Environmental Drivers for Cambial Reactivation of Qilian Junipers (Juniperus przewalskii) in a Semi-Arid Region of Northwestern China




Abstract
:1. Introduction
2. Materials and Methods
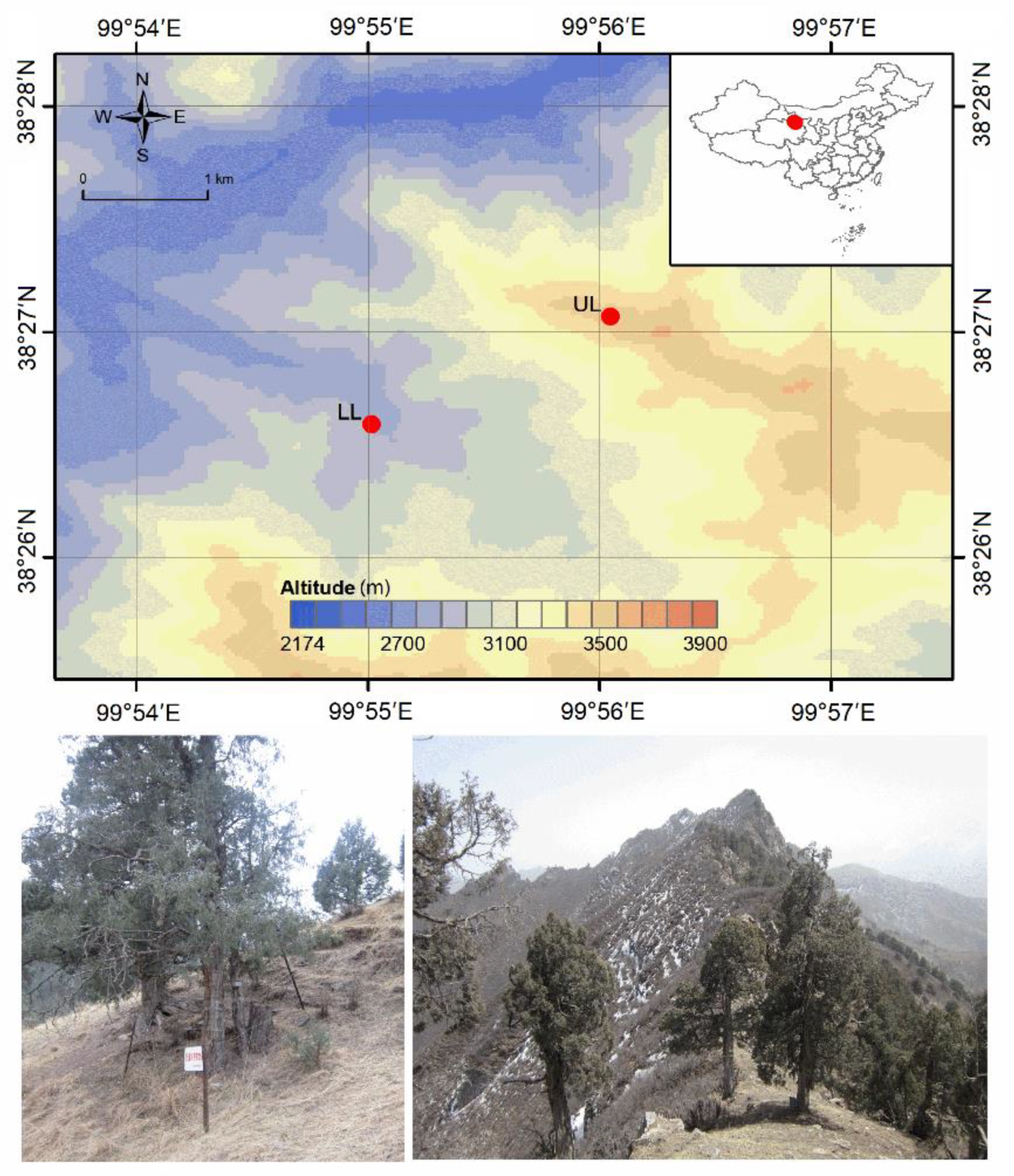
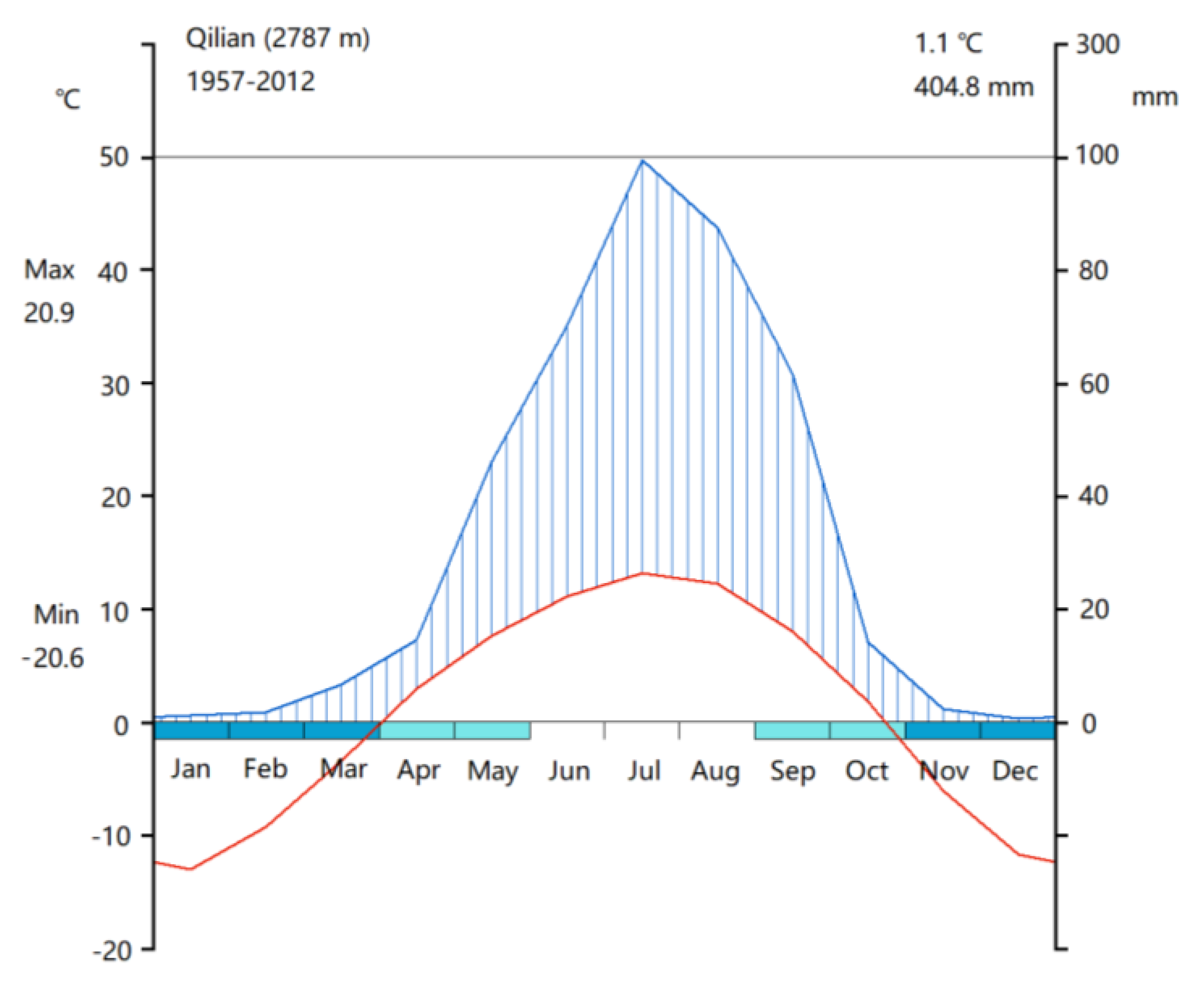
2.1. Study Sites and Tree Selection
2.2. Meteorological Data
2.3. Xylem Sampling and Observation
2.4. Statistical Analyses
3. Results
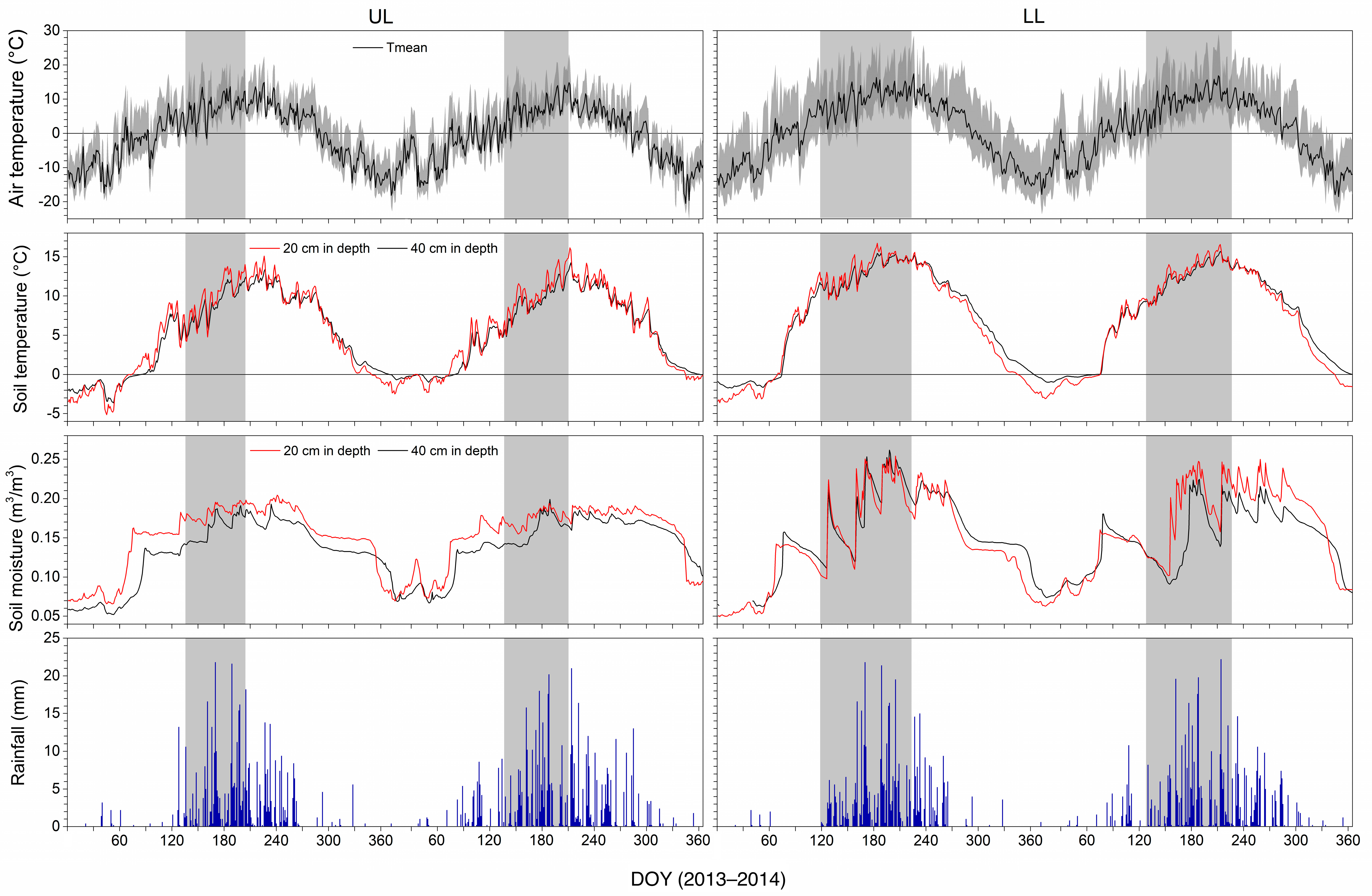
3.1. Weather during 2013 and 2014
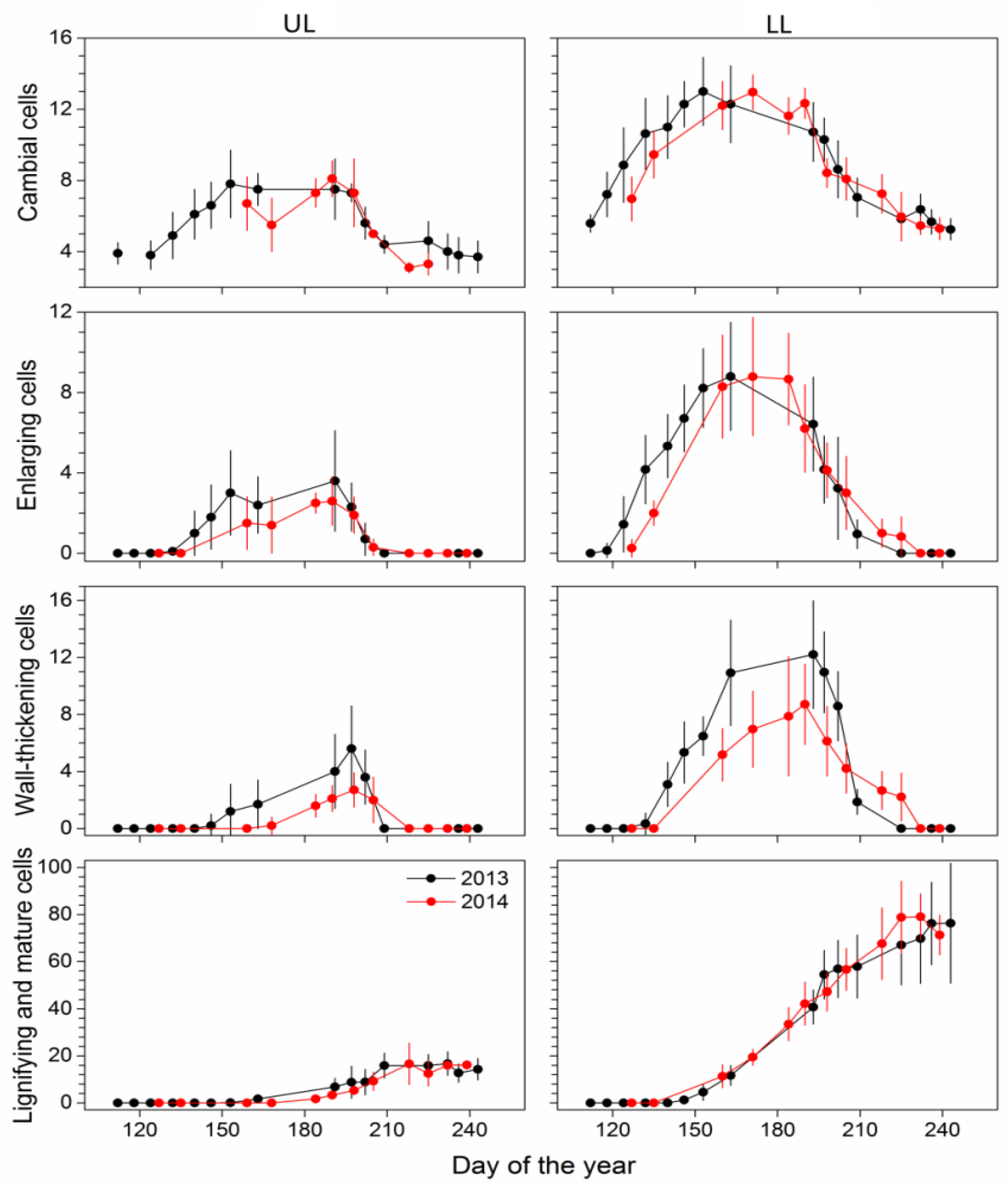
3.2. Cambial Activity
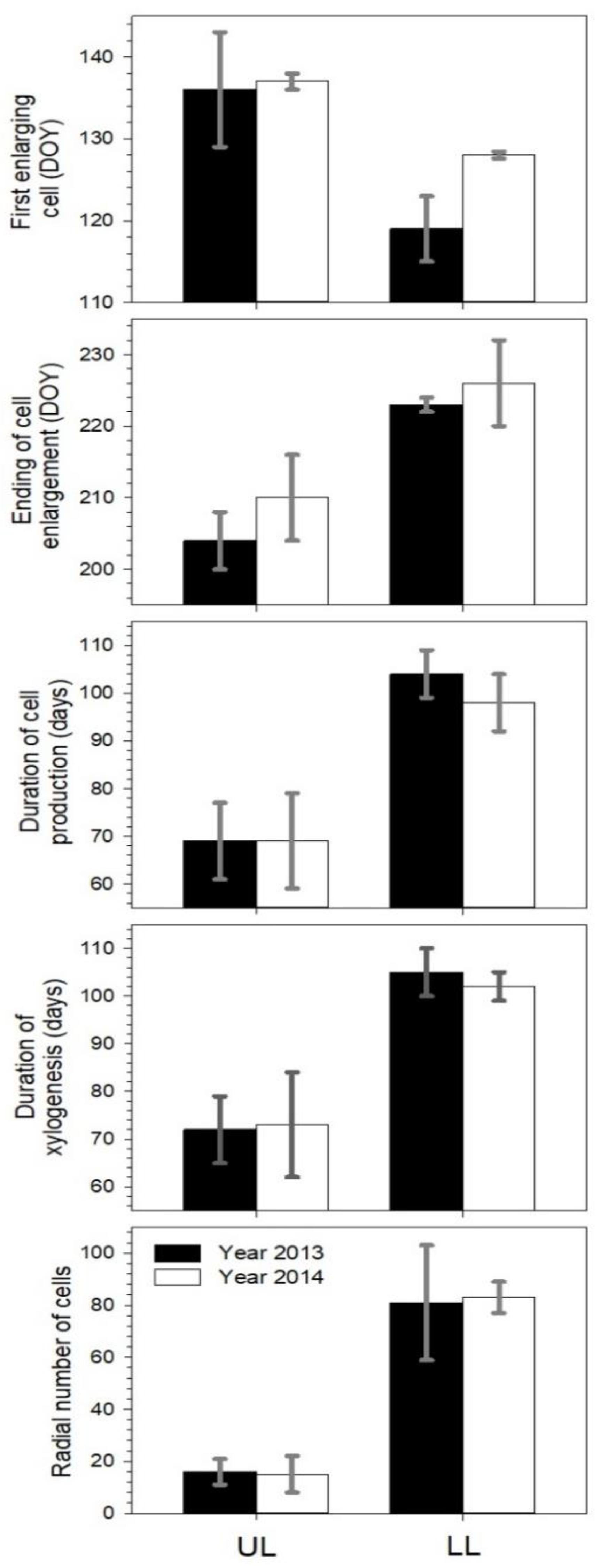
3.3. Xylem Differentiation and Cell Production
4. Discussion
4.1. Impact of Climatic Factors on Cambial Reactivation
4.2. Phenology and Cell Production
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Morin, H.; Deslauriers, A. Multi-scale influence of snowmelt on xylogenesis of black spruce. Arct. Antarct. Alp. Res. 2011, 43, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Ziaco, E.; Biondi, F.; Rossi, S.; Deslauriers, A. Environmental drivers of cambial phenology in Great Basin bristlecone pine. Tree Physiol. 2016, 36, 818–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuine, I. Why does phenology drive species distribution? Philos. Trans. R. Soc. Lond. Ser. B 2010, 365, 3149–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; Huang, J.G.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere. Glob. Chang. Biol. 2016, 22, 3804–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6. [Google Scholar] [CrossRef]
