Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems

Abstract
:1. Introduction
2. Methods
2.1. Study Site Description and Experimental Design
2.2. Gas Flux Measurements
2.3. Soil and Plant Measurements
2.4. Statistical Analyses
3. Results
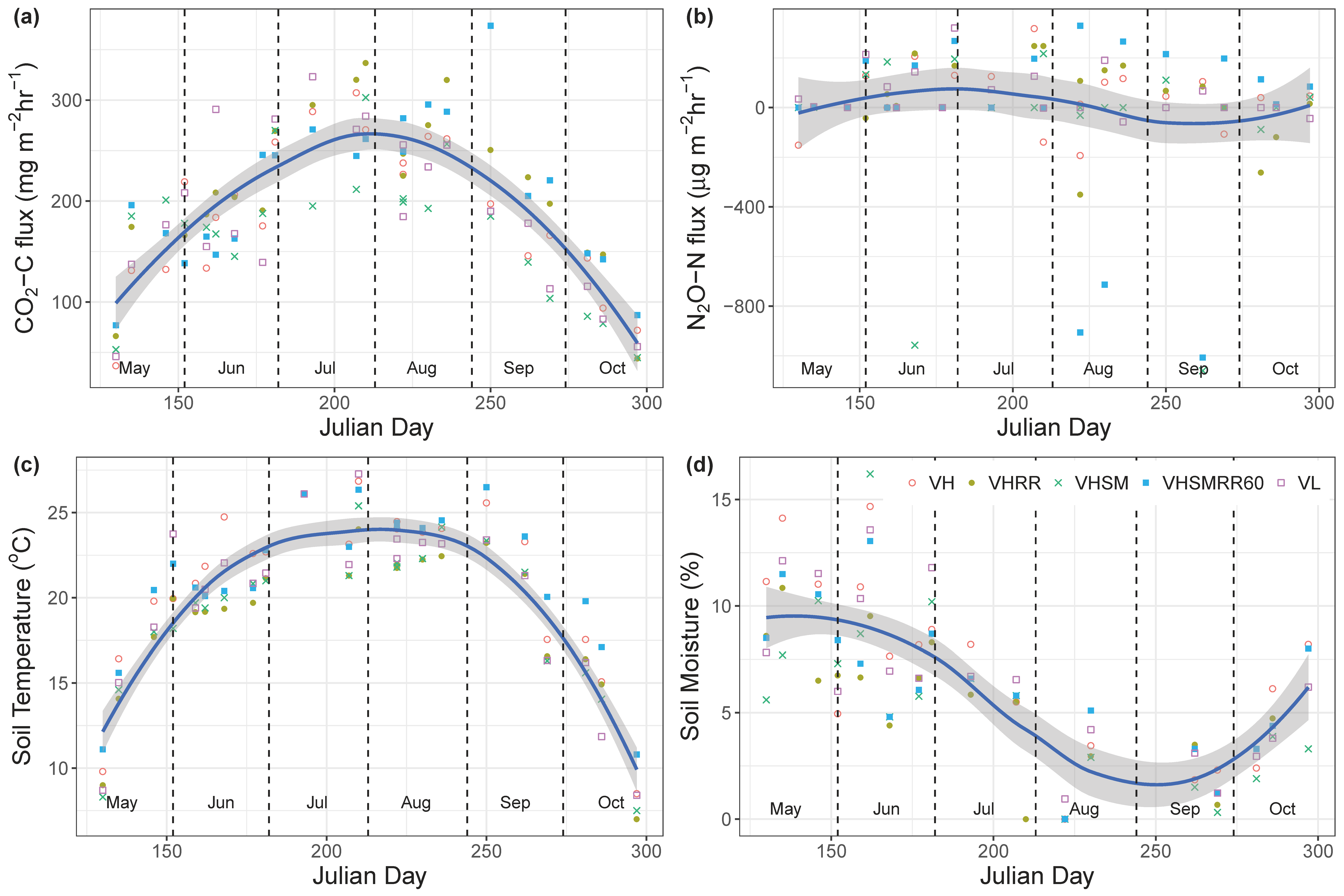
3.1. Gas Flux Analysis
