Assessing Eutrophication Potential of a Freshwater Lake by Relating Its Bioproductivity and Biodiversity: A Case Study of Lake Wilson on Central Oahu, Hawaii

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Investigation
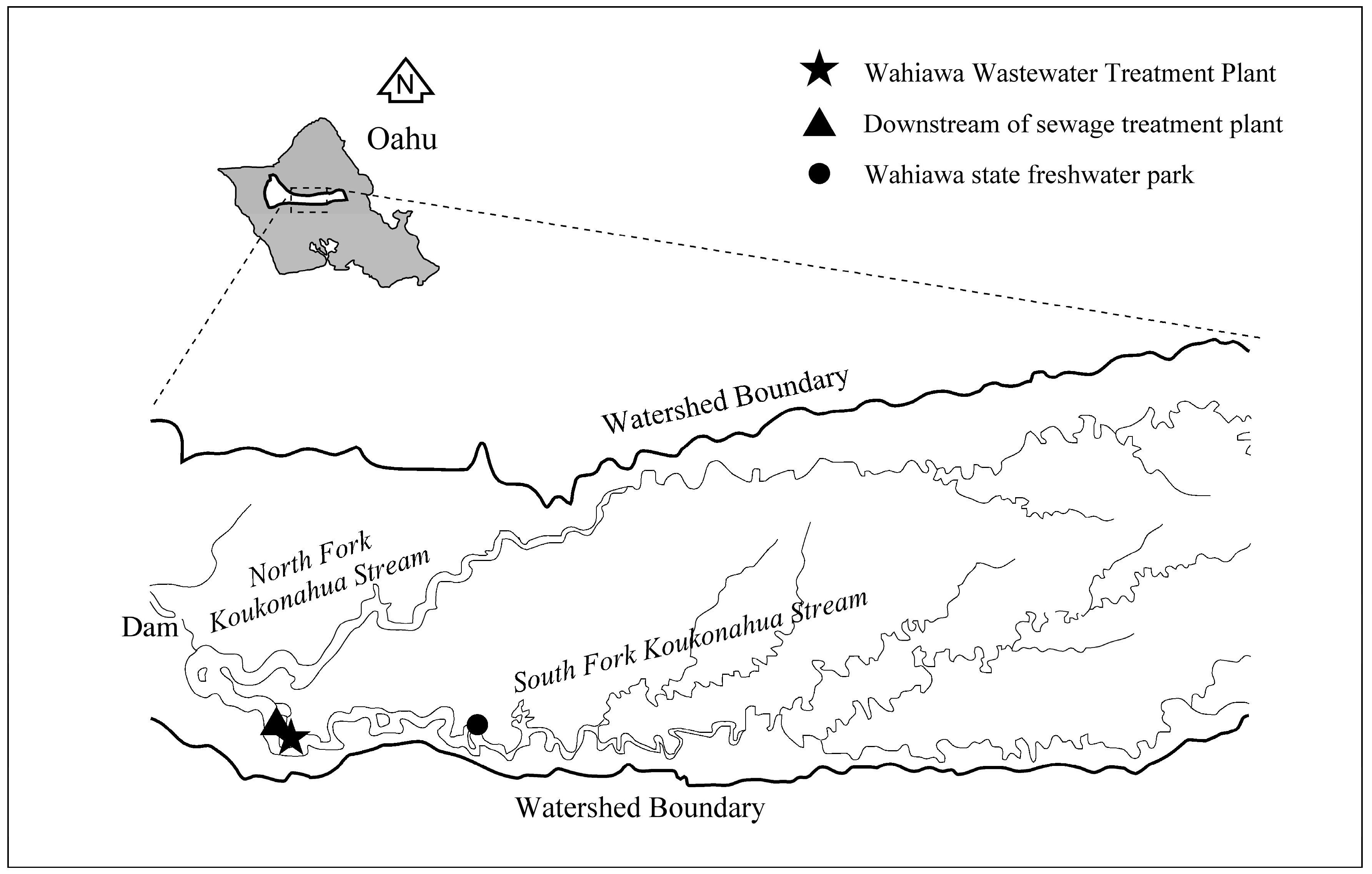
2.1.1. Study Site
2.1.2. Field Experiment
2.2. Laboratory Investigation
2.2.1. Laboratory Apparatus
2.2.2. Laboratory Experiments
2.2.3. Bioproductivity Determination
- Calculate the rate of change of Chl a concentration or dX/dt, X is the Chl a concentration.
