Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn

Abstract
:1. Introduction
2. Experiments
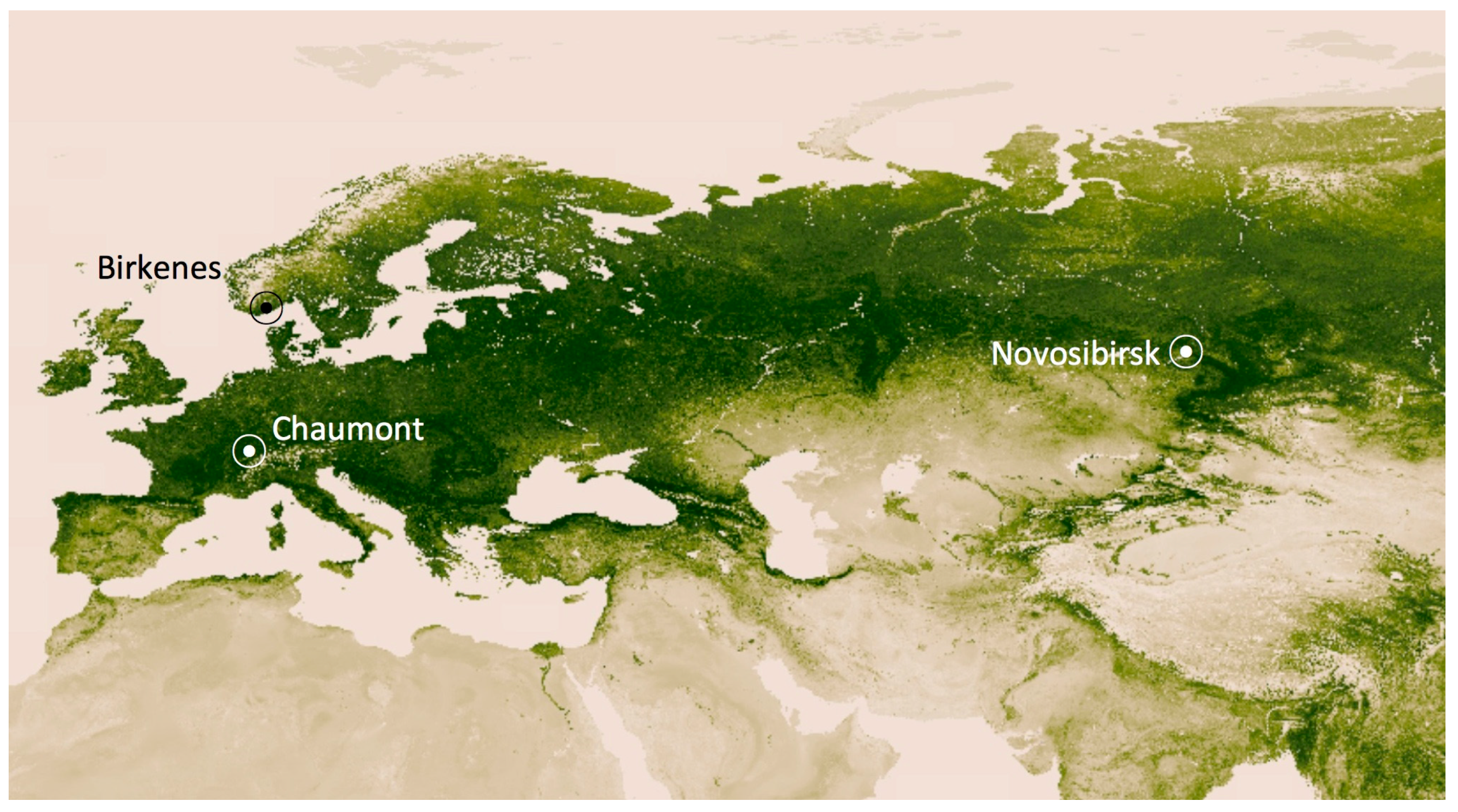
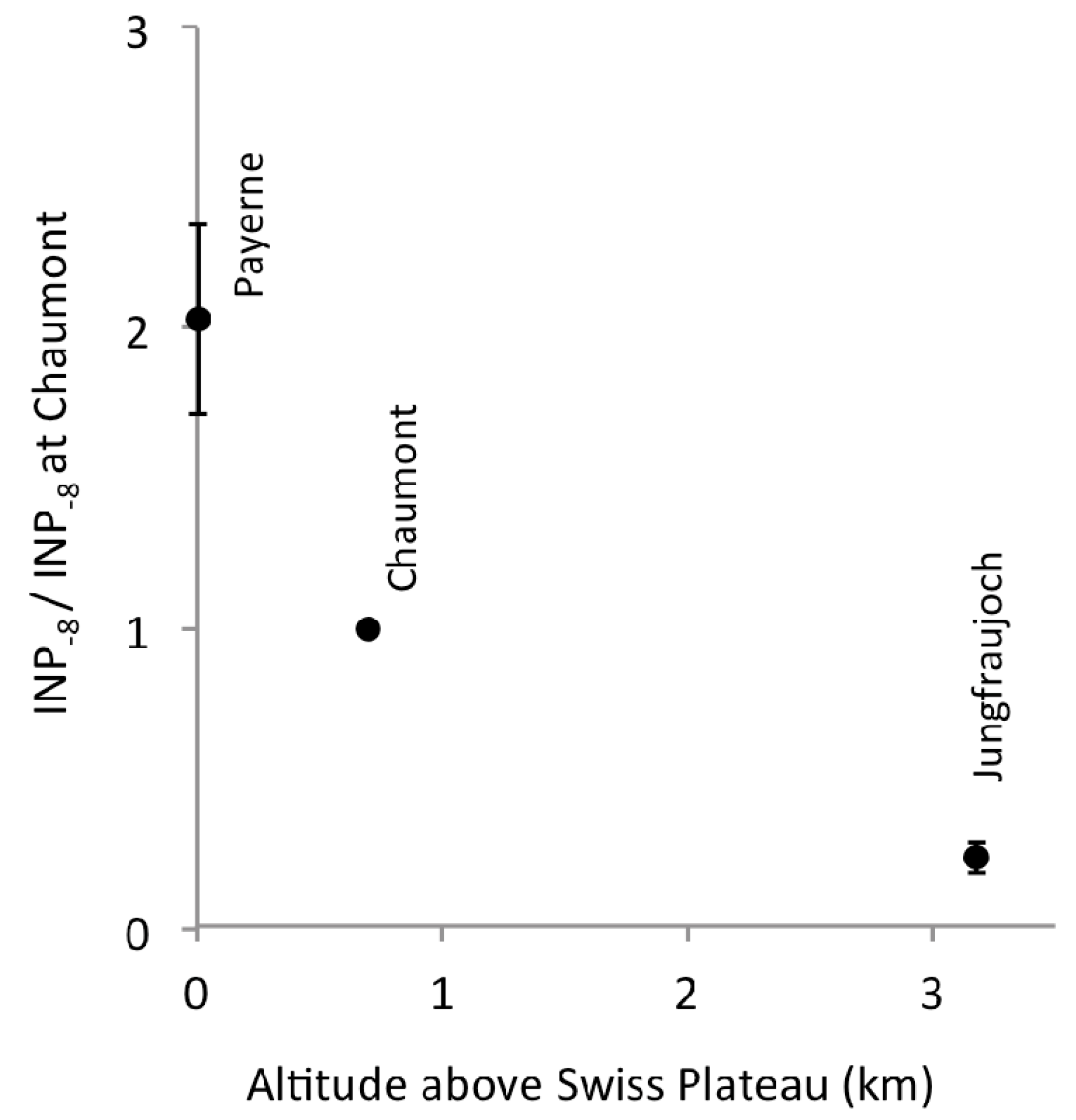
2.1. Altitudinal Gradient
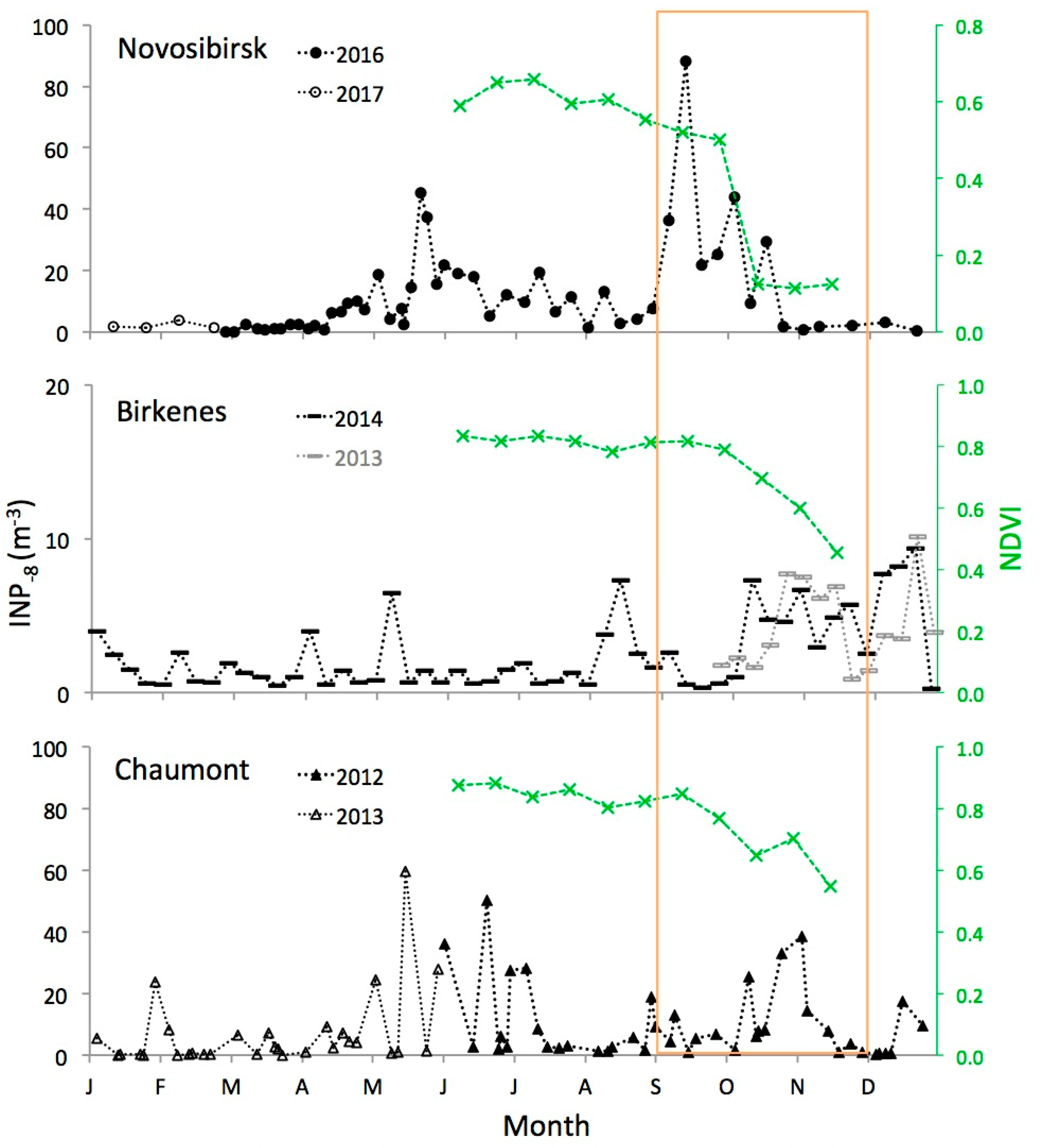
2.2. Time Series
3. Results and Discussion
3.1. Altitudinal Gradient
3.2. Autumn Peak
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maki, L.R.; Galyan, E.L.; Chien, M.C.; Caldwell, D.R. Ice nucleation induced by Pseudomonas syringae. Appl. Microbiol. 1974, 28, 456–459. [Google Scholar] [PubMed]
- Sands, D.C.; Langhans, V.E.; Scharen, A.L.; de Smet, G. The association between bacteria and rain and possible resultant meteorological implications. J. Hung. Meteorol. Serv. 1982, 86, 148–152. [Google Scholar]
- Morris, C.E.; Conen, F.; Huffman, J.A.; Phillips, V.; Pöschl, U.; Sands, D.C. Bioprecipitation: A feedback cycle linking Earth history, ecosystem dynamics and land use through biological ice nucleators in the atmosphere. Glob. Chang. Biol. 2014, 20, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Stopelli, E.; Conen, F.; Guilbaud, C.; Zopfi, J.; Alewell, C.; Morris, C.E. Ice nucleators, bacterial cells and Pseudomonas syringae in precipitation at Jungfraujoch. Biogeosciences 2017, 14, 1189–1196. [Google Scholar] [CrossRef]
- Desprès, V.R.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.S.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.O.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B 2012, 64, 15598. [Google Scholar] [CrossRef]