- Begum, S.; Nakaba, S.; Oribe, Y.; Kubo, T.; Funada, R. Induction of cambial reactivation by localized heating in a deciduous hardwood hybrid poplar (Populus sieboldii x P. grandidentata). Ann. Bot. 2007, 100, 439–447. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Gričar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Čufar, K.; Cuny, H.; Deslauriers, A.; Fonti, P.; Gričar, J.; et al. Chilling and forcing temperatures interact to predict the onset of wood formation in Northern Hemisphere conifers. Glob. Chang. Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Nakaba, S.; Oribe, Y.; Kubo, T.; Funada, R. Cambial sensitivity to rising temperatures by natural condition and artificial heating from late winter to early spring in the evergreen conifer Cryptomeria japonica. Trees 2010, 24, 43–52. [Google Scholar] [CrossRef]
- Gričar, J.; Zupančič, M.; Čufar, K.; Koch, G.; Schmitt, U.; Oven, P. Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Ann. Bot. 2006, 97, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, P.; Rossi, S.; Camarero, J.J.; Ellison, A.M.; Liang, E.; Peñuelas, J. Critical temperature and precipitation thresholds for the onset of xylogenesis of Juniperus przewalskii in a semi-arid area of the north-eastern Tibetan Plateau. Ann. Bot. 2018, 121, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Ziaco, E.; Truettner, C.; Biondi, F.; Bullock, S. Moisture-driven xylogenesis in Pinus ponderosa from a Mojave Desert mountain reveals high phenological plasticity. Plant Cell Environ. 2018, 41, 823–836. [Google Scholar] [CrossRef]
- Abe, H.; Nakai, T. Effect of the water status within a tree on tracheid morphogenesis in Cryptomeria japonica D. Don. Trees 1999, 14, 124–129. [Google Scholar] [CrossRef]