- Calculate the washout terms of three microcosms or DX. Dilution rate D is defined as , the retention time of the three microcosms were regulated to be 4.8, 8.6, and 10.4 days for oligotrophic, mesotrophic, and eutrophic microcosms, respectively.
- Based on steps (1) and (2), calculate by Equations (4) and (6).
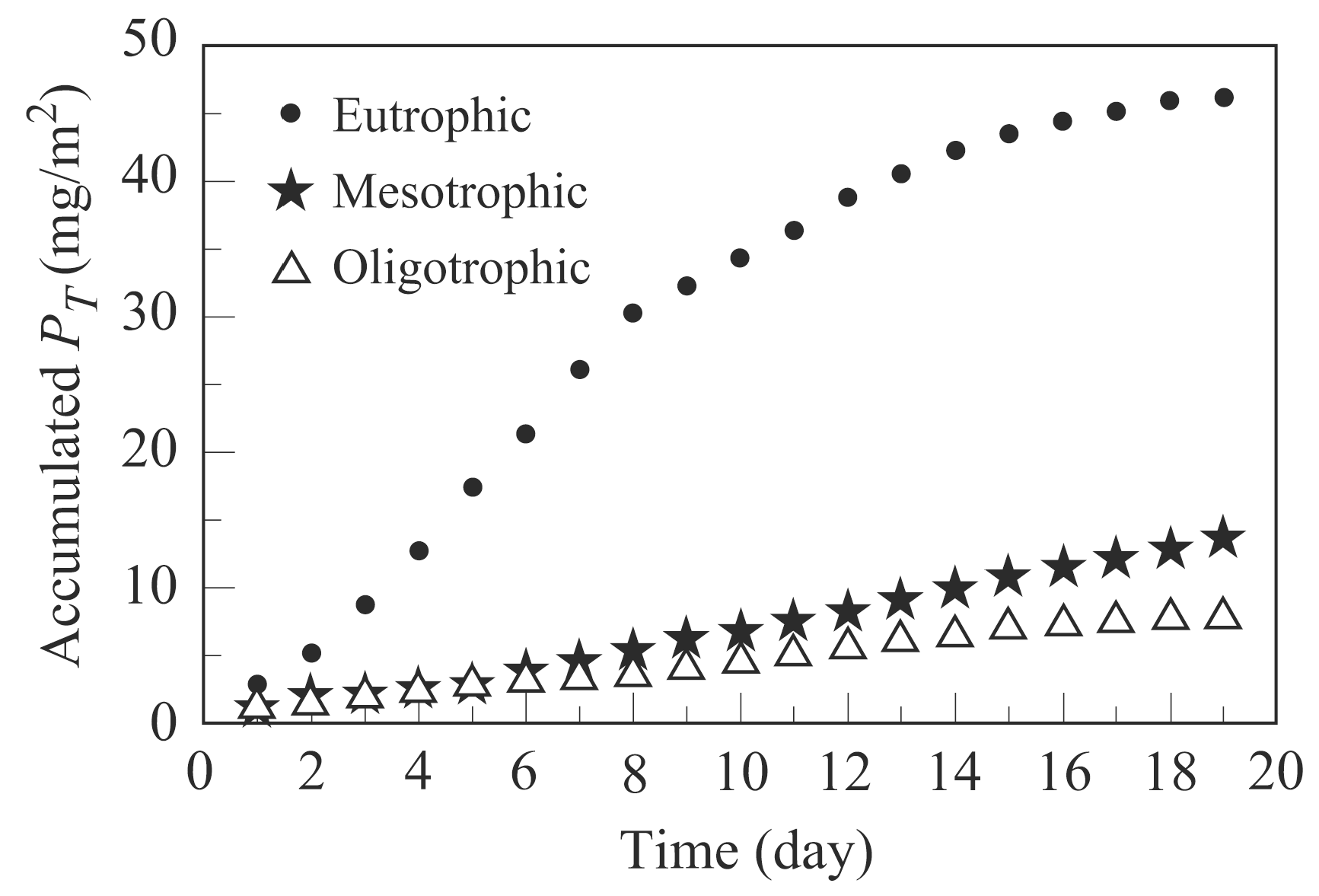
- Calcualte Cumulated areal algal product (PT, mg/m2) by Equation (8). Calculated results are shown in Section 3.2.2.
