

Emission of Greenhouse Gases and Ammonia from the Excreta of Nellore Bulls Submitted to Energy and Tannin Supplementation

 ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Experimental Site
2.2. Treatments and Pasture Management
2.3. Experiment 1—Quantification of N2O and CH4 Emissions
2.4. Experiment 2—Ammonia Volatilization Assessment
2.5. Statistical Analyses
3. Results and Discussions
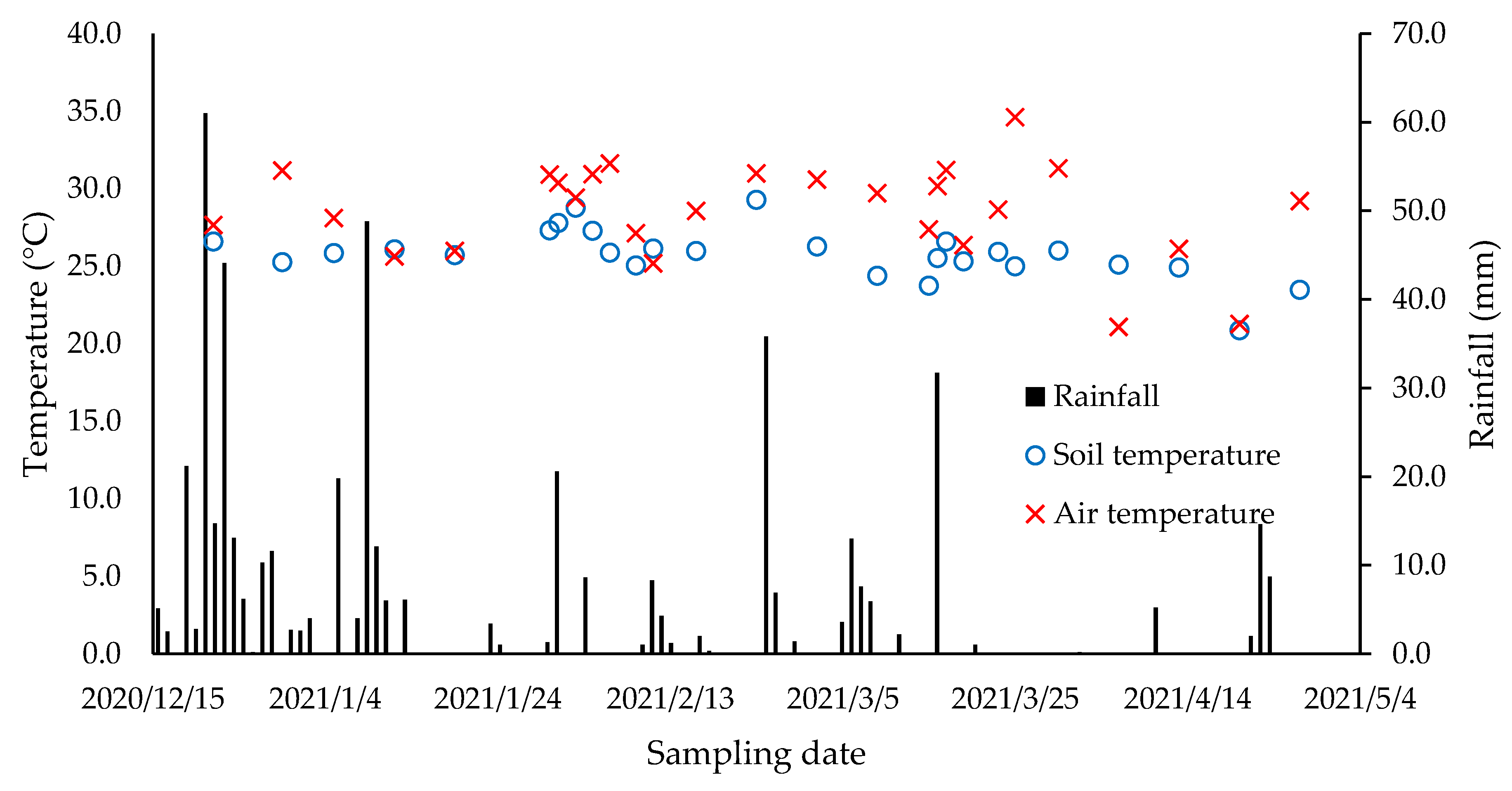
3.1. Rainfall and Temperature Patterns
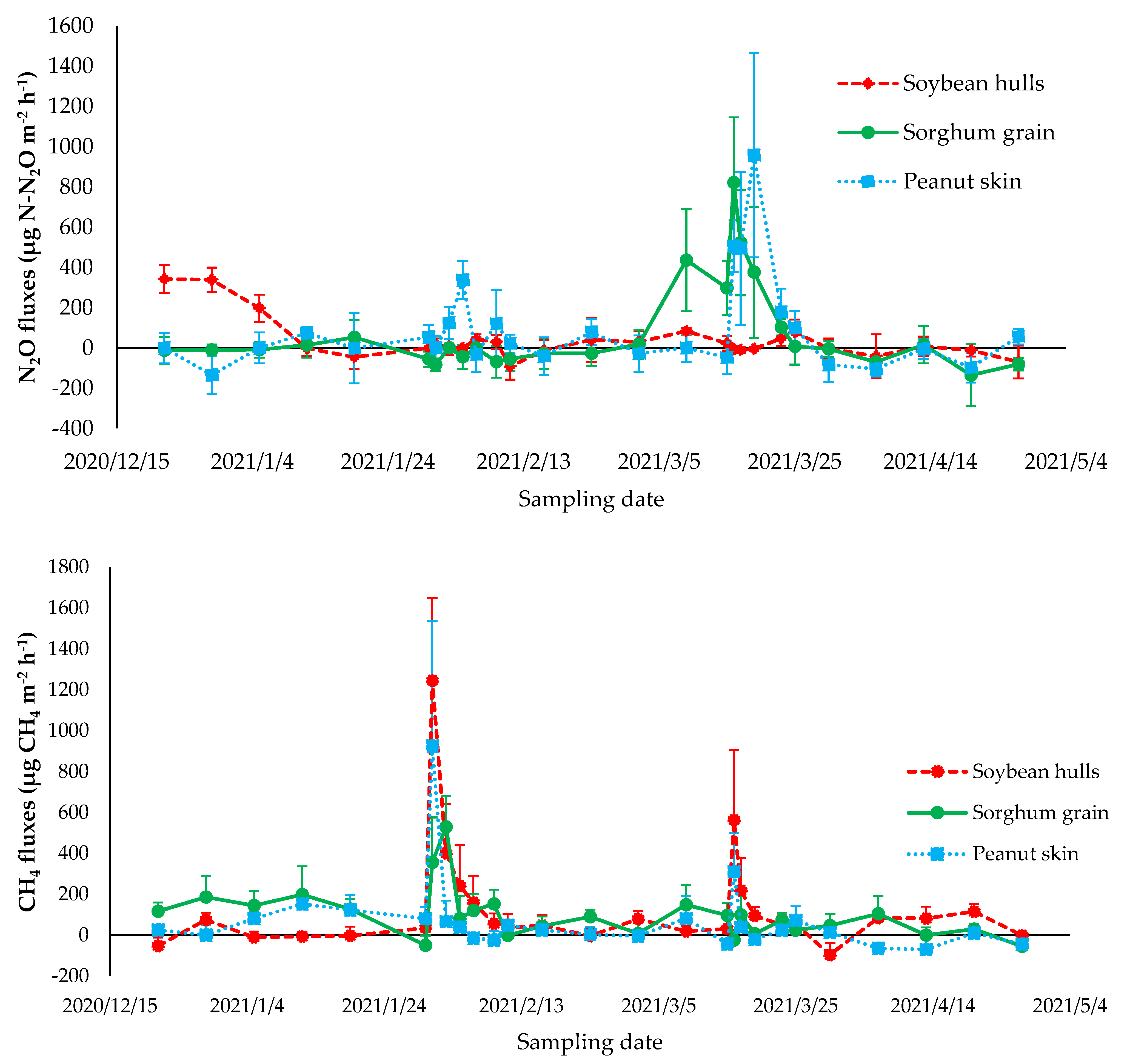
3.2. Greenhouse Gas Emissions
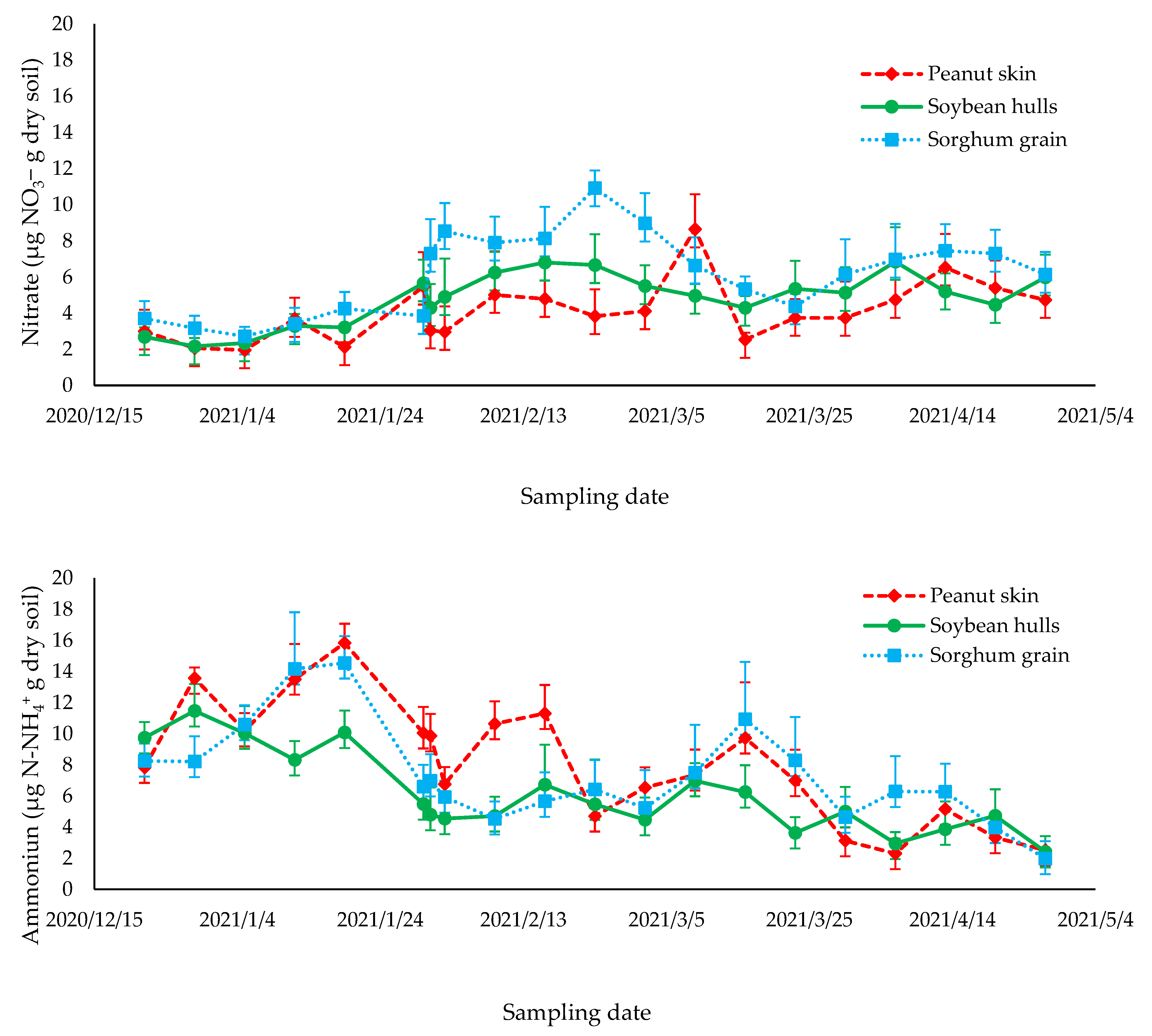
3.3. Ammonia Volatilization
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Silva, H.M.; Dubeux, J.C., Jr.; Silveira, M.L.; Lira, M.A., Jr.; Cardoso, A.S.; Vendramini, J.M. Greenhouse gas mitigation and carbon sequestration potential in humid grassland ecosystems in Brazil: A review. J. Environ. Manag. 2022, 323, 116269. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Barbero, R.P.; Romanzini, E.P.; Teobaldo, R.W.; Ongaratto, F.; Fernandes, M.H.M.R.; Ruggieri, A.C.; Reis, R.A. Intensification: A key strategy to achieve great animal and environmental beef cattle production sustainability in Brachiaria grasslands. Sustainability 2020, 12, 6656. [Google Scholar] [CrossRef]
- Berça, A.S.; Tedeschi, L.O.; Cardoso, A.S.; Reis, R.A. Meta-analysis of the Relationship Between Dietary Condensed Tannins and Methane Emissions by Cattle. Anim. Feed Sci. Technol. 2023, 298, 115564. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change—IPCC. 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories. 2019. Available online: https://www.ipcc.ch/report/2019-refinement-to-the-2006-ipcc-guidelines-for-nationalgreenhouse-gas-inventories (accessed on 1 May 2023).
