods and Materials
2.1. Sampling
2.2. Aerosol Optical Depth
2.3. Air Mass Backward Trajectories
2.4. Region and Distance to Land
2.5. Aluminum
2.6. Bacterial Abundance
2.7. DNA Extraction, Amplification, Sequencing
2.8. Bioinformatics
3. Results and Discussion
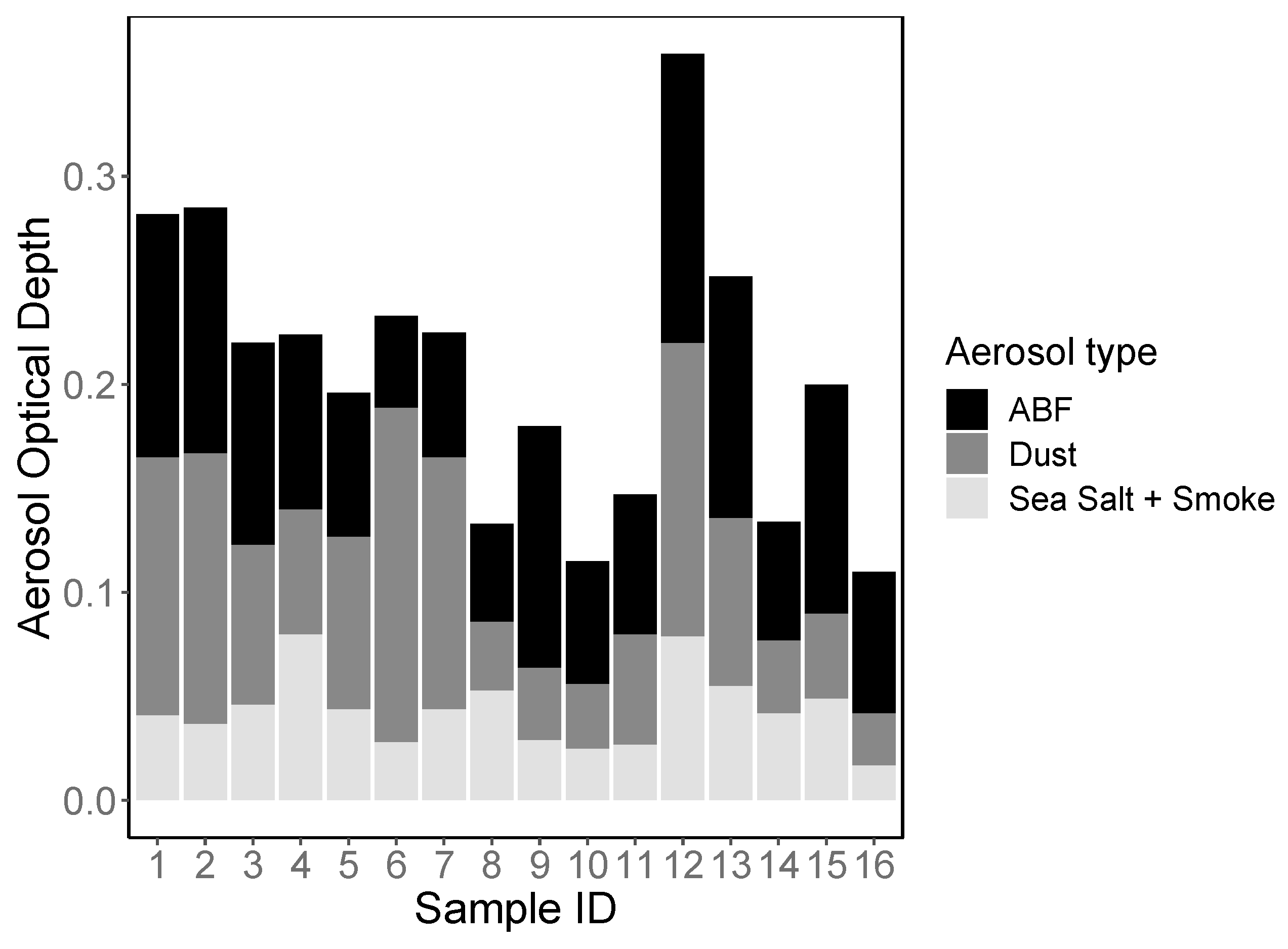
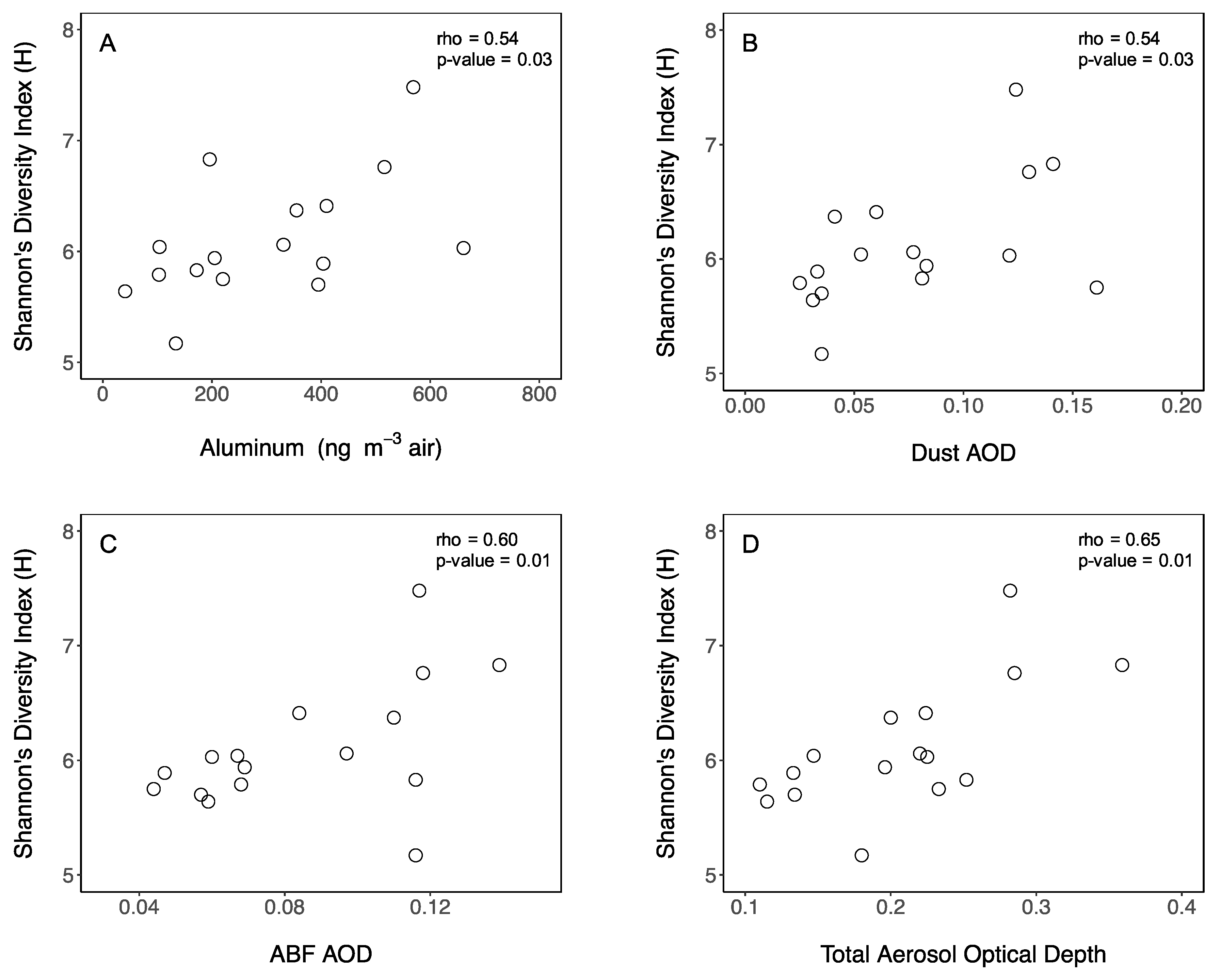
3.1. Aerosol Origin and Chemical Properties
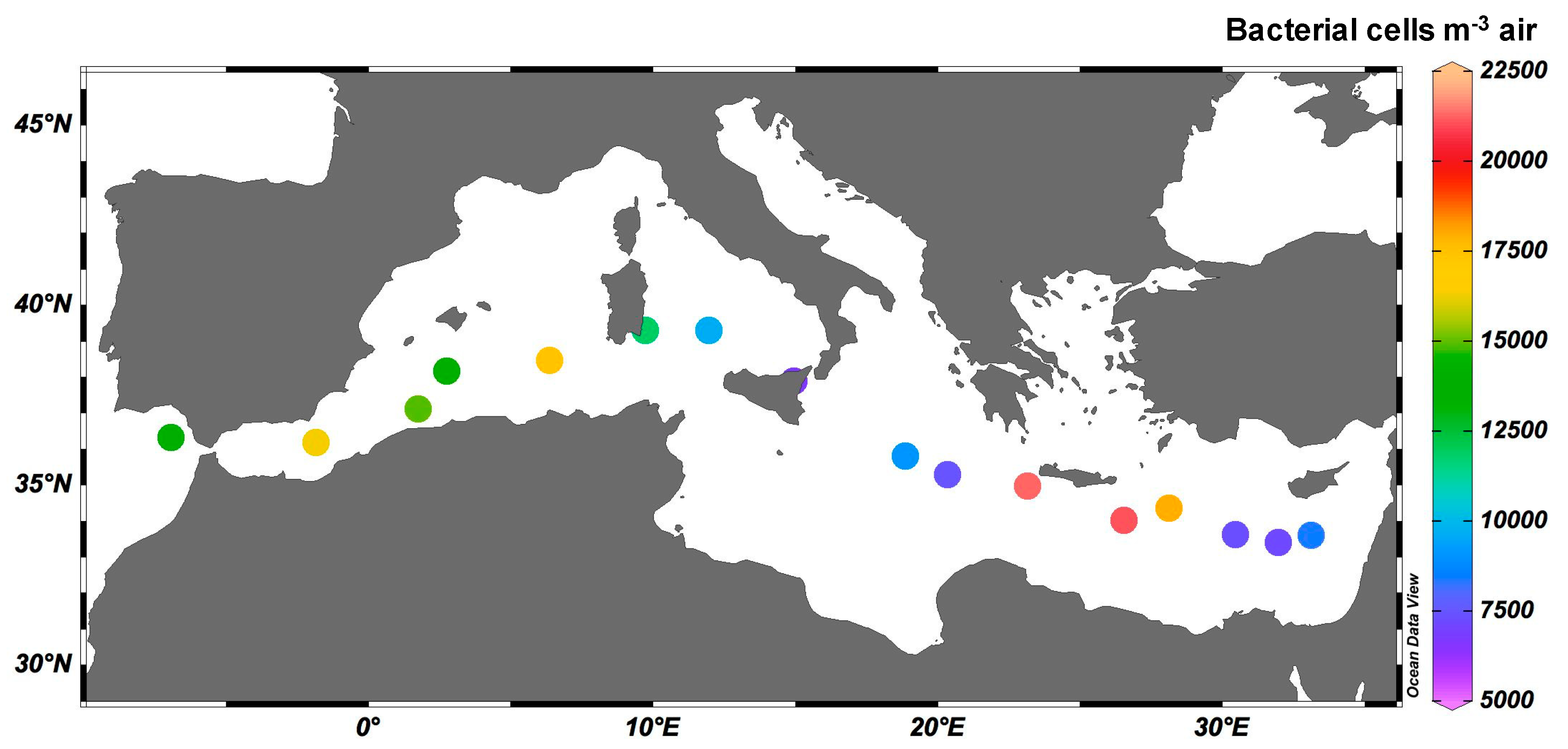
3.2. Airborne Bacterial Abundance
3.3. Airborne Microbiome above the MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayol, E.; Arrieta, J.M.; Jiménez, M.A.; Martínez-Asensio, A.; Garcias-Bonet, N.; Dachs, J.; González-Gaya, B.; Royer, S.J.; Benítez-Barrios, V.M.; Fraile-Nuez, E.; et al. Long-range transport of airborne microbes over the global tropical and subtropical ocean. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Torsvik, V.; Øvreås, L.; Thingstad, T.F. Prokaryotic diversity—Magnitude dynamics, and controlling factors. Science 2002, 296, 1064–1066. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.W.; Kubilay, N.; Kocak, M.; Gray, M.A.; Borden, T.C.; Shinn, E.A. Airborne desert dust and aeromicrobiology over the Turkish Mediterranean coastline. Atmos. Environ. 2007, 41, 4050–4062. [Google Scholar] [CrossRef]
- Prospero, J.M.; Charlson, R.J.; Mohnen, V.; Jaenicke, R.; Delany, A.C.; Moyers, J.; Zoller, W.; Rahn, K. The atmospheric aerosol system—An overview. Rev. Geophys. Space Phys. 1983, 21, 1607–1629. [Google Scholar] [CrossRef]
- Huebert, B.J. Sea Salt Aerosols. Encyclopedia of Earth System Science; Nierenberg, W.A., Ed.; Academic Press: San Diego, CA, USA, 1992; Volume 4, pp. 63–68. [Google Scholar]
- Bovallius, A.; Bucht, B.; Roffey, R.; Anas, P. Long-range air transmission of bacteria. Appl. Environ. Microbiol. 1978, 35, 1231–1232. [Google Scholar] [PubMed]
- Prospero, J.M.; Blades, E.; Mathison, G.; Naidu, R. Interhemispheric transport of viable fungi and bacteria from Africa to the Caribbean with soil dust. Aerobiologia 2005, 21, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.; Kellogg, C.A.; Garrison, V.; Lisle, J.; Borden, T.; Shinn, E. Atmospheric microbiology in the northern Caribbean during African dust events. Aerobiologia 2003, 19, 143–157. [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Rahav, E.; Belkin, N.; Paytan, A.; Herut, B. The Relationship between Air-Mass Trajectories and the Abundance of Dust-Borne Prokaryotes at the SE Mediterranean Sea. Atmosphere 2019, 10, 280. [Google Scholar] [CrossRef]
- Estillore, A.D.; Trueblood, J.V.; Grassian, V.H. Atmospheric chemistry of bioaerosols: Heterogeneous and multiphase reactions with atmospheric oxidants and other trace gases. Chem. Sci. 2016, 7, 6604–6616. [Google Scholar] [CrossRef]