- Butto, V.; Rossi, S.; Deslauriers, A.; Morin, H. Is size an issue of time? Relationship between the duration of xylem development and cell traits. Ann. Bot. 2019, 123, 1257–1265. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Pederson, N.; Zhang, F.; Niu, H.; Zhao, S.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018, 38, 840–852. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agr. Forest Meteorol. 2018, 260–261, 165–175. [Google Scholar] [CrossRef]
- Yang, B.; He, M.; Shishov, V.; Tychkov, I.; Vaganov, E.; Rossi, S.; Ljungqvist, F.C.; Brauning, A.; Griessinger, J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc. Natl. Acad. Sci. USA 2017, 114, 6966–6971. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Balducci, L.; Ren, P.; Rossi, S. Xylogenesis and moisture stress. In Secondary Xylem Biology: Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 45–58. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Gričar, J.; Liang, E.; Čufar, K. Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau? Ann. Bot. 2015, 115, 629–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Wang, J.; Liu, J. A 1556-year-length early summer moisture reconstruction for the Hexi Corridor, Northwestern China. Sci. China Earth Sci. 2019, 62, 953–963. [Google Scholar] [CrossRef]
- Yang, B.; Qin, C.; Shi, F.; Sonechkin, D.M. Tree ring-based annual streamflow reconstruction for the Heihe River in arid northwestern China from AD 575 and its implications for water resource management. Holocene 2011, 22, 773–784. [Google Scholar] [CrossRef]