- Aluwong, T.; Wuyep, P.A.; Allam, L. Livestock environment interactions: Methane emissions from ruminants. Afr. J. Biotechnol. 2011, 10, 1265–1269. [Google Scholar]
- Cardoso, A.S.; Berndt, A.; Leytem, A.; Alves, B.J.; de Carvalho, I.D.N.; Soares, L.H.B.; Urquiaga, S.; Boddey, R.M. Impact of the intensification of beef production in Brazil on greenhouse gas emissions and land use. Agric. Syst. 2016, 143, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Grossi, G.; Goglio, P.; Vitali, A.; Williams, A.G. Livestock and climate change: Impact of livestock on climate and mitigation strategies. Anim. Front. 2019, 9, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, A.S.; Alari, F.O.; de Andrade, N.; Nascimento, T.S.; van Cleef, F.D.O.S.; Alves, B.J.R.; Ruggieri, A.C.; Malheiros, E.B. Nitrous oxide emissions in silvopastoral systems: Key driving variables and seasonality. Agric. For. Meteorol. 2022, 316, 108851. [Google Scholar] [CrossRef]
- Guimarães, B.C.; Gomes, K.F.; Homem, B.G.; Lima, I.B.G.; Spasiani, P.P.; Boddey, R.M.; Alves, B.J.R.; Casagrande, D.R. Emissions of N2O and NH3 from cattle excreta in grass pastures fertilized with N or mixed with a forage vegetable. Nutr. Cycl. Agroecosystems 2022, 1, 22. [Google Scholar]
- Carvalho, G.M.; Brito, L.D.F.; Coelho, L.D.M.; Cardoso, A.S.; Messana, J.D.; Colovate, P.H.D.; Ganga, M.J.G.; da Cruz, M.C.P.; Malheiros, E.B.; Berchielli, T.T. Ammonia, nitrous oxide and methane emissions from the excreta of cattle receiving rumen undegradable protein. Agron. J. 2022, 114, 1655–1671. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Firestone, M.K.; Davidson, E.A. Microbiological basis of NO and N2O production and consumption in soil. In Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere; Andreae, M.O., Schimel, D.S., Eds.; John Wiley and Sons: New York, NY, USA, 1998; pp. 7–21. [Google Scholar]
- Cardoso, A.S.; Brito, L.F.; Janusckiewicz, E.R.; Morgado, E.S.; Barbero, R.P.; Koscheck, J.F.W.; Reis, R.A.; Ruggieri, A.C. Impact of grazing intensity and seasons on greenhouse gas emissions in a tropical grassland. Agric. Syst. 2017, 20, 845–859. [Google Scholar] [CrossRef]
- Coelho, L.M.; Brito, L.F.; Messana, J.D.; Cardoso, A.S.; Carvalho, G.M.; Torres, R.Z.S.; Carlos, R.S.; Malheiros, E.B.; da Cruz, M.C.P.; Berchielli, T.T. Effects of rumen undegradable protein sources on nitrous oxide, methane and ammonia emission from the manure of feedlot-finished cattle. Sci. Rep. 2022, 12, 9166. [Google Scholar] [CrossRef]
- Ferrari, A.C.; Leite, R.G.; Fonseca, N.V.B.; Romanzini, E.P.; Cardoso, A.S.; Barbero, R.P.; Costa, D.F.A.; Ruggieri, A.C.; Reis, R.A. Performance, nutrient use, and methanogenesis of Nellore cattle on a continuous grazing system of Urochloa brizantha and types fed supplement varying on protein and energy sources. Livest. Sci. 2021, 253, 104716. [Google Scholar] [CrossRef]
- Delevatti, L.M.; Romanzini, E.P.; Koscheck, J.F.W.; Araújo, T.L.R.; Renesto, D.M.; Ferrari, A.C.; Barbero, R.P.; Mulliniks, J.T.; Reis, R.A. Forage management intensification and supplementation strategy: Intake and metabolic parameters on beef cattle production. Anim. Feed Sci. Technol. 2019, 247, 74–82. [Google Scholar] [CrossRef]