- Brągoszewska, E.; Pastuszka, J.S. Influence of meteorological factors on the level and characteristics of culturable bacteria in the air in Gliwice, Upper Silesia (Poland). Aerobiologia 2016, 34, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Womack, A.M.; Bohannan, B.J.M.; Green, J.L. Biodiversity and biogeography of the atmosphere. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 3645–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caliz, J.; Triadó-Margarit, X.; Camarero, L.; Casamayor, E.O. A long-term survey unveils strong seasonal patterns in the airborne microbiome coupled to atmospheric circulations. Proc. Natl. Acad. Sci. USA 2018, 115, 12229–12234. [Google Scholar] [CrossRef] [PubMed]
- Reche, I.; D’Orta, G.; Mladenov, N.; Winget, D.M.; Suttle, C.A. Deposition rates of viruses and bacteria above the atmospheric boundary layer. ISME J. 2018, 12, 1154–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodie, E.L.; DeSantis, T.Z.; Parker, J.P.M.; Zubietta, I.X.; Piceno, Y.M.; Andersen, G.L. Urban aerosols harbor diverse and dynamic bacterial populations. Proc. Natl. Acad. Sci. USA 2007, 104, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.K.M.; Hovmoller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef]
- Weir-Brush, J.; Garrison, V.H.; Smith, G.W.; Shinn, E.A. The Relationship Between Gorgonian Coral (Cnidaria: Gorgonacea) Diseases and African Dust Storms. Aerobiologia 2004, 20, 119–126. [Google Scholar] [CrossRef]
- Kaarakainen, P.; Meklin, T.; Rintala, H.; Hyvärinen, A.; Kärkkäinen, P.; Vepsäläinen, A.; Hirvonen, M.-R.; Nevalainen, A. Seasonal Variation in Airborne Microbial Concentrations and Diversity at Landfill, Urban and Rural Sites. Clean 2008, 36, 556–563. [Google Scholar] [CrossRef]
- Guieu, C.; Aumont, O.; Paytan, A.; Bopp, L.; Law, C.S.; Mahowald, N.; Achterberg, E.P.; Marañón, E.; Salihoglu, B.; Crise, A.; et al. Global biogeochemical cycles deposition to Low Nutrient Low Chlorophyll regions. Global Biogeochem. Cycles 2014, 28, 1179–1198. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef]
- Tanhua, T.; Hainbucher, D.; Schroeder, K.; Cardin, V.; Álvarez, M.; Civitarese, G. The Mediterranean Sea system: A review and a nintroduction to the special issue. Ocean Sci. 2013, 9, 789–803. [Google Scholar] [CrossRef]
- Lawrence, C.R.; Neff, J.C. The contemporary physical and chemical flux of aeolian dust: A synthesis of direct measurements of dust deposition. Chem. Geol. 2009, 267, 46–63. [Google Scholar] [CrossRef]
- Loye-Pilot, M.D.; Martin, J.M.; Morelli, J. Atmospheric input of inorganic nitrogen to the Western Mediterranean. Biogeochemistry 1990, 9, 117–134. [Google Scholar] [CrossRef]
- Bergametti, G.; Remoudaki, E.; Losno, R.; Steiner, E.; Chatenet, B.; Buat-Ménard, P. Source, transport and deposition of atmospheric phosphorus over the northwestern Mediterranean. J. Atmos. Chem. 1992, 14, 501–513. [Google Scholar] [CrossRef]
