Bioturbation Effects of Chironomid Larvae on Nitrogen Release and Ammonia-Oxidizing Bacteria Abundance in Sediments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Microcosm Set up
2.2. Preparation of Peeper and Sampling
2.3. Analytical Methods
3. Results
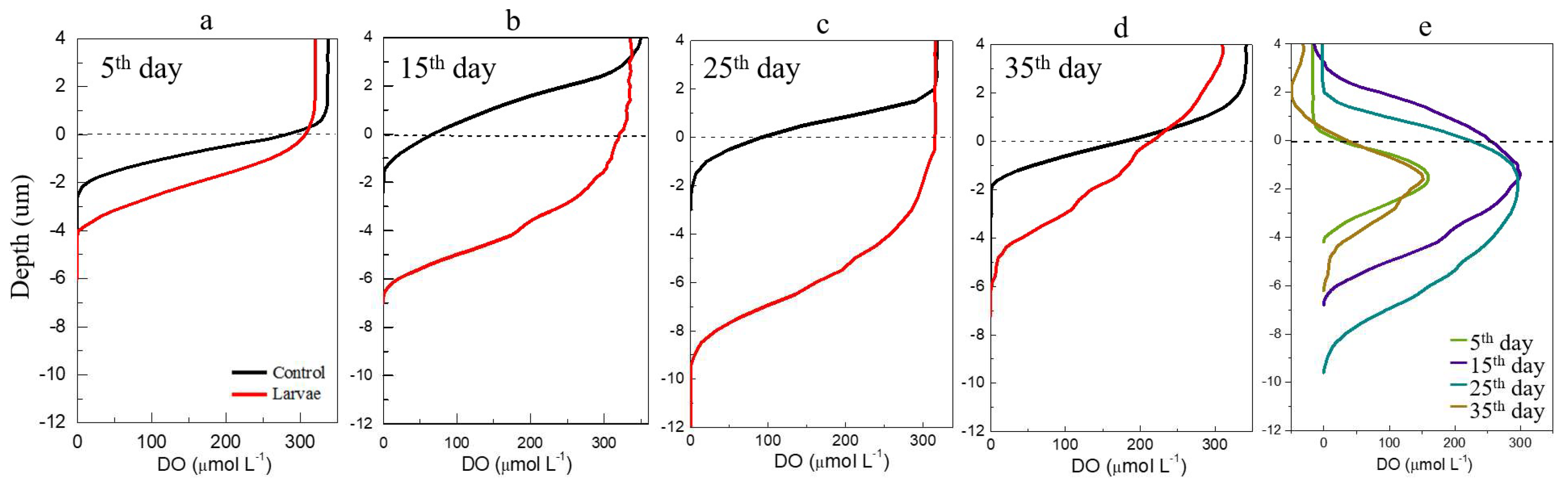
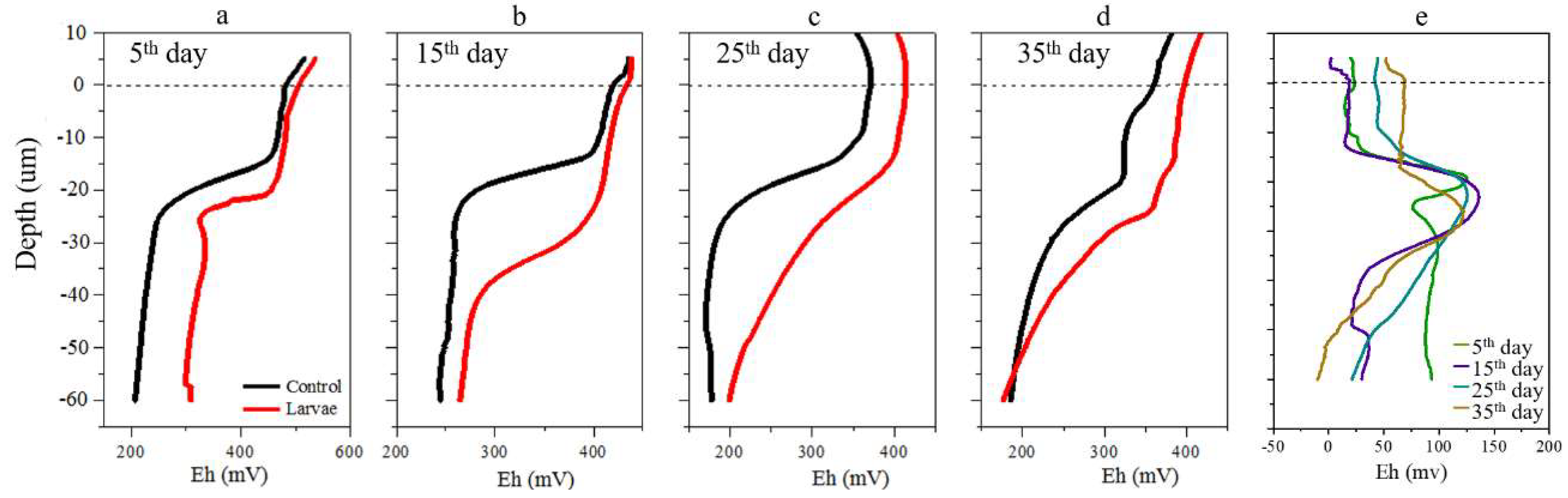
3.1. Changes of DO and Eh in Sediment Cores
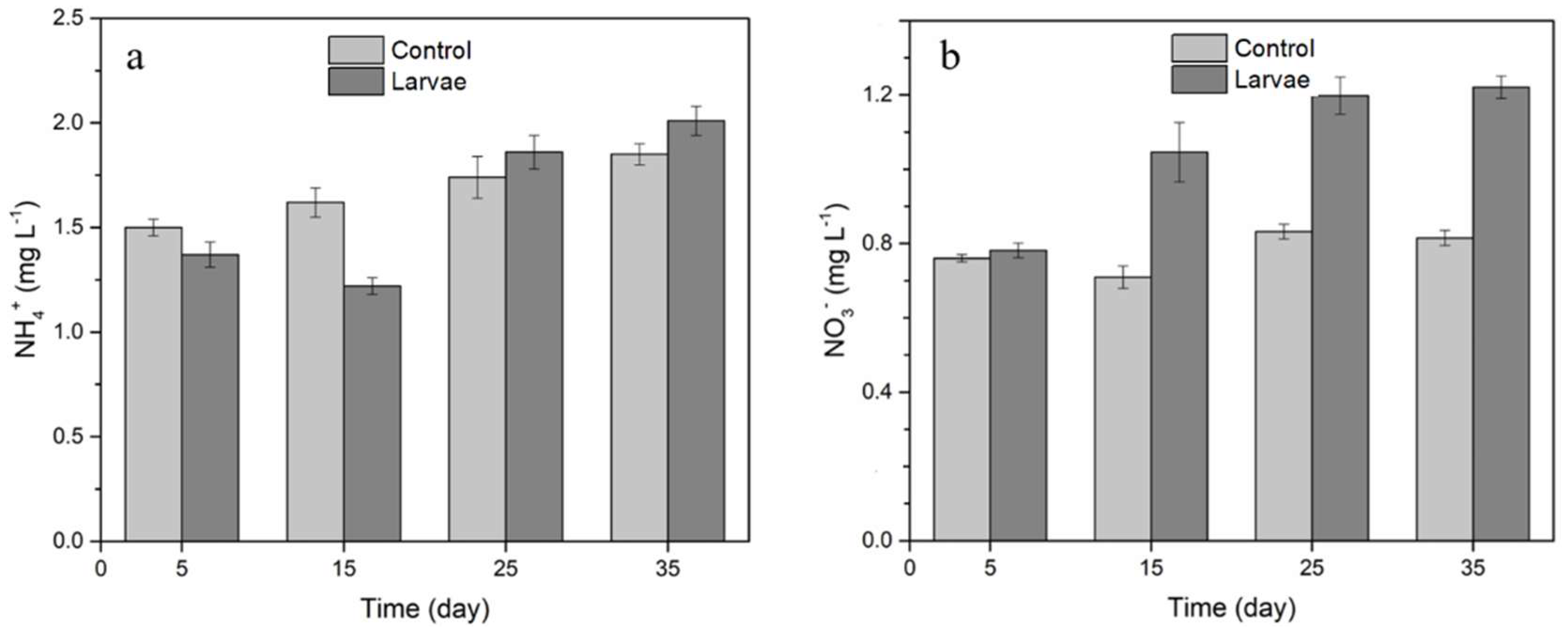
3.2. Changes of NH4+ and NO3− in Overlying Water
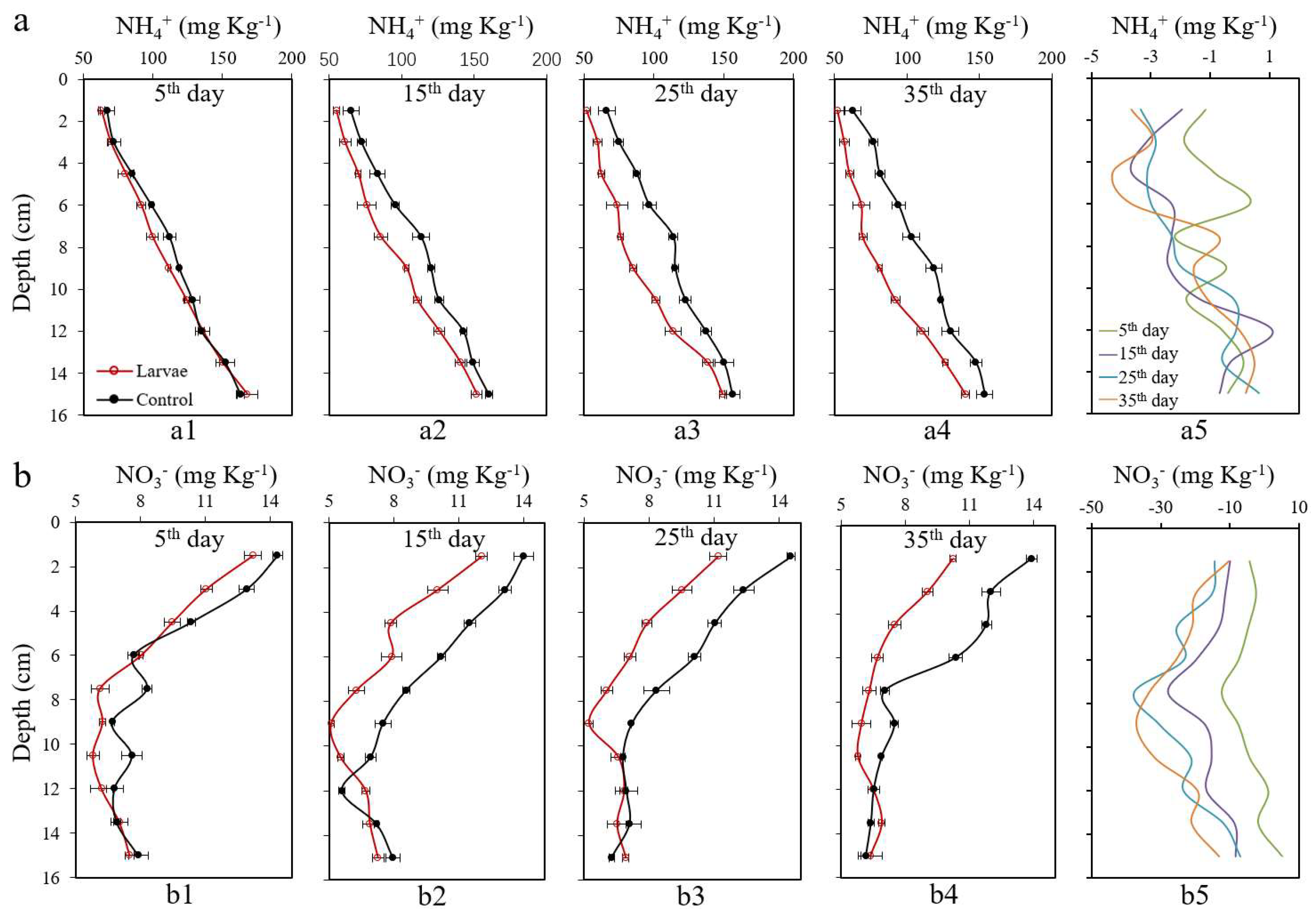
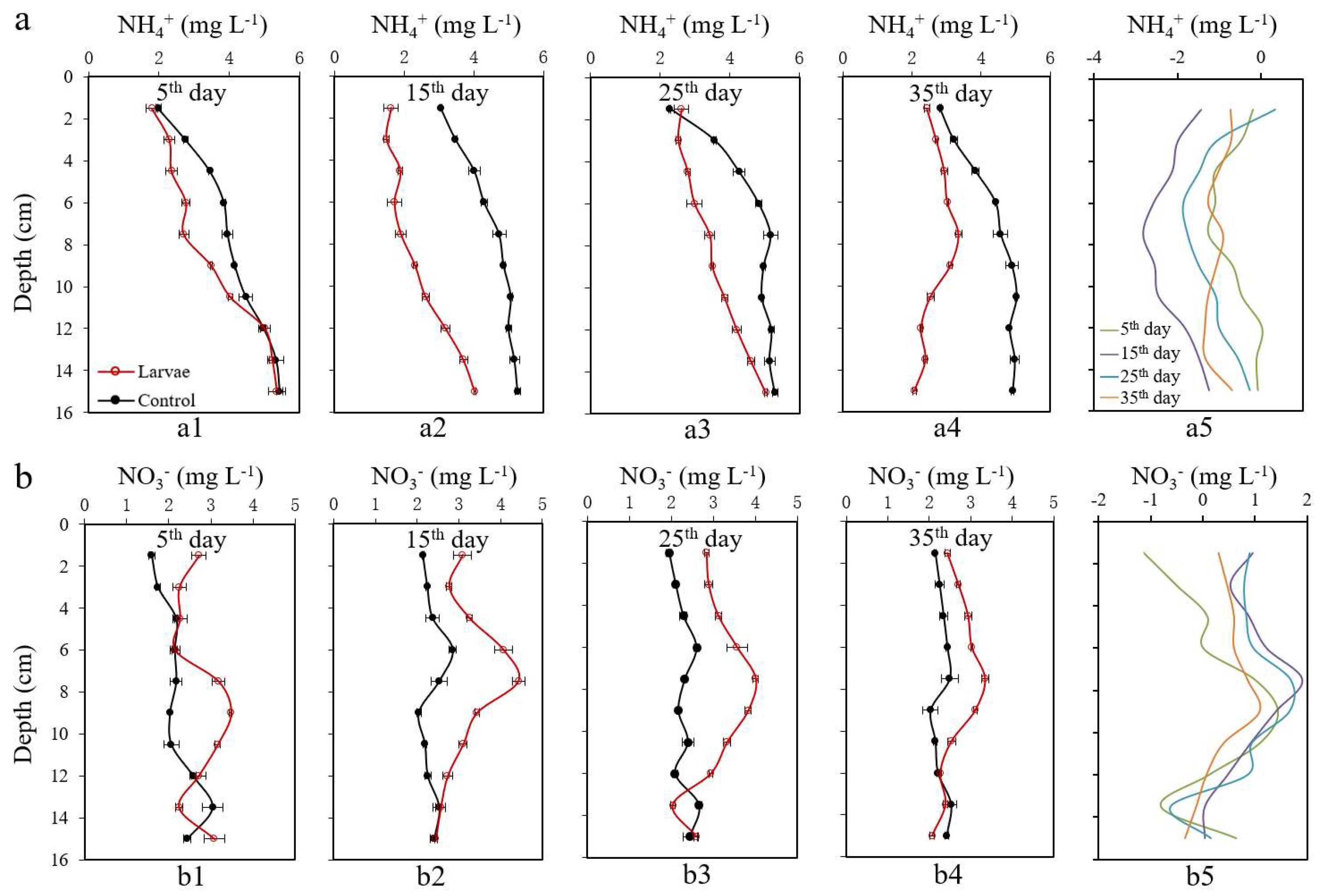
3.3. Changes of NH4+ and NO3− in Sediments
3.4. Changes of NH4+ and NO3− in Pore Water
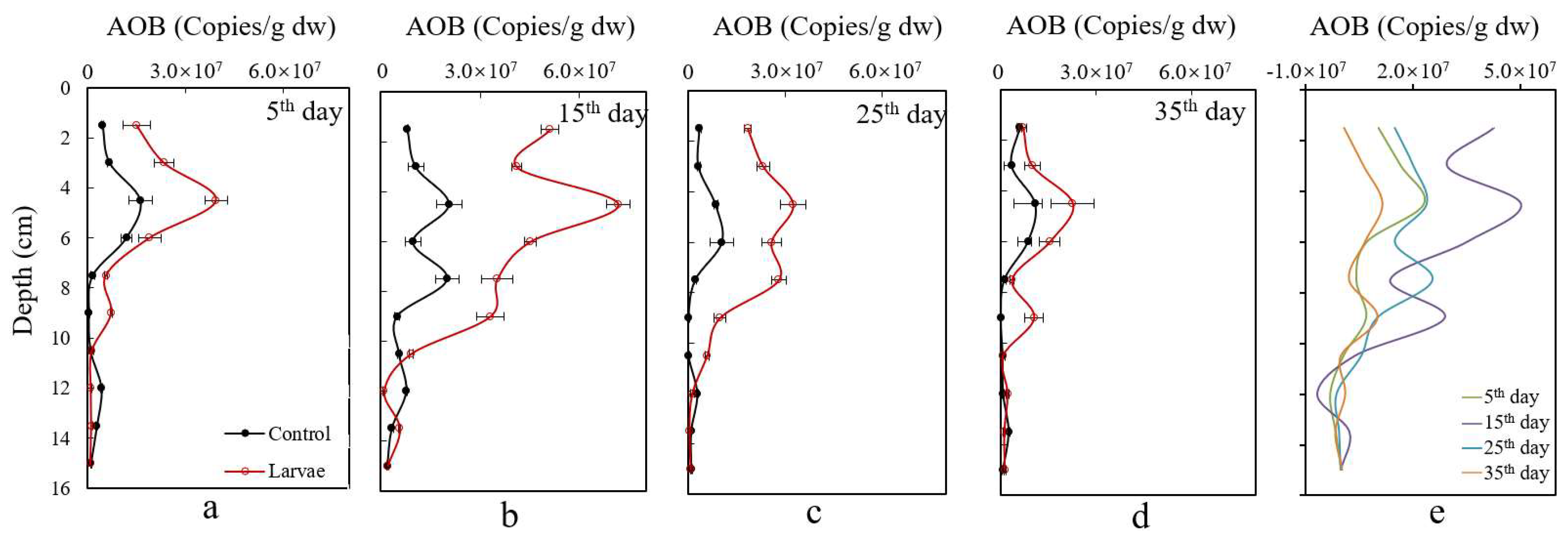
3.5. Changes of Ammonia-Oxidizing Bacteria in Sediments
4. Discussion
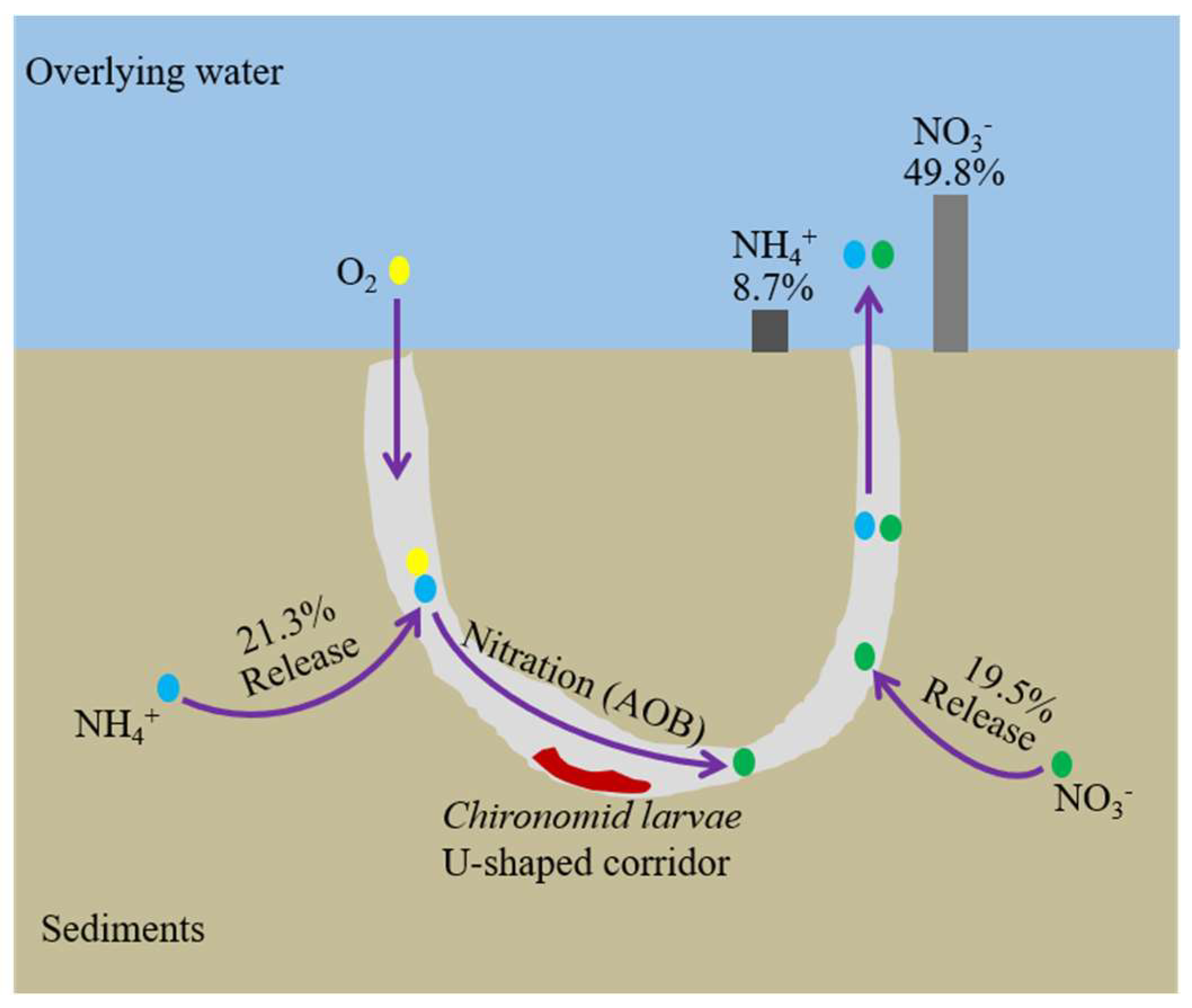
4.1. Assessment of the Effects of the Larvae Bioturbation on Sediment N
4.2. Bioturbation Mechanism on Sediment N
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hou, D.; He, J.; Lü, C.; Dong, S.; Wang, J.; Xie, Z.; Zhang, F. Spatial variations and distributions of phosphorus and nitrogen in bottom sediments from a typical north-temperate lake, China. Environ. Earth Sci. 2014, 71, 3063–3079. [Google Scholar] [CrossRef]