- Leite, R.G.; Cardoso, A.S.; Fonseca, N.V.B.; Silva, M.L.C.; Tedeschi, L.O.; Delevatti, L.M.; Ruggieri, A.C.; Reis, R.A. Effects of nitrogen fertilization on protein and carbohydrate fractions of Marandu palisade grass. Sci. Rep. 2021, 11, 14786. [Google Scholar] [CrossRef]
- D’aurea, A.P.; Cardoso, A.S.; Guimarães, Y.S.R.; Fernandes, L.B.; Ferreira, L.E.; Reis, R.A. Mitigating Greenhouse Gas Emissions from Beef Cattle Production in Brazil through Animal Management. Sustainability 2021, 13, 7207. [Google Scholar] [CrossRef]
- Siniscalchi, D.; Cardoso, A.S.; Corrêa, D.C.D.C.; Ferreira, M.R.; Andrade, M.E.B.; da Cruz, L.H.G.; Ruggieri, A.C.; Reis, R.A. Effects of condensed tannins on greenhouse gas emissions and nitrogen dynamics from urine-treated grassland soil. Environ. Sci. Pollut. Res. 2022, 29, 85026–85035. [Google Scholar] [CrossRef]
- Adamczyk, B.; Simon, J.; Kitunen, V.; Adamczyk, S.; Smolander, A. Tannins and their complex interaction with different organic nitrogen compounds and enzymes: Old paradigms versus recent advances. Chem. Open 2017, 6, 610–614. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; McGinn, S.M.; Martinez, T.F.; McAllister, T.A. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef] [Green Version]
- Teobaldo, R.W.; Cardoso, A.S.; Brito, T.R.; Leite, R.G.; Romanzini, E.P.; Granja-Salcedo, Y.T.; Reis, R.A. Response of Phytogenic Additives on Enteric Methane Emissions and Animal Performance of Nellore Bulls Raised in Grassland. Sustainability 2022, 14, 9395. [Google Scholar] [CrossRef]
- Ebert, P.J.; Bailey, E.A.; Shreck, A.L.; Jennings, J.S.; Cole, N.A. Effect of condensed tannin extract supplementation on growth performance, nitrogen balance, gas emissions, and energetic losses of beef steers. J. Anim. Sci. 2017, 95, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Avila, A.S.; Zambom, M.A.; Faccenda, A.; Fischer, M.L.; Anschau, F.A.; Venturini, T.; Tinini, R.C.R.; Dessbesell, J.G.; Faciola, A.P. Effects of black wattle (Acacia mearnsii) condensed tannins on intake, protozoa population, ruminal fermentation, and nutrient digestibility in Jersey steers. Animals 2020, 10, 1011. [Google Scholar] [CrossRef] [PubMed]
- Koenig, K.M.; Beauchemin, K.A.; McGinn, S.M. Feeding condensed tannins to mitigate ammonia emissions from beef feedlot cattle fed high-protein finishing diets containing distillers grains. J. Anim. Sci. 2018, 96, 4414–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.Y. Modulation of protein metabolism to mitigate nitrous oxide (N2O) emission from excreta of livestock. Curr. Protein Pept. Sci. 2017, 18, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Gemeda, B.S.; Hassen, A. Effect of Tannin and Species Variation on In vitro Digestibility, Gas, and Methane Production of Tropical Browse Plants. J. Anim. Sci. 2015, 28, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef] [Green Version]
- Longhini, V.Z.; Cardoso, A.S.; Berça, A.S.; Boddey, R.M.; Reis, R.A.; Dubeux Junior, J.C.B.; Ruggieri, A.C. Nitrogen supply and affect rainfall emissions from dairy cattle excreta and urea applied on warm-climate pastures. J. Environ. Qual. 2020, 49, 1453–1466. [Google Scholar] [CrossRef]