- Quétel, C.R.; Remoudaki, E.; Davies, J.E.; Miquel, J.-C.; Fowler, S.W.; Lambert, C.E.; Bergametti, G.; Buat-Ménard, P. Impact of atmospheric deposition on particulate iron flux and distribution in northwestern Mediterranean waters. Deep Sea Res. 1993, 40, 989–1002. [Google Scholar] [CrossRef]
- Herut, B.; Krom, M.D.D.; Pan, G.; Mortimer, R. Atmospheric input of nitrogen and phosphorus to the Southeast Mediterranean: Sources, fluxes, and possible impact. Limnol. Oceanogr. 1999, 44, 1683–1692. [Google Scholar] [CrossRef] [Green Version]
- Ridame, C.; Guieu, C. Saharan input of phosphorus to the oligotrophic water of the open western Mediterranean. Limnol. Oceanogr. 2002, 47, 856–869. [Google Scholar] [CrossRef]
- Ridame, C.; Guieu, C.; L’Helguen, S. Strong stimulation of N2 fixation to contrasted Saharan dust events in a Low Nutrient-Low Chlorophyll environment: Results from dust addition in large mesocosms. Biogeosciences 2013, 10, 7333–7346. [Google Scholar] [CrossRef]
- Rahav, E.; Ovadia, G.; Paytan, A.; Herut, B. Contribution of airborne microbes to bacterial production and N2 fixation in seawater upon aerosol deposition. Geophys. Res. Lett. 2016, 43, 719–727. [Google Scholar] [CrossRef]
- Mazar, Y.; Cytryn, E.; Erel, Y.; Rudich, Y. Effect of dust storms on the atmospheric microbiome in the Eastern Mediterranean. Environ. Sci. Technol. 2016, 50, 4194–4202. [Google Scholar] [CrossRef]
- Gat, D.; Mazar, Y.; Cytryn, E.; Rudich, Y. Origin-dependent variations in the atmospheric microbiome community in Eastern Mediterranean dust storms. Environ. Sci. Technol. 2017, 51, 6709–6718. [Google Scholar] [CrossRef]
- Lynch, P.; Reid, J.S.; Westphal, D.L.; Zhang, J.; Hogan, T.F.; Hyer, E.J.; Curtis, C.A.; Hegg, D.A.; Shi, Y.; Campbell, J.R.; et al. An 11-year global gridded aerosol optical thickness reanalysis (v1.0) for atmospheric and climate sciences. Geosci. Model Dev. 2016, 9, 1489–1522. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.S.; Manche, L.; Xu, R.M.; Krainer, A.R. hnRNP A1 associates with telomere ends and stimulates telomerase activity. RNA 2006, 12, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyer, E.J.; Reid, J.S.; Zhang, J. An over-land aerosol optical depth data set for data assimilation by filtering, correction, and aggregation of MODIS Collection 5 optical depth retrievals. Atmos. Meas. Tech. 2011, 4, 379–408. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhang, J.; Reid, J.S.; Holben, B.; Hyer, E.J.; Curtis, C. An analysis of the collection 5 MODIS over-ocean aerosol optical depth product for its implication in aerosol assimilation. Atmos. Chem. Phys. 2011, 11, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Chen, A.L.-W.; Zhang, M.; Watson, J.; Chow, J.; Cao, J.; Chen, H.; Wang, W.; Zhang, J.; Zhan, C.; et al. Bioaerosol Concentrations and Size Distributions during the Autumn and Winter Seasons in an Industrial City of Central China. Aerosol Air Qual. Res. 2019, 19, 1095–1104. [Google Scholar] [CrossRef]