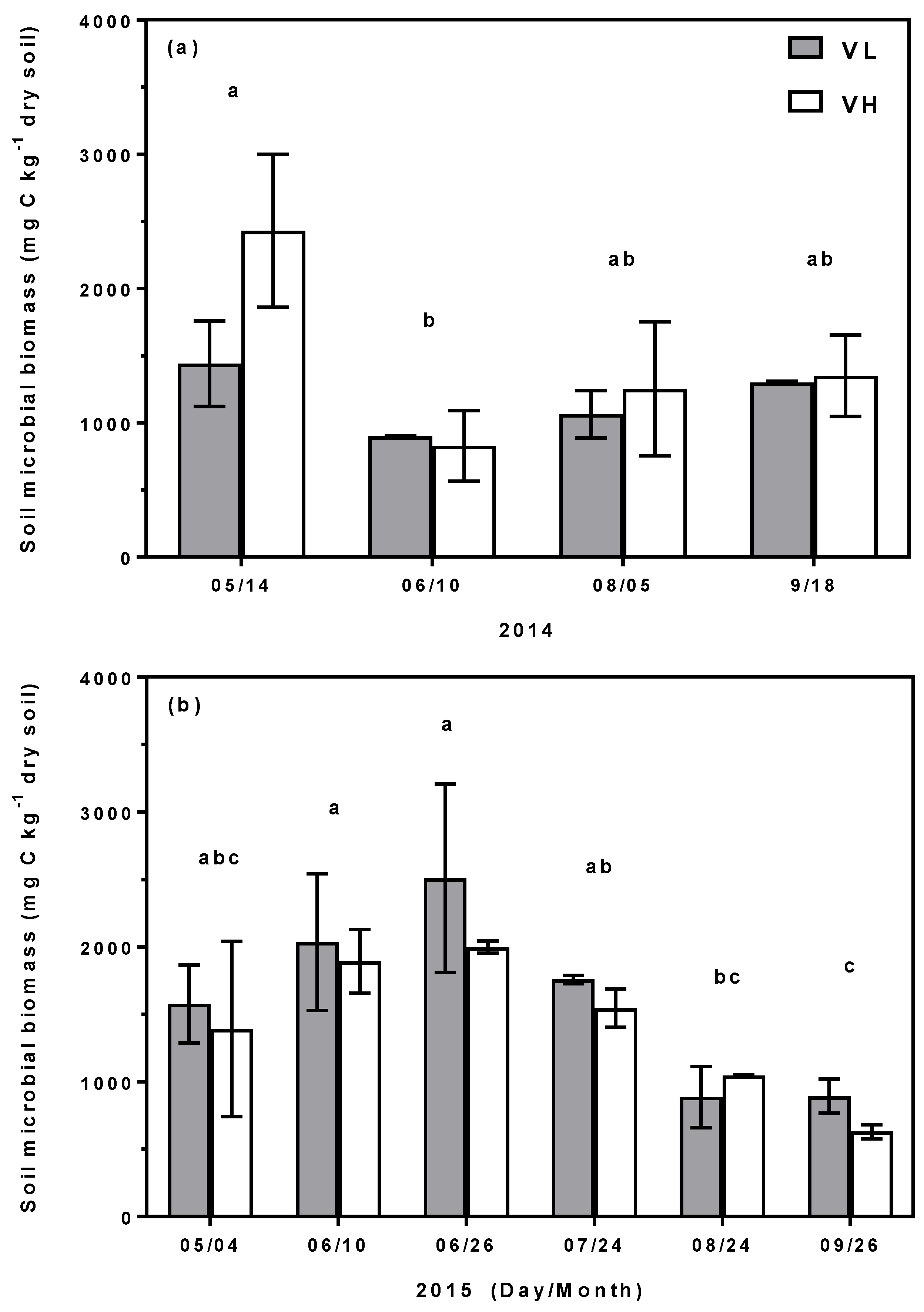
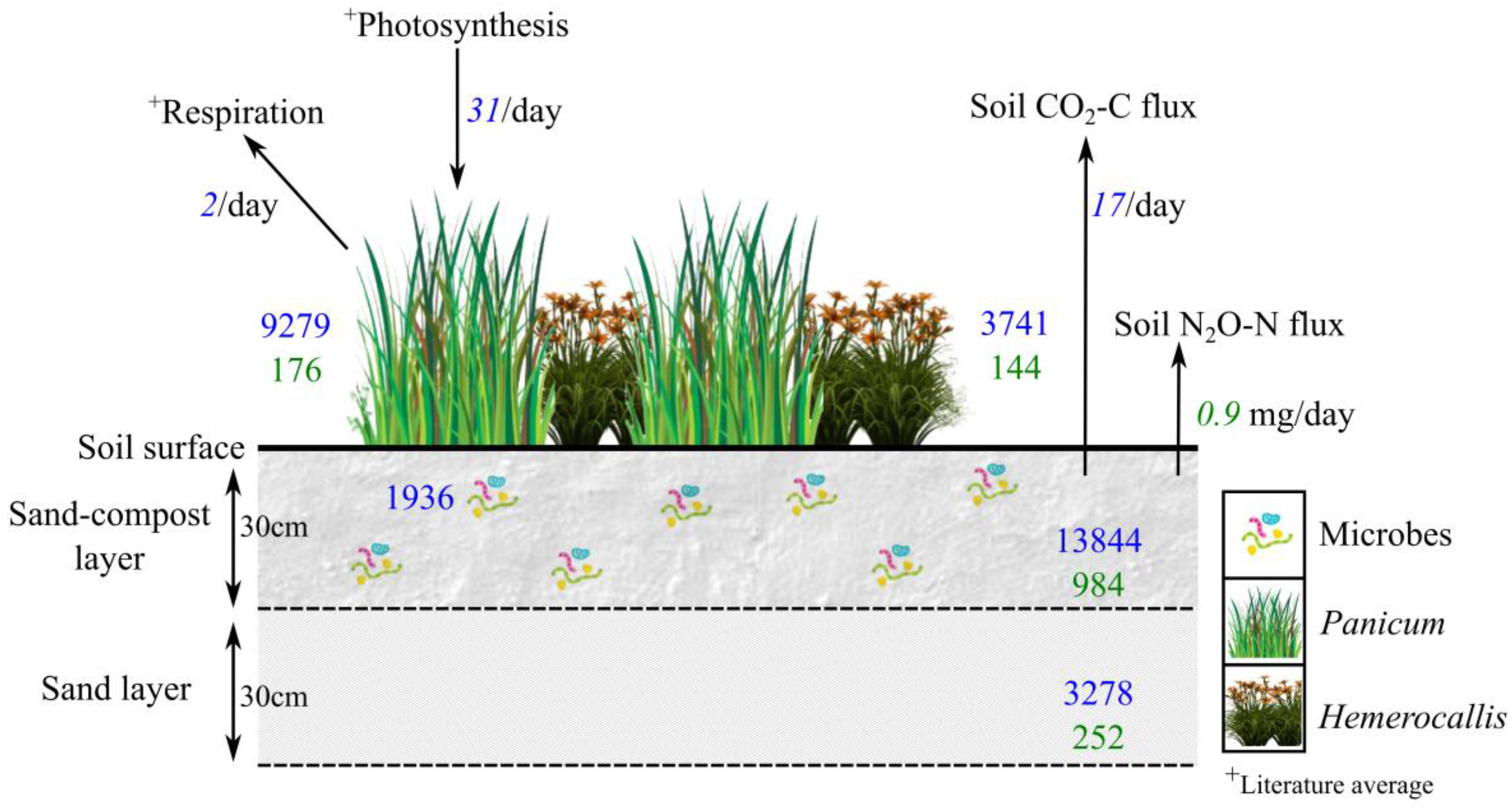
3.2. Bioretention C and N Pools
4. Discussion
4.1. Treatment Effects on Gas Fluxes
4.1.1. CO2 Fluxes
4.1.2. N2O Fluxes
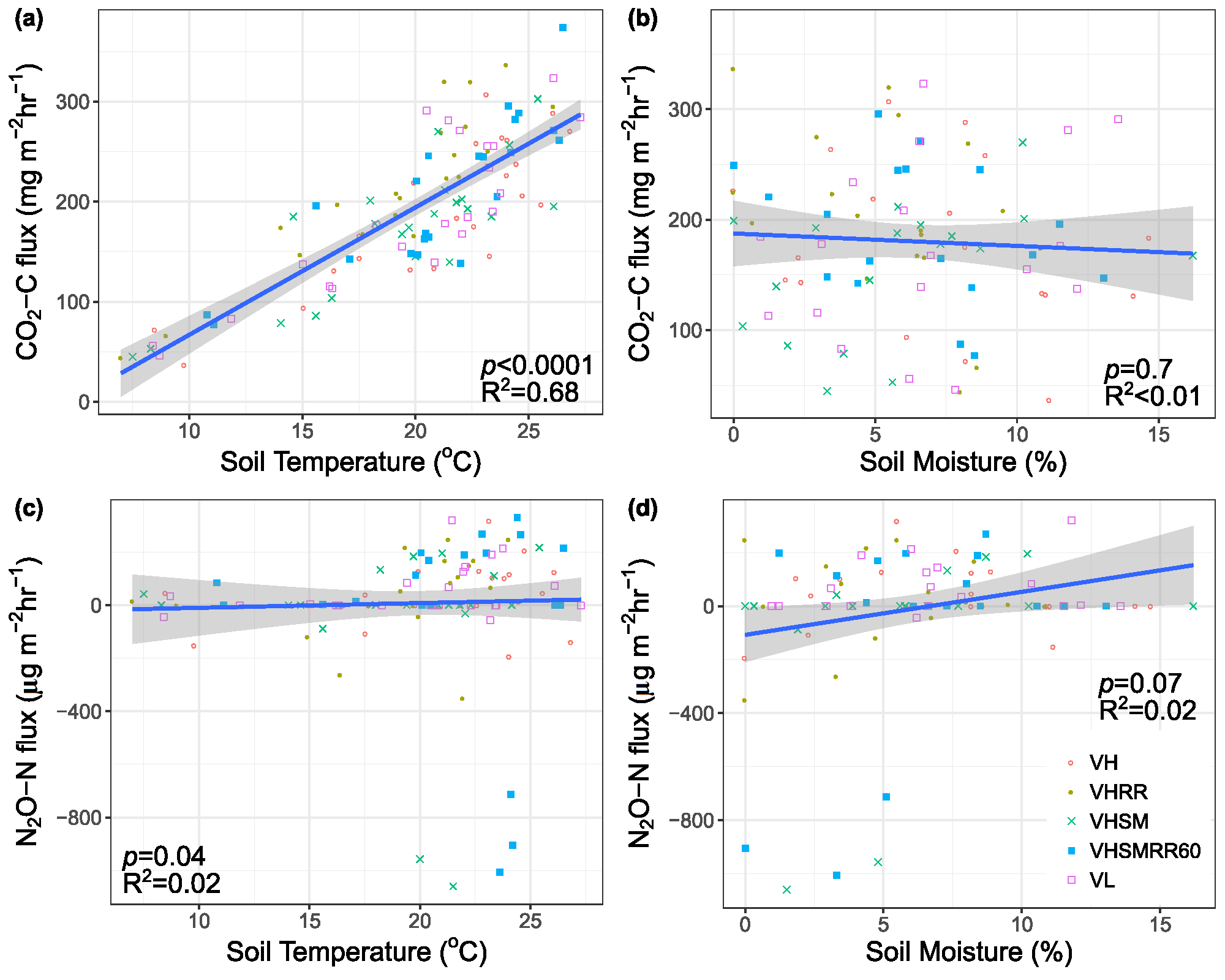
4.2. Environmental Effects on Fluxes
4.3. Carbon and Nitrogen Partitioned Stocks in Soils, Microbial Biomass, and Plants
4.4. Comparison of Bioretention Fluxes to Other Landscapes
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- United States Environmental Protection Agency. What is Green Infrastructure? Available online: http://water.epa.gov/infrastructure/greeninfrastructure/gi_what.cfm (accessed on 8 February 2018).