- Cardoso, A.S.; Oliveira, S.C.; Janusckiewicz, E.R.; Brito, L.F.; Silva, E.M.; Reis, R.A.; Ruggieri, A.C. Seasonal effects on, nitrous oxide, and methane emissions for beef cattle excreta and urea fertilizer applied to a tropical pasture. Soil Tillage Res. 2019, 194, 104–341. [Google Scholar] [CrossRef]
- Ruggieri, A.C.; Cardoso, A.S.; Ongaratto, F.; Casagrande, D.R.; Barbero, R.P.; Brito, L.D.F.; Azenha, M.V.; Oliveira, A.A.; Koscheck, J.F.W.; Reis, R.A. Grazing Intensity Impacts on Herbage Mass, Sward Structure, Greenhouse Gas Emissions, and Animal Performance: Analysis of Brachiaria Pastureland. Agronomy 2020, 10, 1750. [Google Scholar] [CrossRef]
- Van Raij, B.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recommendations for Fertilization and Liming for the State of São Paulo, 2nd ed.; Technical Bulletin, 100; IAC: Campinas, Brazil, 1997; 285p. [Google Scholar]
- Fonseca, N.V.B.; Cardoso, A.S.; Berça, A.S.; Dornellas, I.A.; Ongaratto, F.; Silva, M.L.C.; Ruggieri, A.C.; Reis, R.A. Effect of different nitrogen fertilizers on nitrogen efficiency use in Nellore bulls grazing on Marandu palisade grass. Livest. Sci. 2022, 263, 105012. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Barbero, R.P.; Malheiros, E.B.; Araújo, T.L.R.; Nave, R.L.G.; Mulliniks, J.T.; Berchielli, T.T.; Ruggieri, A.C.; Reis, R.A. Combining Marandu grazgrassing height and supplementation level to optimize growth and productivity of yearling bulls. Anim. Feed Sci. Technol. 2015, 209, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Koscheck, J.F.W.; Romanzini, E.P.; Barbero, R.P.; Delevatti, L.M.; Ferrari, A.C.; Mulliniks, J.T.; Mousquer, J.T.; Berchielli, T.T.; Reis, R.A. How do animal performance and methane emissions vary with forage management intensification and supplementation? Anim. Prod. Sci. 2020, 60, 1201–1209. [Google Scholar] [CrossRef]
- Cardoso, A.S.; José Neto, A.; Azenha, M.V.; Morgado, E.S.; Brito, L.D.F.; Janusckiewicz, E.R.; Reis, R.A.; Ruggieri, A.C. Mineral salt intake effects on faecal-N concentration and the volume and composition of beef cattle urine. Trop. Anim. Health Prod. 2019, 51, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Schofield, P.; Mbugua, D.M.; Pell, A.N. Analysis of condensed tannins: A review. Anim. Feed Sci. Technol. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists, 16th ed.; AOAC International: Arlington, TX, USA, 1995. [Google Scholar]
- Parkinson, J.A.; Allen, S.E. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological materials. Commun. Soil Sci. Plant Anal. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Forster, J.C. Soil sampling, handling, storage and analysis. In Methods in Applied Soil Microbiology and Biochemistry; Kassem, A., Paolo, N., Eds.; Academic Press: London, UK, 1995; pp. 49–121. [Google Scholar]
- Doane, T.A.; Horwáth, W.R. Spectrophotometric determination of nitrate with a single reagent. Anal. Lett. 2003, 36, 2713–2722. [Google Scholar] [CrossRef]
- Alves, B.J.R.; Smith, K.A.; Flores, R.A.; Cardoso, A.S.; Oliveira, W.R.D.; Jantalia, C.P.; Urquiaga, S.; Boddey, R.M. Selection of the most suitable sampling time for static chambers for the estimation of daily mean N2O flux from soils. Soil Biol. Biochem. 2012, 46, 129–135. [Google Scholar] [CrossRef]