- Griffin, D.W.; Westphal, D.L.; Gray, M.A. Airborne microorganisms in the African desert dust corridor over the mid-Atlantic ridge, Ocean Drilling Program, Leg 209. Aerobiologia 2006, 22, 211. [Google Scholar] [CrossRef]
- Draxler, R.R. HYSPLIT_4 User’s Guide, NOAA Technical Memorandum ERL ARL-230; NOAA Air Resources Laboratory: Silver Spring, MD, USA, 1999; Volume 35.
- Clauß, M.; Springoru, A.C.; Hartung, J. Comparison of Different Fluorescence and Non-Fluorescence Staining Techniques for Rapid Detection of Airborne Micro-Organisms Collected on Room Temperature Vulcanizing (RTV) Silicones from Generated Aerosols and from Ambient Air. Aerosol Sci. Technol. 2012, 46, 818–827. [Google Scholar] [CrossRef]
- Mayol, E.; Jiménez, M.A.; Herndl, G.J.; Duarte, C.M.; Arrieta, J.M. Resolving the abundance and air-sea fluxes of airborne microorganisms in the North Atlantic Ocean. Front. Microbial. 2014, 5, 557. [Google Scholar] [CrossRef]
- Massana, R.; Murray, A.E.; Preston, C.M.; Delong, E. Vertical distribution and phylogenetic characterization of marine planktonic Archaea in the Santa Barbara Channel. Appl. Environ. Microbiol. 1997, 63, 50–56. [Google Scholar] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Martin, M. CUTADAPT removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, G.J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Huneeus, N.; Schulz, M.; Balkanski, Y.; Griesfeller, J.; Prospero, J.; Kinne, S.; Bauer, S.l.; Boucher, O.; Chin, M.; Dentener, F. Global dust model intercomparison in AeroCom phase I. Atmos. Chem. Phys. 2011, 11, 7781–7816. [Google Scholar] [CrossRef] [Green Version]
- Xian, P.; Reid, J.S.; Hyer, E.J.; Sampson, C.R.; Rubin, J.I.; Ades, M.; Asencio, N.; Basart, S.; Benedetti, A.; Bhattacharjee, P.S. Current State of the global operational aerosol multi-model ensemble: An update from the International Cooperative for Aerosol Prediction (ICAP). Q. J. R. Meteorol. Soc. 2019, 1–34. [Google Scholar] [CrossRef]
- Cho, B.C.; Hwang, C.Y. Prokaryotic abundance and 16S rRNA gene sequences detected in marine aerosols on the East Sea (Korea). FEMS Microbiol. Ecol. 2011, 76, 327–341. [Google Scholar] [CrossRef]
- Yahya, R.Z.; Arrieta, M.J.; Cusack, M.; Duarte, C.M. Airborne Prokaryote and Virus Abundance Over the Red Sea. Front. Microbiol. 2019, 10, 1112. [Google Scholar] [CrossRef]
- Xia, X.; Wang, J.; Ji, J.; Zhang, J.; Chen, L.; Zhang, R. Bacterial Communities in Marine Aerosols Revealed by 454 Pyrosequencing of the 16S rRNA Gene. J. Atmos. Sci. 2015, 72, 2997–3008. [Google Scholar] [CrossRef]
- Rahav, E.; Shun-Yan, C.; Cui, G.; Liu, H.; Tsagakari, T.M.; Giannakourou, A.; Tsiola, A.; Psarra, S.; Lagaria, A.; Mulholland, M.R.; et al. Evaluating the impact of atmospheric depositions on springtime dinitrogen fixation in the Cretan Sea (Eastern Mediterranean)—A mesocosm approach. Front. Mar. Sci. 2016, 3, 1–13. [Google Scholar] [CrossRef]