- Finally, average algal bioproductivities (, mg/m2/day) in the three microcosms was determined as PT/T.
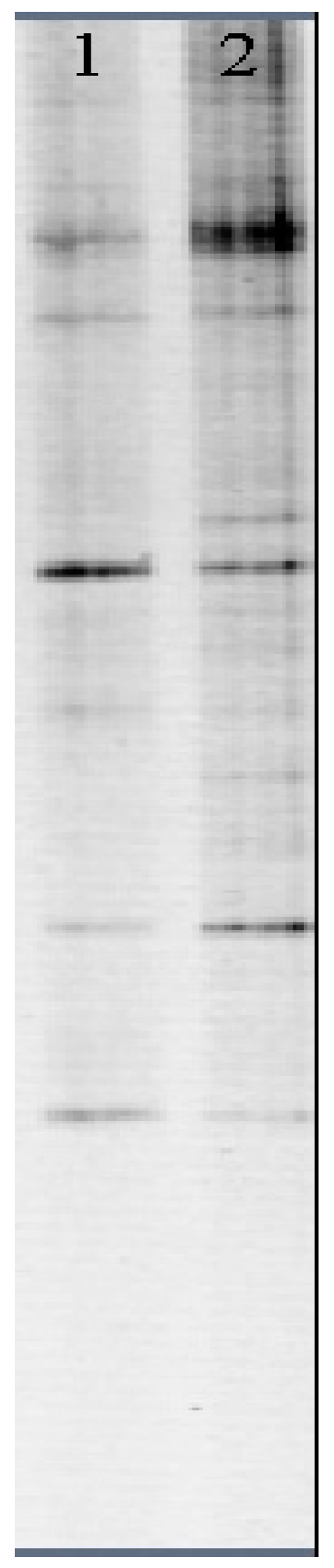
2.2.4. Genetic Biodiversity Determination
3. Results and Discussion
3.1. Trophic States and Genetic Biodiversity of Field Samples
3.2. Laboratory Investigation of Trophic States and Genetic Biodiversity
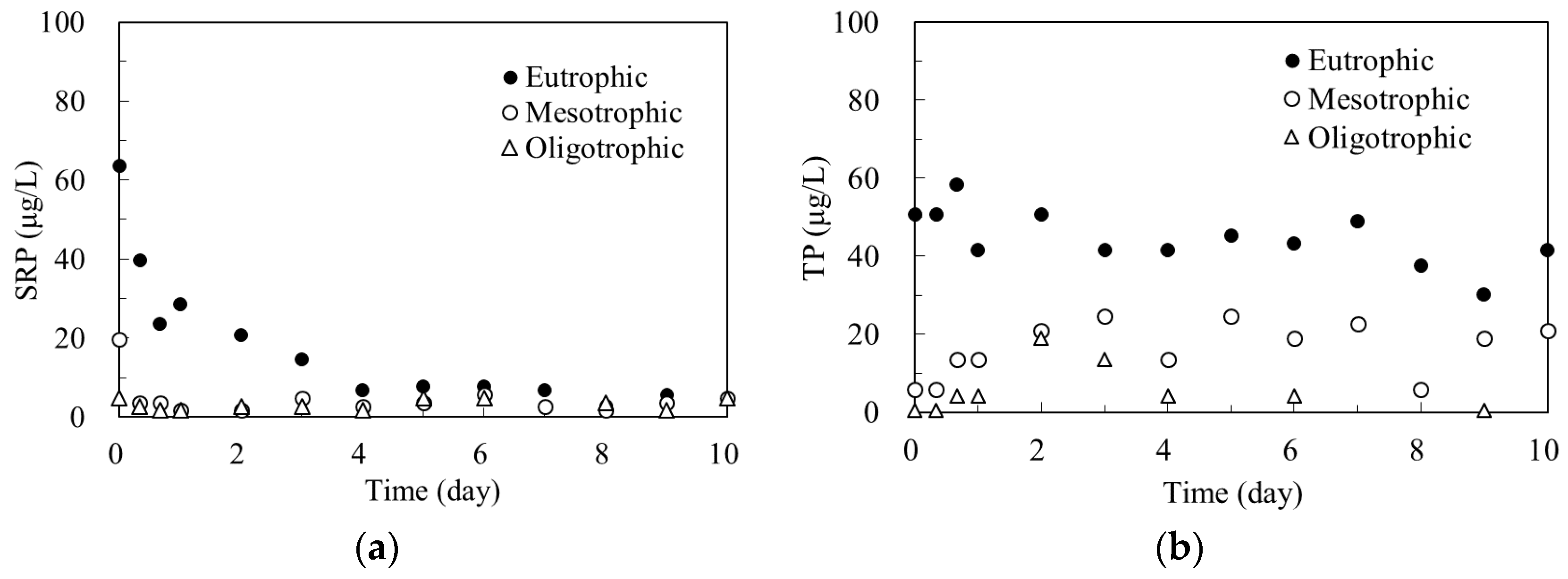
3.2.1. SRP and TP in Algal Microcosms with Different Trophic States
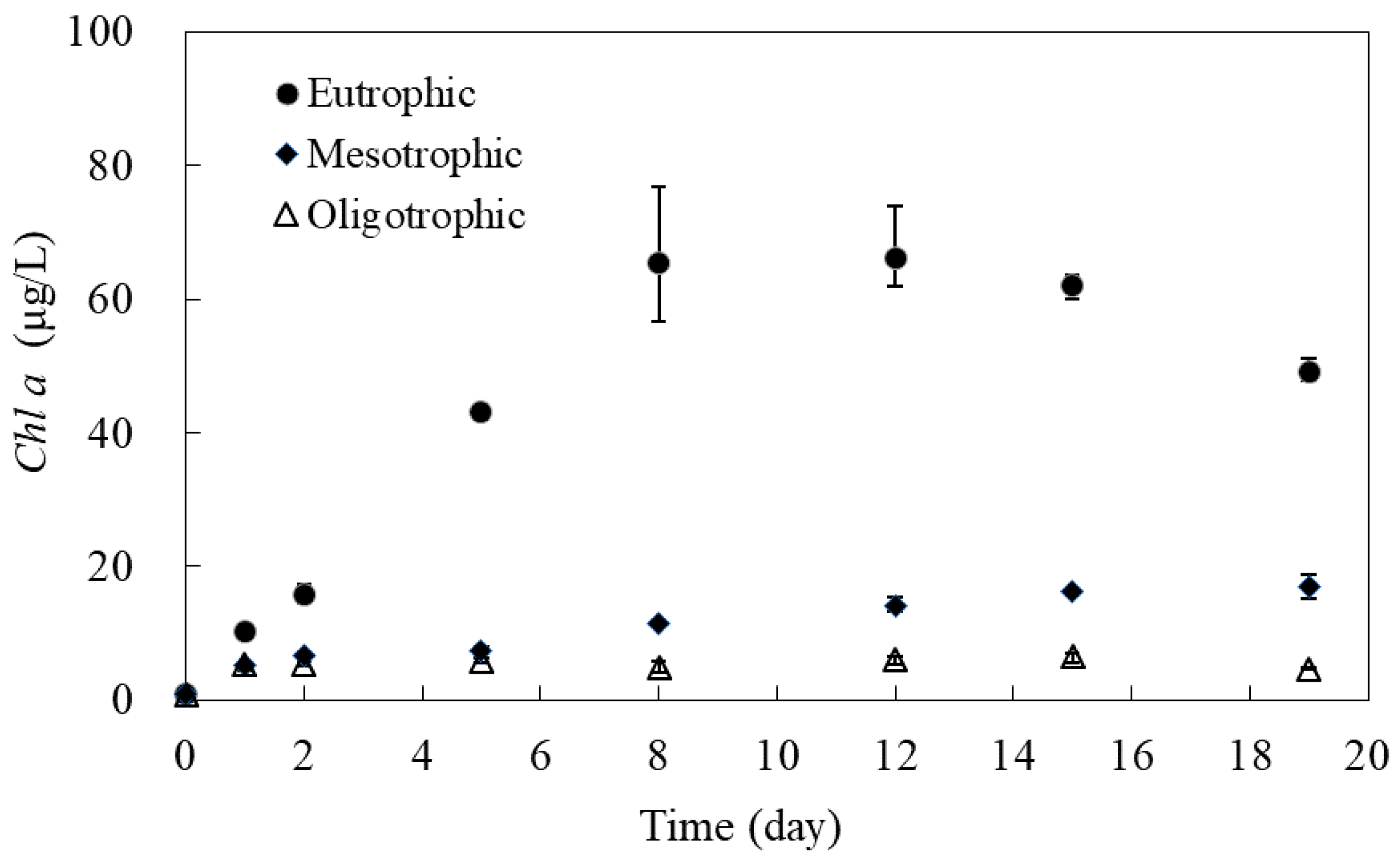
3.2.2. Algal Bioproductivity in Microcosms under Varying Trophic States
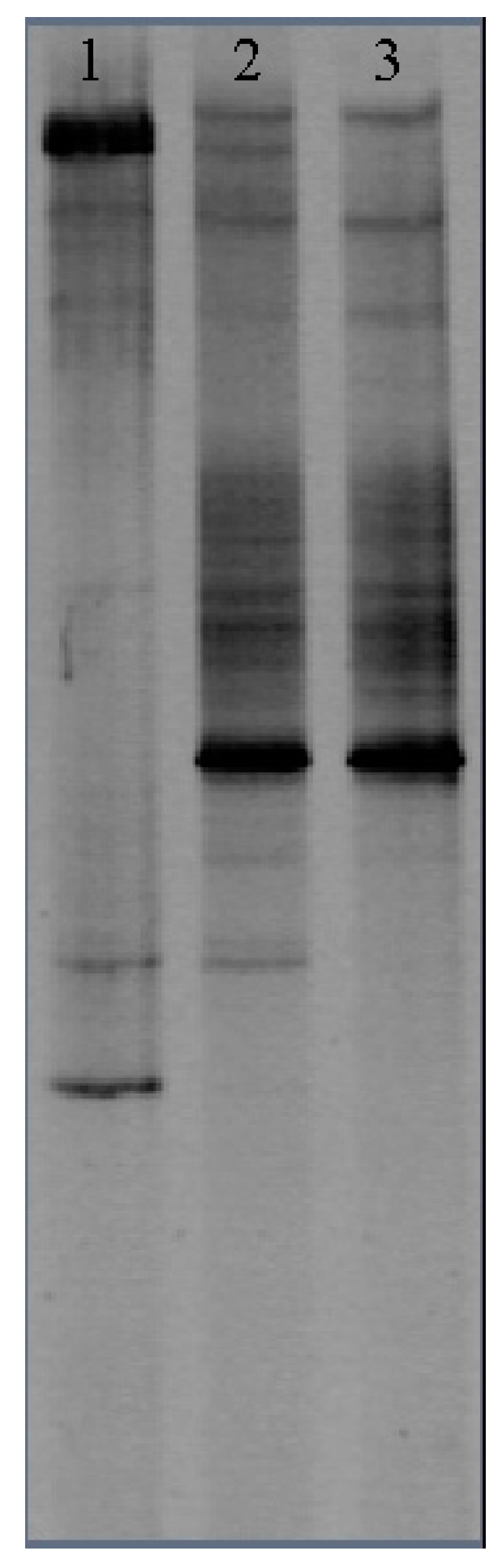
3.2.3. Genetic Biodiversity in Algal Microcosms with Different Trophic States
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E.M. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Long, T.; Liu, C.C.K. Algal bioproductivity in turbulent water: An experimental study. Water 2017, 9, 304. [Google Scholar]
- Chapra, S.C. The eutrophication problem and nutrients. In Surface Water-Quality Modeling; McGraw-Hill: Boston, MA, USA, 2008; pp. 521–558. [Google Scholar]
- Kuo, J.; Wang, Y.; Lung, W.; Liu, C.C.K. Real-time Eutrophication Control of a Reservoir. In Proceedings of the 2013 IAHR Congress, Chengdu, China, 8–13 September 2013; Tsinghua University Press: Beijing, China, 2013. [Google Scholar]
- Magurran, A.E. Biological diversity. Curr. Biol. 2005, 15, R116–R118. [Google Scholar] [CrossRef] [PubMed]
- Mark, N. The Encyclopedia of the Arctic; Routledge: New York, NY, USA, 2005; p. 264. [Google Scholar]