- Amato, P.; Joly, M.; Schaupp, C.; Attard, E.; Möhler, O.; Morris, C.E.; Brunet, Y.; Delort, A.M. Survival and ice nucleation activity of bacteria as aerosols in a cloud simulation chamber. Atmos. Chem. Phys. 2015, 15, 6455–6465. [Google Scholar] [CrossRef]
- Pummer, B.G.; Budke, C.; Augustin-Bauditz, S.; Niedermeier, D.; Felgitsch, L.; Kampf, C.J.; Huber, R.G.; Liedl, K.R.; Loerting, T.; Moschen, T.; et al. Ice nucleation by water-soluble macromolecules. Atmos. Chem. Phys. 2015, 15, 4077–4091. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. Atmospheric ice nuclei from decomposing vegetation. Nature 1972, 236, 163–165. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. World-wide source of leaf derived freezing nuclei. Nature 1973, 246, 212–213. [Google Scholar] [CrossRef]
- Thimonier, A.; Sedivy, I.; Schleppi, P. Estimating leaf area index in different types of mature forest stands in Switzerland: A comparison of methods. Eur. J. For. Res. 2010, 129, 543–562. [Google Scholar] [CrossRef]
- NOAA Environmental Visualisation Laboratory. Green: Vegetation on Our Planet. Available online: https://www.nnvl.noaa.gov/green.php (accessed on 22 May 2017).
- Conen, F.; Henne, S.; Morris, C.E.; Alewell, C. Atmospheric ice nucleators active ≥ −12 °C can be quantified on PM10 filters. Atmos. Meas. Tech. 2012, 5, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Conen, F.; Rodriguez, S.; Hüglin, C.; Henne, S.; Herrmann, E.; Bukowiecki, N.; Alewell, C. Atmospheric ice nuclei at the high-altitude observatory Jungfraujoch, Switzerland. Tellus B 2015, 67, 25014. [Google Scholar] [CrossRef]