- Nocco, M.A.; Rouse, S.E.; Balster, N.J. Vegetation type alters water and nitrogen budgets in a controlled, replicated experiment on residential-sized rain gardens planted with prairie, shrub, and turfgrass. Urban Ecosyst. 2016, 19, 1665–1691. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Special Report of Working Groups 1 and 2: Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Zahran, S.; Brody, S.D.; Vedlitz, A.; Grover, H.; Miller, C. Vulnerability and capacity: Explaining local commitment to climate-change policy. Environ. Plan. C Gov. Policy 2008, 26, 544–562. [Google Scholar] [CrossRef]
- Galford, G.L.; Ann, H.; Sam, C.; Sarah, F.; Julie, N.; Elizabeth, P.; Sarah, P.; Kristin, U.; Daniel, V.B. Considering Vermont’s Future in a Changing Climate: The First Vermont Climate Assessment; Gund Institute for Ecological Economics: Burlington, VT, USA, 2014; p. 219. [Google Scholar]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Miltner, E.; Cogger, C. Carbon Sequestration Potential in Urban Soils. In Carbon Sequestration in Urban Ecosystems; Springer: Dordrecht, The Nederland, 2012; pp. 173–196. [Google Scholar] [CrossRef]
- Northeast Climate Impacts Assessment (NECIA), Union of Concerned Scientists. Climate Change in the U.S. Northeast. A Report of the Northeast Climate Impacts Assessment; Union of Concerned Scientists (UCS): Washington, DC, USA, 2006. [Google Scholar]
- Liu, J.; Sample, D.J.; Bell, C.; Guan, Y. Review and research needs of bioretention used for the treatment of urban stormwater. Water 2014, 6, 1069–1099. [Google Scholar] [CrossRef]
- Bratieres, K.; Fletcher, T.D.; Deletic, A.; Zinger, Y. Nutrient and sediment removal by stormwater biofilters: A large-scale design optimisation study. Water Res. 2008, 42, 3930–3940. [Google Scholar] [CrossRef] [PubMed]
- Hatt, B.E.; Fletcher, T.D.; Deletic, A. Hydrologic and pollutant removal performance of stormwater biofiltration systems at the field scale. J. Hydrol. 2009, 365, 310–321. [Google Scholar] [CrossRef]
- Grover, S.P.; Cohan, A.; Chan, H.S.; Livesley, S.J.; Beringer, J.; Daly, E. Occasional large emissions of nitrous oxide and methane observed in stormwater biofiltration systems. Sci. Total Environ. 2013, 465, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Ewel, K.C.; Cropper, W.P., Jr.; Gholz, H.L. Soil CO2 evolution in Florida slash pine plantations. II. Importance of root respiration. Can. J. For. Res. 1987, 17, 330–333. [Google Scholar] [CrossRef]
- Lytle, D.E.; Cronan, C.S. Comparative soil CO2 evolution, litter decay, and root dynamics in clearcut and uncut spruce-fir forest. For. Ecol. Manag. 1998, 103, 121–128. [Google Scholar] [CrossRef]
- Verstraete, W.; Focht, D.D. Biochemical Ecology of Nitrification and Denitrification. In Advances in Microbial Ecology, Advances in Microbial Ecology; Springer: Boston, MA, USA, 1977; pp. 135–214. [Google Scholar] [CrossRef]
- Groffman, P.M.; Boulware, N.J.; Zipperer, W.C.; Pouyat, R.V.; Band, L.E.; Colosimo, M.F. Soil nitrogen cycle processes in urban riparian zones. Environ. Sci. Technol. 2002, 36, 4547–4552. [Google Scholar] [CrossRef] [PubMed]
- McPhillips, L.; Goodale, C.; Walter, M.T. Nutrient Leaching and Greenhouse Gas Emissions in Grassed Detention and Bioretention Stormwater Basins. J. Sustain. Water Built Environ. 2018, 4, 04017014. [Google Scholar] [CrossRef]
- Pang, X.; Bao, W.; Zhu, B.; Cheng, W. Responses of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agric. For. Meteorol. 2013, 171, 57–64. [Google Scholar] [CrossRef]
- Maier, C.A.; Kress, L.W. Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability. Can. J. For. Res. 2000, 30, 347–359. [Google Scholar] [CrossRef]
- Shrestha, P.; Hurley, S.; Wemple, B.C. Effects of different soil media, vegetation, and hydrologic treatments on nutrient and sediment removal in roadside bioretention systems. Ecol. Eng. 2018, 112, 116–131. [Google Scholar] [CrossRef]
- Cording, A.; Hurley, S.; Whitney, D. Monitoring methods and designs for evaluating. J. Environ. Eng. 2017, 143, 05017006. [Google Scholar] [CrossRef]
- U.S. Census Bureau. Census Interactive Population Search. Available online: https://www.census.gov/2010census/popmap/ipmtext.php (accessed on 8 February 2018).