- Vellend, M. Species diversity and genetic diversity: Parallel processes and correlated patterns. Am. Nat. 2005, 166, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Caron, D.A.; Countway, P.D.; Savai, P.; Gast, R.J.; Schnetzer, A.; Moorthi, S.D.; Dennett, M.R.; Moran, D.M.; Jones, A.C. Defining DNA-based operational taxonomic units for microbial-eukaryote ecology. Appl. Environ. Microbiol. 2009, 75, 5797–5808. [Google Scholar] [CrossRef] [PubMed]
- Triadó-Margarit, X.; Casamayor, E.O. Genetic diversity of planktonic eukaryotes in high mountain lakes (Central Pyrenees, Spain). Environ. Microbiol. 2012, 14, 2445–2456. [Google Scholar] [CrossRef] [PubMed]
- Asai, R.; Ootani, K.; Nomura, Y.; Nakamura, C.; Ikebukuro, K.; Arikawa, Y.; Miyake, J.; Karube, I. PCR-based ribosomal DNA detection technique for microalga (Heterosigma carterae) causing red tide and its application to a biosensor using labeled probe. Mar. Biotechnol. 2003, 5, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Van Hannen, E.J.; Van Agterveld, M.P.; Gons, H.J.; Laanbroek, H.J. Revealing genetic diversity of eukaryotic microorganisms in aquatic environments by denaturing gradient gel electrophoresis. J. Phycol. 1998, 34, 206–213. [Google Scholar] [CrossRef]
- Muyzer, G.; Kornelia, S. Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE) in microbial ecology. Antonie Leeuwenhoek 1998, 73, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.; Thenot, A.; Lepere, C.; Debroas, D. Genetic diversity of small eukaryotes in lakes differing by their trophic status. Appl. Environ. Microbiol. 2005, 71, 5935–5942. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Chen, M.; Sun, Y.; Yang, J.; Chen, F. Genetic diversity of picoeukaryotes in eight lakes differing in trophic status. Can. J. Microbiol. 2011, 57, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H.; Orias, E. The ecological regulation of species diversity. Am. Nat. 1964, 98, 399–414. [Google Scholar] [CrossRef]