- Qin, B.; Zhu, G.; Lu, Z.; Luo, L.; Gao, G.; Gu, B. Estimation of internal nutrient release in large shallow Lake Taihu, China. Sci. China Earth Sci. 2006, 49, 38–50. [Google Scholar] [CrossRef]
- Qin, B.; Weiping, H.U.; Gao, G. Dynamics of sediment resus-pension and the conceptual schema of nutrient release in the large shallow Lake Taihu, China. Chin. Sci. Bull. 2004, 49, 54–64. [Google Scholar] [CrossRef]
- Danielsson, A.; Jönsson, A.; Rahm, L. Resuspension patterns in the Baltic proper. J. Sea Res. 2007, 57, 257–269. [Google Scholar] [CrossRef]
- Spears, B.M.; Carvalho, L.; Perkins, R.; Paterson, D.M. Effects of light on sediment nutrient flux and water column nutrient stoichiometry in a shallow lake. Water Res. 2008, 42, 977. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.; Hupfer, M. Effect of Macrozoobenthos on Two-Dimensional Small-Scale Heterogeneity of Pore Water Phosphorus Concentrations in Lake Sediments: A Laboratory Study. Limnol. Oceanogr. 2005, 50, 1106–1118. [Google Scholar] [CrossRef]
- Hansen, K.; Mouridsen, S.; Kristensen, E. The impact of Chironomus plumosus larvae on organic matter decay and nutrient (N, P) exchange in a shallow eutrophic lake sediment following a phytoplankton sedimentation. Hydrobiologia 1997, 364, 65–74. [Google Scholar] [CrossRef]