- Rahav, E.; Paytan, A.; Chien, C.-T.; Ovadia, G.; Katz, T.; Herut, B. The impact of atmospheric dry deposition associated microbes on the southeastern Mediterranean Sea surface water following an intense dust storm. Front. Mar. Sci. 2016, 3, 127. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Kort, R.; Schulte, A.; Lazarus, D.; Schnetger, B.; Brumsack, H.J.; Broughton, W.J.; Favet, J. Life in Darwin’s dust: Intercontinental transport and survival of microbes in the nineteenth century. Environ. Microbiol. 2007, 9, 2911–2922. [Google Scholar] [CrossRef]
- Tang, K.; Huang, Z.; Huang, J.; Maki, T.L.; Zhang, S.; Shimizu, A.; Ma, X.; Shi, J.; Bi, J.; Zhou, T.; et al. Characterization of atmospheric bioaerosols along the transport pathway of Asian dust during the Dust-Bioaerosol 2016 Campaign. Atmos. Chem. Phys. 2018, 18, 7131–7148. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Lighthart, B. The Annual Bacterial Particle Concentration and Size Distribution in the Ambient Atmosphere in a Rural Area of the Willamette Valley, Oregon. Aerosol Sci. Technol. 2010, 3, 393–403. [Google Scholar] [CrossRef]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Martire, M.L.; Greco, S.; Facchini, M.C.; Rinaldi, M.; O’Dowd, C.; Ceburnis, D.; Danovaro, R. Transfer of labile organic matter and microbes from the ocean surface to the marine aerosol: An experimental approach. Sci. Rep. 2017, 7, 11475. [Google Scholar] [CrossRef]
- Peter, H.; Hörtnagl, P.; Reche, I.; Sommaruga, R. Bacterial diversity and composition during rain events with and without Saharan dust influence reaching a high mountain lake in the Alps. Environ. Microbiol. Rep. 2014, 6, 618–624. [Google Scholar] [CrossRef]
- Sharoni, S.; Trainic, M.; Schatz, D.; Lehahn, Y.; Flores, M.J.; Bidle, K.D.; Ben-Dor, S.; Rudich, Y.; Koren, I.; Vardi, A. Infection of phytoplankton by aerosolized marine viruses. Proc. Natl. Acad. Sci. USA 2015, 112, 6643–6647. [Google Scholar] [CrossRef] [Green Version]
- Cunliffe, M.; Upstill-Goddard, R.C.; Murrell, J.C. Microbiology of aquatic surface microlayers. FEMS Microbiol. Rev. 2010, 35, 233–246. [Google Scholar] [CrossRef]
- Liss, S.N.; Bicho, P.; Saddler, J. Microbiology and biodegradation of resin acids in pulp mill effluents: A minireview. Can. J. Microbiol. 1997, 43, 599–611. [Google Scholar] [CrossRef]
- Fields, N.D.; Oxborrow, G.S.; Puleo, J.R.; Herring, C.M. Evaluation of membrane filter field monitors for microbiologial air sampling. Appl. Microbiol. 1974, 27, 517–520. [Google Scholar]
- Jensen, P.A.; Todd, W.F.; Davis, G.N.; Scarpino, P.V. Evaluation of eight bioaersol samplers challenged with aerosols of free bacteria. Am. Ind. Hyg. Assoc. J. 1992, 53, 660–667. [Google Scholar] [CrossRef]