- National Climatic Data Center, NOAA, Burlington Vermont Airport (Station GHCND: USW00014742). Available online: http://www.ncdc.noaa.gov/oa/ncdc.html (accessed on 28 December 2017).
- Hutchinson, G.L.; Livingston, G.P. Vents and seals in non-steady-state chambers used for measuring gas exchange between soil and the atmosphere. Eur. J. Soil Sci. 2001, 52, 675–682. [Google Scholar] [CrossRef]
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. Fine root dynamics, coarse root biomass, root distribution, and soil respiration in a multispecies riparian buffer in Central Iowa, USA. Agrofor. Syst. 1998, 44, 163–174. [Google Scholar] [CrossRef]
- McPhillips, L.; Walter, M.T. Hydrologic conditions drive denitrification and greenhouse gas emissions in stormwater detention basins. Ecol. Eng. 2015, 85, 67–75. [Google Scholar] [CrossRef]
- Iqbal, J.; Castellano, M.J.; Parkin, T.B. Evaluation of photoacoustic infrared spectroscopy for simultaneous measurement of N2O and CO2 gas concentrations and fluxes at the soil surface. Glob. Chang. Biol. 2013, 19, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, G.L.; Mosier, A.R. Improved soil cover method for field measurement of nitrous oxide fluxes. Soil Sci. Soc. Am. J. 1981, 45, 311–316. [Google Scholar] [CrossRef]
- Rochette, P.; Bertrand, N. Soil-surface gas emissions. In Soil Sampling and Methods of Analysis; Carter, M.R., Gregorich, E.G., Eds.; Canadian Society of Soil Science; CRC Press: Boca Raton, FL, USA, 2008; pp. 851–861. [Google Scholar]
- Jenkinson, D.S.; Powlson, D.S. The effects of biocidal treatments on metabolism in soil—V: A method for measuring soil biomass. Soil Biol. Biochem. 1976, 8, 209–213. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Beck, T.; Joergensen, R.G.; Kandeler, E.; Makeschin, F.; Nuss, E.; Oberholzer, H.R.; Scheu, S. An inter-laboratory comparison of ten different ways of measuring soil microbial biomass C. Soil Biol. Biochem. 1997, 29, 1023–1032. [Google Scholar] [CrossRef]
- Heaton, E.; Voigt, T.; Long, S.P. A quantitative review comparing the yields of two candidate C 4 perennial biomass crops in relation to nitrogen, temperature and water. Biomass Bioenergy 2004, 27, 21–30. [Google Scholar] [CrossRef]
- Hogberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekbald, A.; Hogberg, M.N.; Nyberg, G.; Lofvenius, M.O.; Read, D.J. Large scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–791. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.A. Soil Microbiology, Ecology and Biochemistry; Academic Press: Atlanta, GA, USA, 2014. [Google Scholar]
- Avrahami, S.; Conrad, R.; Braker, G. Effect of Soil Ammonium Concentration on N2O Release and on the Community Structure of Ammonia Oxidizers and Denitrifiers. Appl. Environ. Microbiol. 2002, 68, 5685–5692. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, M.A.; Dick, W.A. Distribution and stability of pyrophosphatase in soils. Soil Biol. Biochem. 1979, 11, 655–659. [Google Scholar] [CrossRef]
- Goberna, M.; Sánchez, J.; Pascual, J.A.; García, C. Surface and subsurface organic carbon, microbial biomass and activity in a forest soil sequence. Soil Biol. Biochem. 2006, 38, 2233–2243. [Google Scholar] [CrossRef]