- Janssen, F.; Huettel, M.; Witte, U. Pore-water advection and solute fluxes in permeable marine sediments (II): Benthic respiration at three sandy sites with different permeabilities (German Bight, North Sea). Limnol. Oceanogr. 2005, 50, 779–792. [Google Scholar] [CrossRef]
- Meysman, F.J.R.; Galaktionov, O.S.; Britta, G.; Middelburg, J.J. Bioirrigation in permeable sediments: Advective pore water transport induced by burrow ventilation. Limnol. Oceanogr. 2006, 51, 142–156. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Q.; Gu, X.; He, W.; Zhang, Z.; Fan, C. Oxygen and phosphorus dynamics in freshwater sediment after the deposition of flocculated cyanobacteria and the role of tubificid worms. J. Hazard. Mater. 2014, 266, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Han, C.; Wang, Y.; Yao, L.; Wang, Y.; Xu, D.; Williams, P.N.; Zhang, C. In situ, high-resolution imaging of labile phosphorus in sediments of a large eutrophic lake. Water Res. 2015, 74, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, H.; Sakamoto, M. Enhancement of Inorganic Nitrogen and Phosphate Release from Lake Sediment by Tubificid Worms and Chironomid Larvae. Oikos 1987, 48, 312–320. [Google Scholar] [CrossRef]
- Andersen, F.; Jørgensen, M.; Jensen, H.S. The Influence of Chironomus plumosus Larvae on Nutrient Fluxes and Phosphorus Fractions in Aluminum Treated Lake Sediment. Water, Air, Soil Pollut. 2006, 6, 101–110. [Google Scholar] [CrossRef]