- Gou, X.; Zhou, F.; Zhang, Y.; Chen, Q.; Zhang, J. Forward modeling analysis of regional scale tree-ring patterns around the northeastern Tibetan Plateau, Northwest China. Biogeosci. Disc. 2013, 10, 9969–9988. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Qin, C.; Wang, J.; He, M.; Melvin, T.M.; Osborn, T.J.; Briffa, K.R. A 3500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2014, 111, 2903–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yang, B.; Deslauriers, A.; Qin, C.; He, M.; Shi, F.; Liu, J. Two phases of seasonal stem radius variations of Sabina przewalskii Kom. in northwestern China inferred from sub-diurnal shrinkage and expansion patterns. Trees 2012, 26, 1747–1757. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Yang, B. Comparing meteorological records between mountainous and valley bottom sites in the upper reaches of the Heihe River, northwestern China: Implications for dendroclimatology. Theor. Appl. Climatol. 2016, 128, 407–419. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T. Assessment of cambial activity and xylogenesis by microsampling tree species: An example at the alpine timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Oladi, R.; Pourtahmasi, K.; Eckstein, D.; Bräuning, A. Seasonal dynamics of wood formation in Oriental beech (Fagus orientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest, Iran. Trees 2011, 25, 425–433. [Google Scholar] [CrossRef]