- Fukami, T.; Morin, P.J. Productivity-biodiversity relationships depend on the history of community assembly. Nature 2003, 24, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The Relationship between productivity and species richness. Annu. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Smith, V.H. Microbial diversity-productivity relationships in aquatic ecosystems. FEMS Microbiol. Ecol. 2007, 62, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Kassen, R.; Buckling, A.; Bell, G.; Rainey, P.B. Diversity peaks at intermediate productivity in a laboratory microcosm. Nature 2000, 406, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Young, R.H.F.; Dugan, G.L.; Lau, L.S.; Yamauchi, H. Eutrophication and Fish Toxicity Potentials in a Multiple-use Subtropical Reservoir; Technical Report No. 89; Water Resources Research Center, University of Hawaii at Manoa: Honolulu, HI, USA, 1975. [Google Scholar]
- Tetra Tech. Total Maximum Daily Loads (TMDLs) for the North and South Forks of Kaukonahua Stream, Oahu, Hawaii; State of Hawaii Department of Health: Honolulu, HI, USA, 2009. [Google Scholar]
- Vollenweider, R.A. Scientific Fundamentals of the Eutrophication of Lakes and Flowing Waters with Particular Reference to Nitrogen and Phosphorus as Factors in Eutrophication; Organization for Economic Co-operation and Development (OECD): Paris, France, 1971. [Google Scholar]
- Bold, H.C. The morphology of Chlamydomonas chlamydogama sp. nov. Bull. Torrey Bot. Club 1949, 76, 101–108. [Google Scholar] [CrossRef]
- Phillips, N.; Celia, M.S.; Clifford, W.M. An effective DNA extraction protocol for brown algae. Phycol. Res. 2001, 49, 97–102. [Google Scholar] [CrossRef]