- Reitzel, K.; Lotter, S.; Dubke, M.; Egemose, S.; Jensen, H.S.; Andersen, F.Ø. Effects of Phoslock ® treatment and chironomids on the exchange of nutrients between sediment and water. Hydrobiologia 2013, 703, 189–202. [Google Scholar] [CrossRef]
- Tátrai, I. Experiments on Nitrogen and Phosphorus Release by Chironomus ex gr. plumosus from the Sediments of Lake Balaton, Hungary. Int. Rev. Hydrobiol. 2010, 73, 627–640. [Google Scholar]
- Granéli, W. The influence of Chironomus plumosus larvae on the exchange of dissolved substances between sediment and water. Hydrobiologia 1979, 66, 149–159. [Google Scholar] [CrossRef]
- Svensson, J.; Leonardson, L. Effects of bioturbation by tube-dwelling chironomid larvae on oxygen uptake and denitrification in eutrophic lake sediments. Plant J. 2010, 35, 289–300. [Google Scholar] [CrossRef]
- Laverock, B.; Kitidis, V.; Tait, K.; Gilbert, J.A.; Osborn, A.M.; Widdicombe, S. Bioturbation determines the response of benthic ammonia-oxidizing microorganisms to ocean acidification. Philos. Trans. R. Soc. Lond. 2013, 368, 20120441. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wu, W.; Ding, S.; Sun, Q.; Zhang, C. A high-resolution dialysis technique for rapid determination of dissolved reactive phosphate and ferrous iron in pore water of sediments. Sci. Total Environ. 