- Rossi, S.; Girard, M.J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Chang. Biol. 2014, 20, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Saeri, S.; Pourtahmasi, K.; Oladi, R.; Rathgeber, C. Wood formation in Juniperus excelsa ssp. polycarpos in the high mountains of North-East Iran. J. Trop. For. Sci. 2013, 25, 421–428. [Google Scholar]
- Deslauriers, A.; Morin, H.; Bégin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. For. Res. 2003, 33, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Oribe, Y.; Funada, R. Regulation of cambial activity in relation to environmental conditions: Understanding the role of temperature in wood formation of trees. Physiol. Plant. 2013, 147, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Rossi, S.; Simard, S.; Rathgeber, C.B.K.; Deslauriers, A.; De Zan, C. Effects of a 20-day-long dry period on cambial and apical meristem growth in Abies balsamea seedlings. Trees 2009, 23, 85–93. [Google Scholar] [CrossRef]
- Turcotte, A.; Morin, H.; Krause, C.; Deslauriers, A.; Thibeault-Martel, M. The timing of spring rehydration and its relation with the onset of wood formation in black spruce. Agric. For. Meteorol. 2009, 149, 1403–1409. [Google Scholar] [CrossRef]
- Bernal, M.; Estiarte, M.; Peñuelas, J. Drought advances spring growth phenology of the Mediterranean shrub Erica multiflora. Plant Biol. 2011, 13, 252–257. [Google Scholar] [CrossRef]
- Prieto, P.; Peñuelas, J.; Niinemets, Ü.; Ogaya, R.; Schmidt, I.; Beier, C.; Tietema, A.; Sowerby, A.; Emmett, B.A.; Kovács Láng, E.; et al. Changes in the onset of spring growth in shrubland species in response to experimental warming along a north–south gradient in Europe. Glob. Ecol. Biogeogr. 2009, 18, 473–484. [Google Scholar] [CrossRef]