- Van Hannen, E.J.; Zwart, G.; Van Agterveld, M.P.; Gons, H.J.; Ebert, J.; Laanbroek, H.J. Changes in bacterial and eukaryotic community structure after mass lysis of filamentous cyanobacteria associated with viruses. Appl. Environ. Microbiol. 1999, 65, 795–801. [Google Scholar]
- Muyzer, G.; Ellen, C.D.W.; Andre, G.U. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 397–423, 623–656. [Google Scholar] [CrossRef]
- Eichner, C.A.; Erb, R.W.; Timmis, K.N.; Wagner-Döbler, I. Thermal gradient gel electrophoresis analysis of bioprotection from pollutant shocks in the activated sludge microbial community. Appl. Environ. Microbiol. 1999, 65, 102–109. [Google Scholar] [PubMed]
- Fromin, N.; Hamelin, J.; Tarnawski, S.; Roesti, D.; Jourdain-Miserez, K.; Forestier, N.; Teyssier-Cuvelle, S.; Gillet, F.; Aragno, M.; Rossi, P. Statistical analysis of denaturing gel electrophoresis (DGE) fingerprinting patterns. Environ. Microbiol. 2002, 4, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.E.; Hollibaugh, J.T.; Orrego, C. Phylogenetic compositions of bacterioplankton from two California estuaries compared by denaturing gradient gel electrophoresis of 16S rDNA fragments. Appl. Environ. Microbiol. 1996, 62, 2676–2680. [Google Scholar] [PubMed]
- Hudson, J.J.; Taylor, W.D.; Schindler, D.W. Phosphate concentrations in lakes. Nature 2000, 406, 54–56. [Google Scholar] [CrossRef] [PubMed]
- Chapra, S.C.; Tarapchak, S. A Chlorophyll model and its relationship to phosphorus loading plots for lakes. Water Resour. Res. 1976, 12, 1260–1264. [Google Scholar] [CrossRef]
- Hutcheson, K. A test for comparing diversities based on the Shannon formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef]