- Calderón, F.J.; Jackson, L.E.; Scow, K.M.; Rolston, D.E. Short-Term Dynamics of Nitrogen, Microbial Activity, and Phospholipid Fatty Acids after Tillage. Soil Sci. Soc. Am. J. 2001, 65, 118–126. [Google Scholar] [CrossRef]
- Dietz, M.E.; Clausen, J.C. Saturation to improve pollutant retention in a rain garden. Environ. Sci. Technol. 2006, 40, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Lucas, W.; Greenway, M. A comparative study of nutrient retention performance in vegetated and non-vegetated bioretention mesocosms. NOVATECH 2007, section 5.2, 1086–1089. [Google Scholar]
- Pataki, D.E.; Carreiro, M.M.; Cherrier, J.; Grulke, N.E.; Jennings, V.; Pincetl, S.; Pouyat, R.V.; Whitlow, T.H.; Zipperer, W.C. Coupling biogeochemical cycles in urban environments: Ecosystem services, green solutions, and misconceptions. Front. Ecol. Environ. 2011, 9, 27–36. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Lichter, J. Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2. Nature 2001, 411, 466. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Ngouajio, M. Soil microbial biomass, functional microbial diversity, and nematode community structure as affected by cover crops and compost in an organic vegetable production system. Appl. Soil Ecol. 2012, 58, 45–55. [Google Scholar] [CrossRef]
- Malhi, S.S.; McGill, W.B. Nitrification in three Alberta soils: Effect of temperature, moisture and substrate concentration. Soil Biol. Biochem. 1982, 14, 393–399. [Google Scholar] [CrossRef]
- Stevens, R.J.; Laughlin, R.J.; Burns, L.C.; Arah, J.R.M.; Hood, R.C. Measuring the contributions of nitrification and denitrification to the flux of nitrous oxide from soil. Soil Biol. Biochem. 1997, 29, 139–151. [Google Scholar] [CrossRef]
- Yoshinari, T.; Hynes, R.; Knowles, R. Acetylene inhibition of nitrous oxide reduction and measurement of denitrification and nitrogen fixation in soil. Soil Biol. Biochem. 1977, 9, 177–183. [Google Scholar] [CrossRef]
- Sørensen, J. Denitrification Rates in a Marine Sediment as Measured by the Acetylene Inhibition Technique. Appl. Environ. Microbiol. 1978, 36, 139–143. [Google Scholar] [PubMed]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Raich, J.W.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Liikanen, A.; Huttunen, J.T.; Karjalainen, S.M.; Heikkinen, K.; Väisänen, T.S.; Nykänen, H.; Martikainen, P.J. Temporal and seasonal changes in greenhouse gas emissions from a constructed wetland purifying peat mining runoff waters. Ecol. Eng. 2006, 26, 241–251. [Google Scholar] [CrossRef]
- Søvik, A.K.; Augustin, J.; Heikkinen, K.; Huttunen, J.T.; Necki, J.M.; Karjalainen, S.M.; Kløve, B.; Liikanen, A.; Mander, Ü.; Puustinen, M.; et al. Emission of the greenhouse gases nitrous oxide and methane from constructed wetlands in Europe. J. Environ. Qual. 2006, 35, 2360–2373. [Google Scholar] [CrossRef] [PubMed]
- Bettez, N.D.; Groffman, P.M. Denitrification Potential in Stormwater Control Structures and Natural Riparian Zones in an Urban Landscape. Environ. Sci. Technol. 2012, 46, 10909–10917. [Google Scholar] [CrossRef] [PubMed]