- Kesavan, J.; Schepers, D.; McFarland, A.R. Sampling and Retention Efficiencies of Batch-Type Liquid-Based Bioaerosol Samplers. Aerosol Sci. Technol. 2010, 44, 817–829. [Google Scholar] [CrossRef]
- Grinshpun, S.A.; Willeke, K.; Ulevicius, V.; Donnelly, J.; Lin, X.; Mainelis, G. Collection of Airborne Microorganisms: Advantages and Disadvantages of Different Methods. J. Aerosol Sci. 1996, 27 (Suppl. 1), S247–S248. [Google Scholar] [CrossRef]
- Mainelis, G.; Grinshpun, S.A.; Willeke, K.; Reponen, T.; Ulevicius, V.; Hintz, P. Collection of Airborne Microorganisms by Electrostatic Precipitation. Aerosol Sci. Tech. 1999, 30, 127–144. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Guillaume, J.B.; Duchaine, C. Bioaerosol Sampler Choice Should Consider Efficiency and Ability of Samplers to Cover Microbial Diversity. Appl. Environ. Microbiol. 2018, 84, e01589-18. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Region | Airmass Origin | Distance from Land (km) | Total AOD | Aluminum (ng m−3 air) | Observed OTUs | Shannon’s Index (H) |
|---|---|---|---|---|---|---|---|
| 1 | Eastern | Z2 | 155 | 0.282 | 569 | 241 | 7.48 |
| 2 | Eastern | Z3 | 154 | 0.285 | 516 | 158 | 6.76 |
| 3 | Eastern | Z3 | 102 | 0.22 | 331 | 100 | 6.06 |
| 4 | Eastern | Z3 | 204 | 0.224 | 410 | 141 | 6.41 |
| 5 | Eastern | Z1 | 156 | 0.196 | 205 | 119 | 5.94 |
| 6 | Central | Z1 | 78 | 0.233 | 220 | 95 | 5.75 |
| 7 | Central | Z1 | 213 | 0.225 | 661 | 96 | 6.03 |
| 8 | Central | Z1 | 288 | 0.133 | 404 | 92 | 5.89 |
| 9 | Central | Z2 | 48 | 0.18 | 134 | 66 | 5.17 |
| 10 | Central | Z2 | 112 | 0.115 | 41 | 76 | 5.64 |
| 11 | Central | Z1 | 49 | 0.147 | 104 | 96 | 6.04 |
| 12 | Western | Z4 | 135 | 0.359 | 196 | 164 | 6.83 |
| 13 | Western | Z4 | 126 | 0.252 | 172 | 82 | 5.83 |
| 14 | Western | Z4 | 56 | 0.134 | 395 | 70 | 5.7 |
| 15 | Western | Z4 | 36 | 0.2 | 355 | 108 | 6.37 |
| 16 | Western | Z1 | 42 | 0.11 | 103 | 75 | 5.79 |
| Common Bacteria | Open Ocean Studies | Med Coastal Studies | Med Seawater Samples | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phyla | Class | Order | Family | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 6 |
| Actinobacteria | Actinobacteria | Actinomycetales | Actinomycetaceae | × | |||||||||||||||
| Actinobacteria | Actinobacteria | Actinomycetales | Corynebacteriaceae | × | × | × | |||||||||||||
| Actinobacteria | Actinobacteria | Bifidobacteriales | Bifidobacteriaceae | × | |||||||||||||||
| Bacteroidetes | Flavobacteriia | Flavobacteriales | Weeksellaceae | × | |||||||||||||||
| Bacteroidetes | Saprospirae | Saprospirales | Chitinophagaceae | × | × | × | |||||||||||||
| Firmicutes | Bacilli | Bacillales | Bacillaceae | × | × | × | × | × | × | × | × | × | × | × | × | × | |||
| Firmicutes | Bacilli | Bacillales | Paenibacillaceae | × | × | ||||||||||||||
| Firmicutes | Bacilli | Bacillales | Planococcaceae | × | × | × | |||||||||||||