- Van Der Weerden, T.J.; Luo, J.; Di, H.J.; Podolyan, A.; Phillips, R.L.; Saggar, S.; Klein, C.A.M.; Cox, N.; Ettema, P.; Rys, G. Nitrous oxide emissions from urea fertiliser and effluent with and without inhibitors applied to pasture. Agric. Ecosyst. Environ. 2016, 219, 58–70. [Google Scholar] [CrossRef]
- Araújo, E.S.; Marsola, T.; Miyazawa, M.; Soares, L.H.B.; Urquiaga, S.; Boddey, R.M.; Alves, B.J.R. Static semi-open chamber calibration for quantification of volatilized ammonia from the soil. Pesqui. Agropecu. Bras. 2009, 44, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, A.S.; Longhini, V.Z.; Berça, A.S.; Ongaratto, F.; Siniscalchi, D.; Reis, R.A.; Ruggieri, A.C. Pasture management and greenhouse gases emissions. Biosci. J. 2022, 64, 1–11. [Google Scholar] [CrossRef]
- Dobbie, K.E.; McTaggart, I.P.; Smith, K.A. Nitrous oxide emissions from intensive agricultural systems: Variations between crops and seasons, key driving variables, and mean emission factors. J. Geophys. Res. 1999, 104, 26891–26899. [Google Scholar] [CrossRef]
- Rivera, A.R.; Berchielli, T.T.; Messana, J.D.; Velasquez, P.T.; Franco, A.V.M.; Fernandes, L.B. Ruminal fermentation and methane production in cattle fed tifton 85 grass hay and concentrated with additives. Rev. Bras. Zootec. 2010, 39, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, A.S.; Junqueira, J.B.; Reis, R.A.; Ruggieri, A.C. How do greenhouse gas emissions vary with biofertilizer type and soil temperature and moisture in a tropical grassland? Pedosphere 2020, 30, 607–617. [Google Scholar] [CrossRef]
- Rowlings, D.W.; Grace, P.R.; Scheer, C.; Liu, S. Rainfall variability drives interannual variation in N2O emissions from a humid, subtropical pasture. Sci. Total Environ. 2015, 512, 8–18. [Google Scholar] [CrossRef]
- Calsamiglia, S.; Ferret, A.; Reynolds, C.K.; Kristensen, N.B.; Van Vuuren, A. Strategies for optimizing nitrogen use by ruminants. Animal 2010, 4, 1184–1196. [Google Scholar] [CrossRef]
- Yang, K.; Wei, C.; Zhao, G.Y.; Xu, Z.W.; Lin, S.X. Dietary supplementation of tannic acid modulates nitrogen excretion pattern and urinary nitrogenous constituents of beef cattle. Livest. Sci. 2016, 191, 148–152. [Google Scholar] [CrossRef]
- Berça, A.S.; Cardoso, A.S.; Longhini, V.Z.; Tedeschi, L.O.; Boddey, R.M.; Berndt, A.; Reis, R.A.; Ruggieri, A.C. Methane production and nitrogen balance of dairy heifers grazing palisade grass cv. Marandu alone or with forage peanut. J. Anim. Sci. 2019, 97, 4625–4634. [Google Scholar] [CrossRef]
- Bretas, I.L.; Paciullo, D.S.; Alves, B.J.R.; Martins, M.R.; Cardoso, A.S.; Lima, M.A.; Rodrigues, A.R.R.; Silva, F.F.; Chizzotti, F.H. Nitrous oxide, methane, and ammonia emissions from cattle excreta on Brachiaria decumbens growing in monoculture or silvopasture with Acacia mangium and Eucalyptus grandis. Agric. Ecosyst. Environ. 2020, 295, 106896. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | P resin | OM | pH | K | Ca | Mg | H + Al | BS | CEC | V |
|---|---|---|---|---|---|---|---|---|---|---|
| mg/dm3 | g/dm3 | % | ||||||||
| Peanut skin | 36 | 36 | 5.1 | 3.8 | 33 | 16 | 33 | 53 | 86 | 61 |
| Soybean Hull | 52 | 37 | 5.2 | 5.2 | 37 | 16 | 33 | 58 | 92 | 63 |