- Oribe, Y.; Funada, R.; Shibagaki, M.; Kubo, T. Cambial reactivation in locally heated stems of the evergreen conifer Abies sachalinensis (Schmidt) Masters. Planta 2001, 212, 684–691. [Google Scholar] [CrossRef]
- Gričar, J.; Prislan, P.; Gryc, V.; Vavrčík, H.; de Luis, M.; Čufar, K. Plastic and locally adapted phenology in cambial seasonality and production of xylem and phloem cells in Picea abies from temperate environments. Tree Physiol. 2014, 34, 869–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaganov, E.A.; Hughes, M.K.; Kirdyanov, A.V.; Schweingruber, F.H.; Silkin, P.P. Influence of snowfall and melt timing on tree growth in subarctic Eurasia. Nature 1999, 400, 149–151. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, H. Condensated water and its ecological environment effect in northwestern of China. Acta Geosci. Sin. 1999, 20, 762–766. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Air Temperature (°C) | Soil Temperature (°C) | Soil Moisture (m3/m3) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Site | Mean | Maximum | Minimum | At 20 cm | At 40 cm | At 20 cm | At 40 cm | Rainfall (mm) |
| 2013 | UL | −0.3 (7.6) | 6.1 (13.2) | −4.5 (3.7) | 5.3 (10.4) | 5.0 (9.5) | 0.15 (0.18) | 0.13 (0.16) | 500.4 (420.2) |
| 2014 | −0.9 (6.5) | 5.3 (12.3) | −5.2 (2.5) | 5.5 (10.5) | 5.3 (9.6) | 0.15 (0.18) | 0.14 (0.16) | 584.8 (411.8) | |
| 2013 | LL | 1.5 (10.5) | 10.9 (18.5) | −4.8 (4.6) | 7.0 (13.6) | 7.4 (13.2) | 0.14 (0.19) | 0.16 (0.19) | 505.3 (431.8) |
| 2014 | 1.0 (9.6) | 10.2 (17.9) | −5.2 (3.4) | 6.8 (13.0) | 7.2 (12.6) | 0.16 (0.18) | 0.14 (0.16) | 549.6 (388.0) | |