| Firmicutes | Bacilli | Bacillales | Staphylococcaceae | × | × | × | × | ||||||||||||
| Firmicutes | Bacilli | Turicibacterales | Turicibacteraceae | × | |||||||||||||||
| Firmicutes | Clostridia | Clostridiales | Clostridiaceae | × | × | × | |||||||||||||
| Firmicutes | Clostridia | Clostridiales | Peptostreptococcaceae | ||||||||||||||||
| Firmicutes | Clostridia | Clostridiales | Veillonellaceae | ||||||||||||||||
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Bradyrhizobiaceae | × | × | × | × | × | × | × | |||||||||
| Proteobacteria | Alphaproteobacteria | Rhodobacterales | Rhodobacteraceae | × | × | × | × | × | × | × | × | × | × | ||||||
| Proteobacteria | Alphaproteobacteria | Rhodospirillales | Rhodospirillaceae | × | × | × | × | × | × | × | |||||||||
| Proteobacteria | Alphaproteobacteria | Sphingomonadales | Sphingomonadaceae | × | × | × | × | × | × | × | × | ||||||||
| Proteobacteria | Betaproteobacteria | Burkholderiales | Comamonadaceae | × | × | × | × | × | × | × | |||||||||
| Proteobacteria | Gammaproteobacteria | Enterobacteriales | Enterobacteriaceae | × | × | × | × | × | × | × | × | × | |||||||
| Proteobacteria | Gammaproteobacteria | Pseudomonadales | Moraxellaceae | × | × | × | × | × | |||||||||||
| Proteobacteria | Gammaproteobacteria | Pseudomonadales | Pseudomonadaceae | × | × | × | × | × | × | × | × | × | |||||||
| Proteobacteria | Gammaproteobacteria | Salinisphaerales | Salinisphaeraceae | × | |||||||||||||||
| Proteobacteria | Gammaproteobacteria | Vibrionales | Vibrionaceae | × | × | × | × | × | × | ||||||||||
| Proteobacteria | Gammaproteobacteria | Xanthomonadales | Sinobacteraceae | × | |||||||||||||||
| Deinococcus-Thermus | Deinococci | Thermales | Thermaceae | × | |||||||||||||||
| Rare Bacteria | Open Ocean Studies | Med Coastal Studies | Med Seawater Studies | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phyla | Class | Order | Family | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | 6 |
| Actinobacteria | Actinobacteria | Actinomycetales | Dietziaceae | × | |||||||||||||||
| Actinobacteria | Actinobacteria | Actinomycetales | Micrococcaceae | × | × | × | × | × | × | ||||||||||
| Actinobacteria | Rubrobacteria | Rubrobacterales | Rubrobacteraceae | × | × | ||||||||||||||
| Bacteroidetes | BME43 | Unassigned | Unassigned | ||||||||||||||||
| Bacteroidetes | Bacteroidia | Bacteroidales | Porphyromonadaceae | ||||||||||||||||
| Bacteroidetes | Cytophagia | Cytophagales | Amoebophilaceae | × | |||||||||||||||
| Bacteroidetes | Saprospirae | Saprospirales | Undefined | ||||||||||||||||
| Bacteroidetes | Saprospirae | Saprospirales | Saprospiraceae | ||||||||||||||||
| Chlamydiae | Chlamydiia | Chlamydiales | Undefined | ||||||||||||||||
| Chlamydiae | Chlamydiia | Chlamydiales | Rhabdochlamydiaceae | ||||||||||||||||
| Cyanobacteria | 4C0d2 | MLE112 | Unassigned | × | × | × | |||||||||||||