- Federal Office of Topography Swisstopo. Landsat Mosaic. Available online: https://shop.swisstopo.admin.ch/en/products/images/ortho_images/landsat (accessed on 10 October 2017).
- Conen, F.; Eckhardt, S.; Gundersen, H.; Stohl, A.; Yttri, K.E. Rainfall drives atmospheric ice nucleating particles in the maritime climate of Southern Norway. Atmos. Chem. Phys. Discuss. 2017. [Google Scholar] [CrossRef]
- Lukasova, V.; Lang, M.; Skvarenina, J. Seasonal changes in NDVI in relation to phenological phases, LAI and PAI of beech forests. Balt. For. 2014, 20, 248–262. [Google Scholar]
- Nagai, S.; Inoue, T.; Ohtsuka, T.; Kobayashi, H.; Kurumado, K.; Moraoka, H.; Nasahara, K.N. Relationship between spatio-temporal characteristics of leaf-fall phenology and seasonal variations in near surface-and satellite-observed vegetation indices in a cool-temperate deciduous broad-leaved forest in Japan. Int. J. Remote Sens. 2014, 35, 3520–3536. [Google Scholar] [CrossRef]
- Twohy, C.H.; McMeeking, G.R.; DeMott, P.J.; McCluskey, C.S.; Hill, T.C.J.; Burrows, S.M.; Kulkarni, G.R.; Tanarhte, M.; Kafle, D.N.; Toohey, D.W. Abundance of fluorescent biological aerosol particles at temperatures conducive to the formation of mixed-phase and cirrus clouds. Atmos. Chem. Phys. 2016, 16, 8205–8225. [Google Scholar] [CrossRef]
- Wang, Q.; Adiku, S.; Tenhunen, J.; Granier, A. On the relationship of NDVI with leaf area index in a deciduous forest site. Remote Sens. Environ. 2005, 94, 244–255. [Google Scholar] [CrossRef]