- Decina, S.M.; Hutyra, L.R.; Gately, C.K.; Getson, J.M.; Reinmann, A.B.; Gianotti, A.G.S.; Templer, P.H. Soil respiration contributes substantially to urban carbon fluxes in the greater Boston area. Environ. Pollut. 2016, 212, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Thomson, P.E.; Clayton, H.; Mctaggart, I.P.; Conen, F. Effects of temperature, water content and nitrogen fertilisation on emissions of nitrous oxide by soils. Atmos. Environ. 1998, 32, 3301–3309. [Google Scholar] [CrossRef]
- Tzoulas, K.; Korpela, K.; Venn, S.; Yli-Pelkonen, V.; Kaźmierczak, A.; Niemela, J.; James, P. Promoting ecosystem and human health in urban areas using Green Infrastructure: A literature review. Landsc. Urban Plan. 2007, 81, 167–178. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Photosynthesis, respiration, and long-distance transport. In Plant Physiological Ecology; Springer: Dordrecht, The Nederland, 1998; pp. 10–153. [Google Scholar]
- Ryan, M.G. Effects of climate change on plant respiration. Ecol. Appl. 1991, 1, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, M.A.; Reed, R.L.; McLaughlin, S.B.; Wullschleger, S.D.; Conger, B.V.; Parrish, D.J.; Wolf, D.D.; Taliaferro, C.; Hopkins, A.A.; Ocumpaugh, W.R.; et al. Switchgrass as a sustainable bioenergy crop. Bioresour. Technol. 1996, 56, 83–93. [Google Scholar] [CrossRef]
- Davidson, E.A.; Savage, K.; Verchot, L.V.; Navarro, R. Minimizing artifacts and biases in chamber-based measurements of soil respiration. Agric. For. Meteorol. 2002, 113, 21–37. [Google Scholar] [CrossRef]
- Kaye, J.P.; Groffman, P.M.; Grimm, N.B.; Baker, L.A.; Pouyat, R.V. A distinct urban biogeochemistry? Trends Ecol. Evol. 2006, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Zhang, J.; Li, P.; Xie, H.; Wu, J.; Wang, J. Nitrous oxide emissions from surface flow and subsurface flow constructed wetland microcosms: Effect of feeding strategies. Ecol. Eng. 2011, 37, 1815–1821. [Google Scholar] [CrossRef]
- Dotro, G.; Jefferson, B.; Jones, M.; Vale, P.; Cartmell, E.; Stephenson, T. A review of the impact and potential of intermittent aeration on continuous flow nitrifying activated sludge. Environ. Technol. 2011, 32, 1685–1697. [Google Scholar] [CrossRef]
- Mander, Ü.; Dotro, G.; Ebie, Y.; Towprayoon, S.; Chiemchaisri, C.; Nogueira, S.F.; Jamsranjav, B.; Kasak, K.; Truu, J.; Tournebize, J.; et al. Greenhouse gas emission in constructed wetlands for wastewater treatment: A review. Ecol. Eng. 2014, 66, 19–35. [Google Scholar] [CrossRef]
- Livesley, S.J.; Dougherty, B.J.; Smith, A.J.; Navaud, D.; Wylie, L.J.; Arndt, S.K. Soil-atmosphere exchange of carbon dioxide, methane and nitrous oxide in urban garden systems: Impact of irrigation, fertiliser and mulch. Urban Ecosyst. 2010, 13, 273–293. [Google Scholar] [CrossRef]
- Townsend-Small, A.; Czimczik, C.I. Carbon sequestration and greenhouse gas emissions in urban turf. Geophys. Res. Lett. 2010, 37, L02707. [Google Scholar] [CrossRef]
- Kaye, J.P.; Burke, I.C.; Mosier, A.R.; Pablo Guerschman, J. Methane and nitrous oxide fluxes from urban soils to the atmosphere. Ecol. Appl. 2004, 14, 975–981. [Google Scholar] [CrossRef]
- Giasson, M.-A.; Ellison, A.M.; Bowden, R.D.; Crill, P.M.; Davidson, E.A.; Drake, J.E.; Frey, S.D.; Hadley, J.L.; Lavine, M.; Melillo, J.M.; et al. Soil respiration in a northeastern US temperate forest: A 22-year synthesis. Ecosphere 2013, 4, 1–28. [Google Scholar] [CrossRef]