| Site | Year | Site × Year | ||||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| First enlarging cell | 34.10 | <0.0001 | 4.96 | 0.038 | 2.98 | 0.100 |
| Ending of enlargement | 56.95 | <0.0001 | 3.99 | 0.060 | 0.31 | 0.585 |
| Duration of cell production | 67.00 | <0.0001 | 0.39 | 0.539 | 0.62 | 0.441 |
| Duration of xylogenesis | 68.20 | <0.0001 | 0.11 | 0.739 | 0.22 | 0.641 |
| Radial number of cells | 170.37 | <0.0001 | 0.01 | 0.916 | 0.21 | 0.655 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Q.; Rossi, S.; Yang, B.; Qin, C.; Li, G. Environmental Drivers for Cambial Reactivation of Qilian Junipers (Juniperus przewalskii) in a Semi-Arid Region of Northwestern China. Atmosphere 2020, 11, 232. https://doi.org/10.3390/atmos11030232
Zeng Q, Rossi S, Yang B, Qin C, Li G. Environmental Drivers for Cambial Reactivation of Qilian Junipers (Juniperus przewalskii) in a Semi-Arid Region of Northwestern China. Atmosphere. 2020; 11(3):232. https://doi.org/10.3390/atmos11030232
Chicago/Turabian StyleZeng, Qiao, Sergio Rossi, Bao Yang, Chun Qin, and Gang Li. 2020. "Environmental Drivers for Cambial Reactivation of Qilian Junipers (Juniperus przewalskii) in a Semi-Arid Region of Northwestern China" Atmosphere 11, no. 3: 232. https://doi.org/10.3390/atmos11030232
APA StyleZeng, Q., Rossi, S., Yang, B., Qin, C., & Li, G. (2020). Environmental Drivers for Cambial Reactivation of Qilian Junipers (Juniperus przewalskii) in a Semi-Arid Region of Northwestern China. Atmosphere, 11(3), 232. https://doi.org/10.3390/atmos11030232