- Magurran, A.E. Diversity indices and species abundance models. In Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; pp. 35–36. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locations | Chl a (µg/L) | TN (mg/L) | TOC (mg/L) | TP (µg/L) | SRP (µg/L) | TSS (mg/L) |
|---|---|---|---|---|---|---|
| DTSP | 23.9 | 0.35 | 4.14 | 58.3 | 17.7 | 5.0 |
| WSFP | 17.4 | 0.24 | 3.35 | 32.1 | 11.7 | 4.3 |
| Sampling Station | Lane | H′ | S |
|---|---|---|---|
| DSTP | 1 | 1.725 | 9 |
| WSFP | 2 | 2.571 | 20 |
| Trophic Levels | Lane | H′ | S |
|---|---|---|---|
| Oligotrophic | 1 | 1.762 | 7 |
| Mesotrophic | 2 | 2.390 | 13 |
| Eutrophic | 3 | 2.113 | 10 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.-C.; Liu, C.C.K. Assessing Eutrophication Potential of a Freshwater Lake by Relating Its Bioproductivity and Biodiversity: A Case Study of Lake Wilson on Central Oahu, Hawaii. Water 2018, 10, 296. https://doi.org/10.3390/w10030296
Lee T-C, Liu CCK. Assessing Eutrophication Potential of a Freshwater Lake by Relating Its Bioproductivity and Biodiversity: A Case Study of Lake Wilson on Central Oahu, Hawaii. Water. 2018; 10(3):296. https://doi.org/10.3390/w10030296
Chicago/Turabian StyleLee, Tsu-Chuan, and Clark C. K. Liu. 2018. "Assessing Eutrophication Potential of a Freshwater Lake by Relating Its Bioproductivity and Biodiversity: A Case Study of Lake Wilson on Central Oahu, Hawaii" Water 10, no. 3: 296. https://doi.org/10.3390/w10030296
APA StyleLee, T. -C., & Liu, C. C. K. (2018). Assessing Eutrophication Potential of a Freshwater Lake by Relating Its Bioproductivity and Biodiversity: A Case Study of Lake Wilson on Central Oahu, Hawaii. Water, 10(3), 296. https://doi.org/10.3390/w10030296