| Sorghum Grain | 18 | 33 | 4.8 | 2.3 | 24 | 13 | 36 | 39 | 75 | 52 |
| Forage | Peanut Skin | Soybean Hull | Sorghum Grain | |
|---|---|---|---|---|
| DM | 18.8 | 89.8 | 87.7 | 88.1 |
| CP | 13.8 | 18.4 | 11.5 | 11.4 |
| ADF | 34.6 | 38 | 48.1 | 7.5 |
| NDF | 67.9 | 40.9 | 65.7 | 16.2 |
| Starch | - | 4.4 | 4.9 | 52.9 |
| EE | 1.8 | 17.9 | 0.7 | 3.7 |
| MM | 8 | 9.2 | 1.6 | 2.7 |
| NFC | - | 13.7 | 20.5 | 66 |
| TDN | - | 94 | 66 | 84 |
| Lignin | 3.8 | - | - | - |
| DM dig | 73 | - | - | - |
| OM | 92 | - | - | - |
| Treatment | Tannins Concentration | Standard Deviation | |
|---|---|---|---|
| (mg Tannins/g DM) | (% Tannins/g DM) | ||
| Soybean hull | 1.69 | 0.17 | 0.01 |
| Sorghum grain | 30.34 | 3.03 | 0.03 |
| Peanut skin | 64.48 | 6.45 | 3.07 |
| Application | Treatments | Dung | Urine | |||
|---|---|---|---|---|---|---|
| DM (%) | N (%) | C (%) | pH | N (g/L) | ||
| First | Peanut skin | 14.5 | 3.31 | 45.5 | 9.26 | 2.00 |
| First | Soybean hull | 18.0 | 1.77 | 29.0 | 8.54 | 2.35 |
| First | Sorghum grain | 16.5 | 2.38 | 35.5 | 8.73 | 1.26 |
| Second | Peanut skin | 14.0 | 2.10 | 32.5 | 8.76 | 0.84 |
| Second | Soybean hull | 15.0 | 1.77 | 29.0 | 9.00 | 0.84 |
| Second | Sorghum grain | 16.0 | 2.38 | 35.5 | 8.63 | 0.84 |
| CH4 | N2O | |
|---|---|---|
| (mg CH4 m2) | (mg N2O m2) | |
| Soybean hull | 272.7 a | 116.3 a |
| Sorghum grains | 146.5 a | 145.4 a |
| Peanut skin | 245.5 a | 149.2 a |
| Standard error of the mean | 37.7 | 32.5 |
| p-value | 0.3599 | 0.9116 |
| Excreta Type | |||
|---|---|---|---|
| Supplement Type | Dung | Dung + Urine | Urine |
| Soybean hull | 2.47 | 4.42 | 13.55 |
| Sorghum grains | 2.34 | 4.32 | 14.14 |
| Peanut skin | 1.93 | 3.73 | 14.54 |
| Average | 2.25 A | 4.16 B | 14.08 C |
| SEM | 0.34 | 0.66 | 1.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rigobello, I.L.; Cardoso, A.d.S.; Fonseca, N.V.B.; Ongaratto, F.; Silva, M.M.; Bahia, A.S.R.d.S.; Dornellas, I.A.; Reis, R.A. Emission of Greenhouse Gases and Ammonia from the Excreta of Nellore Bulls Submitted to Energy and Tannin Supplementation. Atmosphere 2023, 14, 1112. https://doi.org/10.3390/atmos14071112
Rigobello IL, Cardoso AdS, Fonseca NVB, Ongaratto F, Silva MM, Bahia ASRdS, Dornellas IA, Reis RA. Emission of Greenhouse Gases and Ammonia from the Excreta of Nellore Bulls Submitted to Energy and Tannin Supplementation. Atmosphere. 2023; 14(7):1112. https://doi.org/10.3390/atmos14071112
Chicago/Turabian StyleRigobello, Izabela Larosa, Abmael da Silva Cardoso, Natalia Vilas Boas Fonseca, Fernando Ongaratto, Matheus Mello Silva, Angelica Santos Rabelo de Souza Bahia, Isadora Alves Dornellas, and Ricardo Andrade Reis. 2023. "Emission of Greenhouse Gases and Ammonia from the Excreta of Nellore Bulls Submitted to Energy and Tannin Supplementation" Atmosphere 14, no. 7: 1112. https://doi.org/10.3390/atmos14071112
APA StyleRigobello, I. L., Cardoso, A. d. S., Fonseca, N. V. B., Ongaratto, F., Silva, M. M., Bahia, A. S. R. d. S., Dornellas, I. A., & Reis, R. A. (2023). Emission of Greenhouse Gases and Ammonia from the Excreta of Nellore Bulls Submitted to Energy and Tannin Supplementation. Atmosphere, 14(7), 1112. https://doi.org/10.3390/atmos14071112