- Myoung, B.; Choi, Y.S.; Hong, S.; Park, S.K. Inter- and intra-annual variability of vegetation in the northern hemisphere and its association with precursory meteorological factors. Glob. Biogeochem. Cycles 2013, 27, 31–42. [Google Scholar] [CrossRef]
- Staebler, R.M.; Fitzjarrald, D.R. Measuring canopy structure and the kinematics of subcanopy flows in two forrests. J. Appl. Meteorol. 2005, 44, 1161–1179. [Google Scholar] [CrossRef]
- Limpert, E.; Stahel, W.A.; Abbt, M. Log-normal distributions across the sciences: Keys and clues. BioScience 2001, 51, 341–352. [Google Scholar] [CrossRef]
- Limpert, E.; Burke, J.; Galan, C.; del Mar Trigo, M.; West, J.S.; Stahel, W.A. Data, not only in aerobiology: How normal is the normal distribution? Aerobiologia 2008, 24, 121–124. [Google Scholar] [CrossRef]
- Boose, Y.; Kanji, Z.A.; Kohn, M.; Sireau, B.; Zipori, A.; Crawford, I.; Lloyd, G.; Bukowiecki, N.; Herrmann, E.; Kupiszewski, P.; et al. Ice nucleating particle measurements at 241 K during winter months at 3580 m MSL in the Swiss Alps. J. Atmos. Sci. 2016, 73, 2203–2228. [Google Scholar] [CrossRef]
- Schrod, J.; Weber, D.; Drücke, J.; Kelehis, C.; Pikridas, M.; Ebert, M.; Cvetkovic, B.; Nickovic, S.; Marinou, E.; Baars, H.; et al. Ice nucleating particles over the Eastern Mediterranean measured by unmanned aircraft systems. Atmos. Chem. Phys. 2017, 17, 4817–4835. [Google Scholar] [CrossRef]
- Hande, L.B.; Engler, C.; Hoose, C.; Tegen, I. Seasonal variability of Saharan desert dust and ice nucleating particles over Europe. Atmos. Chem. Phys. 2015, 15, 4389–4397. [Google Scholar] [CrossRef]