2012, 422, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Revsbech, N.P. Analysis of microbial communities with electrochemical microsensors and microscale biosensors. Methods Enzymol. 2005, 397, 147. [Google Scholar] [PubMed]
- Revsbech, N.P.; Risgaard-Petersen, N.; Schramm, A.; Nielsen, L.P. Nitrogen transformations in stratified aquatic microbial ecosystems. Antonie Van Leeuwenhoek 2006, 90, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Hesslein, R.H. An in situ sampler for close interval pore water studies. Limnol. Oceanogr. 1976, 21, 912–914. [Google Scholar] [CrossRef]
- Mayer, L.M. Chemical Water Sampling in Lakes and Sediments with Dialysis Bags. Limnol. Oceanogr. 1976, 21, 909–912. [Google Scholar] [CrossRef]
- Mosier, A.C.; Francis, C.A. Relative abundance and diversity of ammonia-oxidizing archaea and bacteria in the San Francisco Bay estuary. Environ. Microbiol. 2008, 10, 3002. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Ding, S.; Liu, L.; Xu, D.; Han, C.; Zhang, C. Iron-coupled inactivation of phosphorus in sediments by macrozoobenthos (chironomid larvae) bioturbation: Evidences from high-resolution dynamic measurements. Environ. Pollut. 2015, 204, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Aller, R.C.; Aller, J.Y. The effect of biogenic irrigation intensity and solute exchange on diagenetic reaction rates in marine sediments. J. Mar. Res. 1998, 56, 905–936. [Google Scholar] [CrossRef]