| Firmicutes | Bacilli | Bacillales | Unassigned | × | × | × | × | ||||||||||||
| Firmicutes | Bacilli | Bacillales | Alicyclobacillaceae | × | |||||||||||||||
| Firmicutes | Bacilli | Bacillales | Thermoactinomycetaceae | × | |||||||||||||||
| Firmicutes | Bacilli | Bacillales | Exiguobacteraceae | × | |||||||||||||||
| Firmicutes | Bacilli | Gemellales | Gemellaceae | × | × | ||||||||||||||
| Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | × | |||||||||||||||
| Firmicutes | Bacilli | Lactobacillales | Streptococcaceae | × | × | ||||||||||||||
| Firmicutes | Clostridia | Clostridiales | Tissierellaceae | ||||||||||||||||
| Proteobacteria | Undefined | Unassigned | Unassigned | ||||||||||||||||
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Hyphomicrobiaceae | × | |||||||||||||||
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Methylobacteriaceae | × | |||||||||||||||
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Methylocystaceae | × | |||||||||||||||
| Proteobacteria | Alphaproteobacteria | Rhizobiales | Phyllobacteriaceae | × | × | × | × | × | × | × | |||||||||
| Proteobacteria | Alphaproteobacteria | Rhodobacterales | Hyphomonadaceae | × | × | ||||||||||||||
| Proteobacteria | Betaproteobacteria | Burkholderiales | Oxalobacteraceae | × | × | × | × | × | × | ||||||||||
| Proteobacteria | Gammaproteobacteria | Alteromonadales | Alteromonadaceae | × | × | × | × | × | × | × | × | ||||||||
| Proteobacteria | Gammaproteobacteria | Oceanospirillales | Halomonadaceae | × | × | ||||||||||||||
| Proteobacteria | Gammaproteobacteria | Pasteurellales | Pasteurellaceae | × | |||||||||||||||
| Spirochaetes | Spirochaetes | Spirochaetales | Spirochaetaceae | ||||||||||||||||
| WPS | Unassigned | Unassigned | Unassigned | ||||||||||||||||
| Deinococcus-Thermus | Deinococci | Deinococcales | Deinococcaceae | ||||||||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mescioglu, E.; Rahav, E.; Belkin, N.; Xian, P.; Eizenga, J.M.; Vichik, A.; Herut, B.; Paytan, A. Aerosol Microbiome over the Mediterranean Sea Diversity and Abundance. Atmosphere 2019, 10, 440. https://doi.org/10.3390/atmos10080440
Mescioglu E, Rahav E, Belkin N, Xian P, Eizenga JM, Vichik A, Herut B, Paytan A. Aerosol Microbiome over the Mediterranean Sea Diversity and Abundance. Atmosphere. 2019; 10(8):440. https://doi.org/10.3390/atmos10080440
Chicago/Turabian StyleMescioglu, Esra, Eyal Rahav, Natalia Belkin, Peng Xian, Jordan M. Eizenga, Ania Vichik, Barak Herut, and Adina Paytan. 2019. "Aerosol Microbiome over the Mediterranean Sea Diversity and Abundance" Atmosphere 10, no. 8: 440. https://doi.org/10.3390/atmos10080440
APA StyleMescioglu, E., Rahav, E., Belkin, N., Xian, P., Eizenga, J. M., Vichik, A., Herut, B., & Paytan, A. (2019). Aerosol Microbiome over the Mediterranean Sea Diversity and Abundance. Atmosphere, 10(8), 440. https://doi.org/10.3390/atmos10080440