- Ma, B.L.; Wu, T.Y.; Tremblay, N.; Deen, W.; Morrison, M.J.; Mclaughlin, N.B.; Gregorich, E.G.; Stewart, G. Nitrous oxide fluxes from corn fields: On-farm assessment of the amount and timing of nitrogen fertilizer. Glob. Chang. Biol. 2010, 16, 156–170. [Google Scholar] [CrossRef]
- Vymazal, J. Constructed wetlands for wastewater treatment. Water 2010, 2, 530–549. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil OM (%) | Soil C:N | Soil pH | Soil EC (µS cm−1) | Soil Media Bulk Density (g cm−3 Dry Soil) | Soil NH4+-N Concentration (ppm) | Soil NO3−-N Concentration (ppm) |
|---|---|---|---|---|---|---|
| 1.95 (0.09) | 13.39 (0.65) | 7.04 (0.02) | 30.18 (0.23) | 1.19 (0.03) | 0.311 (0.10) | 3.932 (0.69) |
| n = 7 | n = 2 | n = 7 | n = 7 | n = 2 | n = 3 | n = 3 |
| Sampled Year | Soil Depth | Total C | Total N | C:N |
|---|---|---|---|---|
| g kg−1 Dry Soil | ||||
| May 2014 | 0–10 cm | 18.36 | 1.69 | 10.9 |
| May 2016 | 0–10 cm | 14.17 | 0.9 | 15.7 |
| November 2016 | 0–30 cm | 10.27 | 0.73 | 14.1 |
| November 2016 | 30–40 cm | 1.82 | 0.14 | 13.5 |
| Season | Plant Species | C | N | C:N |
|---|---|---|---|---|
| g kg−1 Dry Plant | ||||
| Summer (August) | Panicum | 446 | 11.4 | 39 |
| Hemerocallis | 445 | 24.6 | 18 | |
| Fall (November) | Panicum | 461 | 5.85 | 79 |
| Hemerocallis | 449 | 9.76 | 46 | |
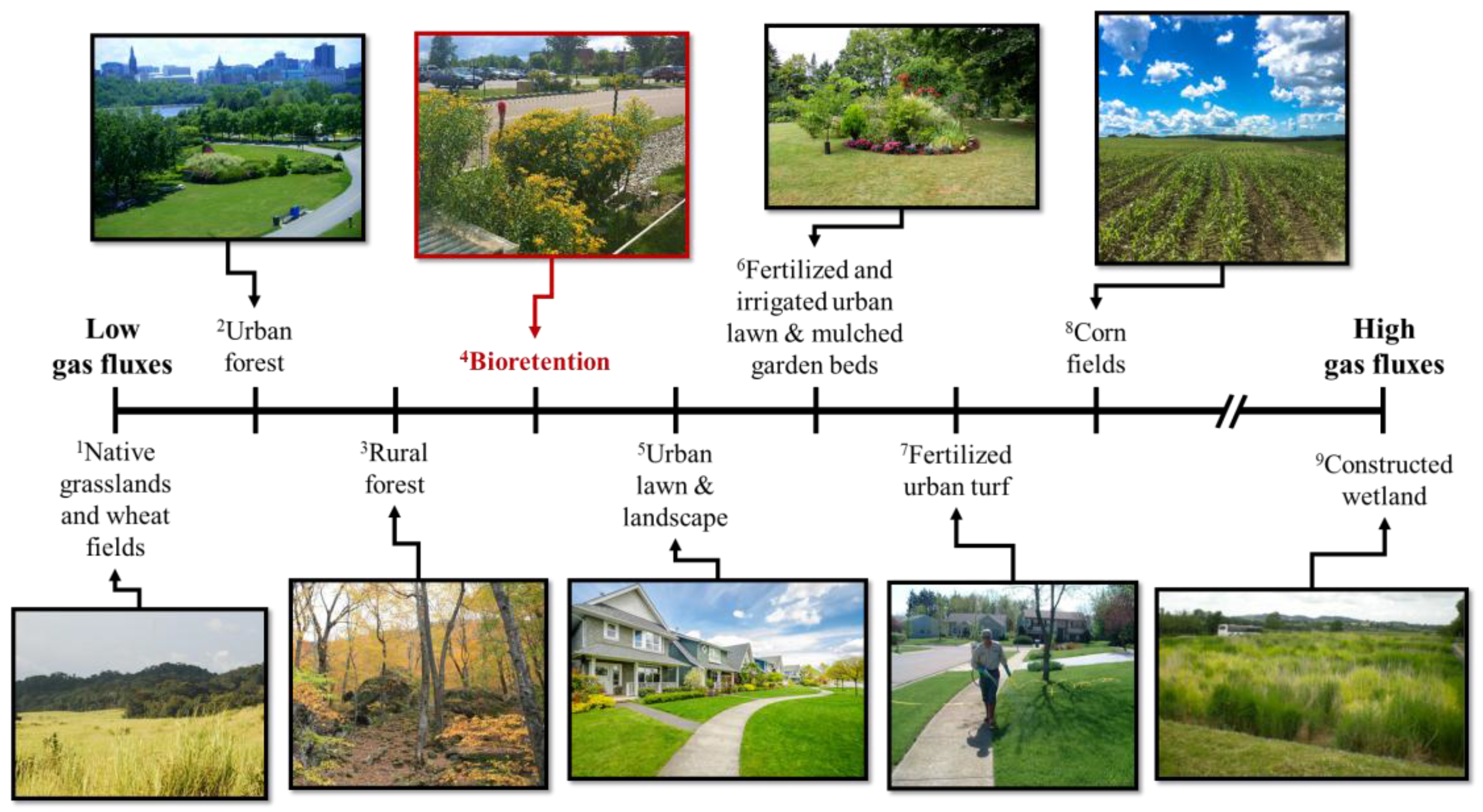
| System | Reference | Study Location | CO2-C Flux (mg C m−2 h−1) | N2O-N Flux (µg N m−2 h−1) | Image Credit |
|---|---|---|---|---|---|
| 1Native grasslands and wheat fields (unfertilized & unirrigated) | [69] | Colorado, USA | na | <4 | www.worldwildlife.org |
| 2Urban Forest | [56] | Massachusetts, USA | 113 | na | Paliza Shrestha |
| 3Rural forest | [70] | Massachusetts, USA | 133 | na | Paliza Shrestha |
| 4Bioretention | This Study | Vermont, USA | 194 | 10 | Paliza Shrestha |
| 5Urban lawn & landscaped | [56] | Massachusetts, USA | 194 & 291 | na | www.zillow.com |
| 6Fertilized and irrigated urban lawn | [69] | Colorado, USA | na | >10 | Elizabeth Schaller Bhuju |
| 6Fertilized and irrigated urban lawn & mulched garden beds | [67] | Melbourne, Australia | ~450 | 17 to 28 & 14 | Elizabeth Schaller Bhuju |
| 7Fertilized urban turf | [68] | Southern California, USA | na | 93 | mda.maryland.gov |
| 8Fertilized corn fields | [71] | Ontario, Canada | na | 30 to 900 | Vanesa Perillo |
| 9Constructed Wetland | [54] | Northern Europe (Estonia, Finland, Norway, Poland) | −35 to 3875 | −88 to 41,600 | [72] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrestha, P.; Hurley, S.E.; Adair, E.C. Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems. Water 2018, 10, 185. https://doi.org/10.3390/w10020185
Shrestha P, Hurley SE, Adair EC. Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems. Water. 2018; 10(2):185. https://doi.org/10.3390/w10020185
Chicago/Turabian StyleShrestha, Paliza, Stephanie E. Hurley, and E. Carol Adair. 2018. "Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems" Water 10, no. 2: 185. https://doi.org/10.3390/w10020185
APA StyleShrestha, P., Hurley, S. E., & Adair, E. C. (2018). Soil Media CO2 and N2O Fluxes Dynamics from Sand-Based Roadside Bioretention Systems. Water, 10(2), 185. https://doi.org/10.3390/w10020185