- Kanitz, T.; Seifert, P.; Ansman, A.; Engelmann, R.; Althausen, D.; Casiccia, C.; Rohwer, E.G. Contrasting the impact of aerosol at northern and southern midlatitudes on heterogeneous ice formation. Geophys. Res. Lett. 2011, 38, L17802. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Novosibirsk | Chaumont | Birkenes |
|---|---|---|---|
| sampling period | 02.16–02.17 | 06.12–05.13 | 01.14–12.14 |
| (MM.YY) | |||
| season | number of samples | ||
| spring | 24 | 18 | 14 |
| summer | 14 | 18 | 13 |
| autumn | 11 | 18 | 13 |
| winter | 7 | 18 | 12 |
| median INP−8 (m−3) | |||
| spring | 3.4 | 3.4 | 1.0 |
| summer | 10.5 | 2.9 | 1.4 |
| autumn | 21.8 | 7.4 | 2.9 |
| winter | 1.6 | 0.4 | 2.0 |
| multiplicative standard deviation (s*) | |||
| spring | 4.6 | 3.9 | 1.8 |
| summer | 2.2 | 4.0 | 2.1 |
| autumn | 7.9 | 2.3 | 3.2 |
| winter | 2.1 | 8.2 | 4.5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conen, F.; Yakutin, M.V.; Yttri, K.E.; Hüglin, C. Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere 2017, 8, 202. https://doi.org/10.3390/atmos8100202
Conen F, Yakutin MV, Yttri KE, Hüglin C. Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere. 2017; 8(10):202. https://doi.org/10.3390/atmos8100202
Chicago/Turabian StyleConen, Franz, Mikhail V. Yakutin, Karl Espen Yttri, and Christoph Hüglin. 2017. "Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn" Atmosphere 8, no. 10: 202. https://doi.org/10.3390/atmos8100202
APA StyleConen, F., Yakutin, M. V., Yttri, K. E., & Hüglin, C. (2017). Ice Nucleating Particle Concentrations Increase When Leaves Fall in Autumn. Atmosphere, 8(10), 202. https://doi.org/10.3390/atmos8100202