- Walshe, B.M. Feeding Mechanism of Chironomus larvae. Nature 1947, 160, 474. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.; Laskov, C.; Hupfer, M. The relationship between Chironomus plumosus burrows and the spatial distribution of pore-water phosphate, iron and ammonium in lake sediments. Freshw. Biol. 2007, 52, 331–343. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Kemp, W.M. Influence of the submersed plant, Potamogeton perfoliatus, on nitrogen cycling in estuarine sediments. Limnol. Oceanogr. 1992, 37, 1483–1495. [Google Scholar] [CrossRef]
- Roskosch, A.; Hette, N.; Hupfer, M.; Lewandowski, J. Alteration of Chironomus plumosus ventilation activity and bioirrigation-mediated benthic fluxes by changes in temperature, oxygen concentration, and seasonal variations. Freshw. Sci. 2012, 31, 269–281. [Google Scholar] [CrossRef]
- Charpentier, J.; Martin, G.; Wacheux, H.; Gilles, P. ORP regulation and Activated Sludge: 15 years of experience. Water Sci. Technol. 1998, 38, 197–208. [Google Scholar]
- Li, B.; Bishop, P. Oxidation-reduction potential (ORP) regulation of nutrient removal in activated sludge wastewater treatment plants. Water Sci. Technol. 2002, 46, 35–38. [Google Scholar] [PubMed]
- Bertics, V.; Sohm, J.; Treude, T.; Chow, C.E.T.; Capone, D.G.; Fuhrman, J.A.; Ziebis, W. Burrowing deeper into benthic nitrogen cycling: The impact of bioturbation on nitrogen fixation coupled to sulfate reduction. Mar. Ecol. Progress 2010, 409, 1–15. [Google Scholar] [CrossRef]
- Pelegrí, S.P.; Blackburn, T.H. Nitrogen cycling in lake sediments bioturbated by Chironomus plumosus larvae, under different degrees of oxygenation. Hydrobiologia 1996, 325, 231–238. [Google Scholar] [CrossRef]
- Svensson, J.M. Influence of Chironomus plumosus larvae on ammonium flux and denitrification (measured by the acetylene blockage- and the isotope pairing-technique) in eutrophic lake sediment. Hydrobiologia 1997, 346, 157–168. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sediment Layer (cm) | Cay (%) | Silt (%) | Sand (%) | NO3− (mg·kg−1) | NH4+ (mg·kg−1) | TN (mg·kg−1) |
|---|---|---|---|---|---|---|
| 0–4 | 2.69 | 43.12 | 54.19 | 12.92 | 67.18 | 2412.66 |
| 4–8 | 3.04 | 42.60 | 54.36 | 6.32 | 99 | 2074.7 |
| 8–12 | 3.39 | 44.34 | 52.26 | 6.52 | 128.44 | 1823.36 |
| 12–16 | 6.37 | 49.68 | 43.96 | 7.37 | 152.07 | 1694.65 |
| Treatment | Time (day) | r |
|---|---|---|
| Chironomid larvae | 5 | 0.477 |
| 15 | −0.612 ** | |
| 25 | −0.411 * | |
| 35 | −0.570 * | |
| Control | 5 | 0.181 |
| 15 | 0.163 | |
| 25 | 0.233 | |
| 35 | 0.199 |
| Items | Time (day) | r |
|---|---|---|
| NH4+ vs. AOB | 5 | −0.731 * |
| 15 | −0.838 ** | |
| 25 | −0.843 ** | |
| 35 | −0.497 | |
| NO3− vs. AOB | 5 | 0.655 |
| 15 | 0.785 ** | |
| 25 | 0.711 * | |
| 35 | 0.632 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, X.; Liu, L.; Yan, W.; Wu, T.; Zhao, L.; Wang, X. Bioturbation Effects of Chironomid Larvae on Nitrogen Release and Ammonia-Oxidizing Bacteria Abundance in Sediments. Water 2018, 10, 512. https://doi.org/10.3390/w10040512
Xing X, Liu L, Yan W, Wu T, Zhao L, Wang X. Bioturbation Effects of Chironomid Larvae on Nitrogen Release and Ammonia-Oxidizing Bacteria Abundance in Sediments. Water. 2018; 10(4):512. https://doi.org/10.3390/w10040512
Chicago/Turabian StyleXing, Xigang, Ling Liu, Wenming Yan, Tingfeng Wu, Liping Zhao, and Xixi Wang. 2018. "Bioturbation Effects of Chironomid Larvae on Nitrogen Release and Ammonia-Oxidizing Bacteria Abundance in Sediments" Water 10, no. 4: 512. https://doi.org/10.3390/w10040512
APA StyleXing, X., Liu, L., Yan, W., Wu, T., Zhao, L., & Wang, X. (2018). Bioturbation Effects of Chironomid Larvae on Nitrogen Release and Ammonia-Oxidizing Bacteria Abundance in Sediments. Water, 10(4), 512. https://doi.org/10.3390/w10040512