Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake

Abstract
:1. Introduction
2. Methods
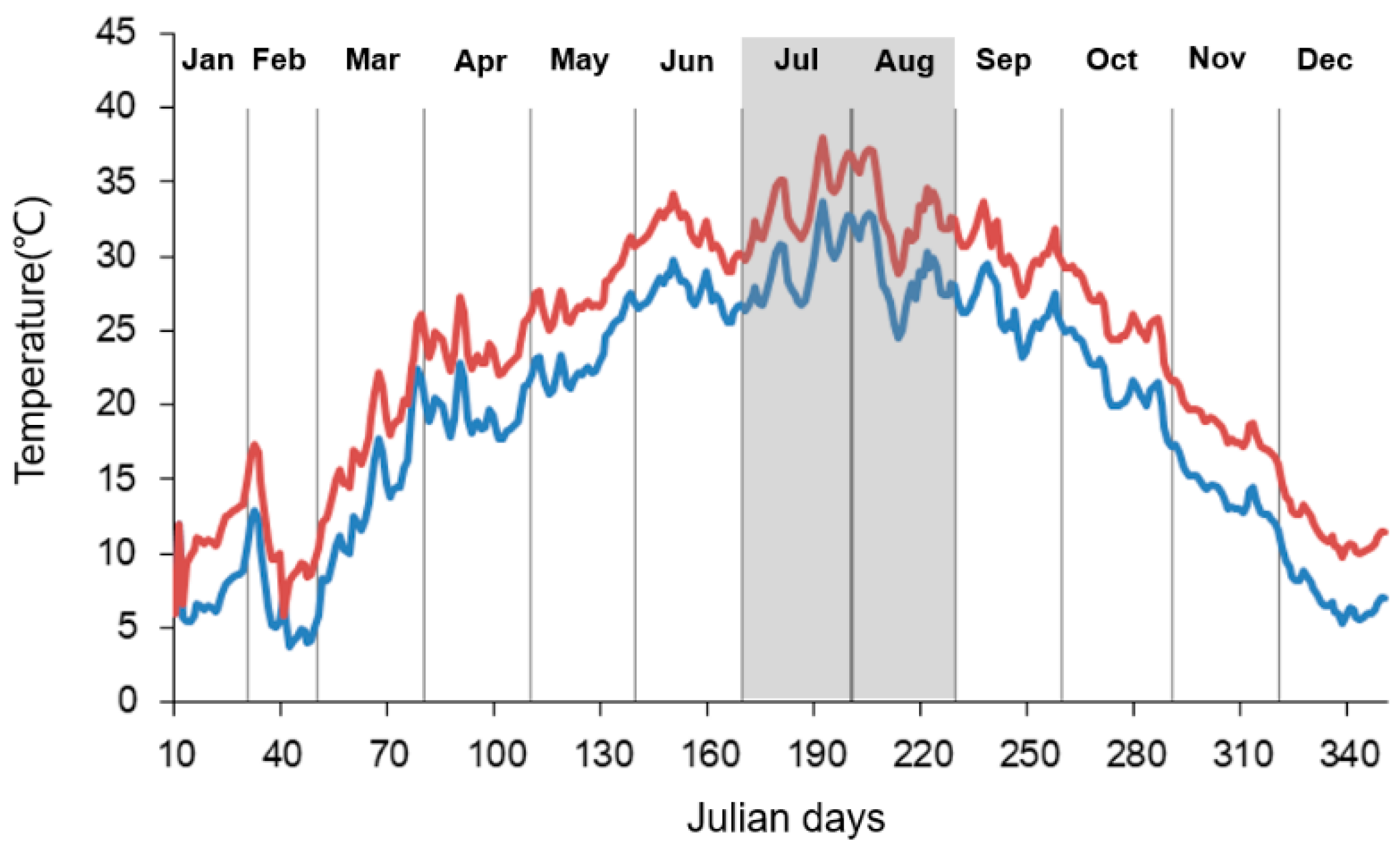
2.1. Mesocosm Establishment
2.2. Sampling and Analysis
2.3. Nitrogen Uptake Experiments
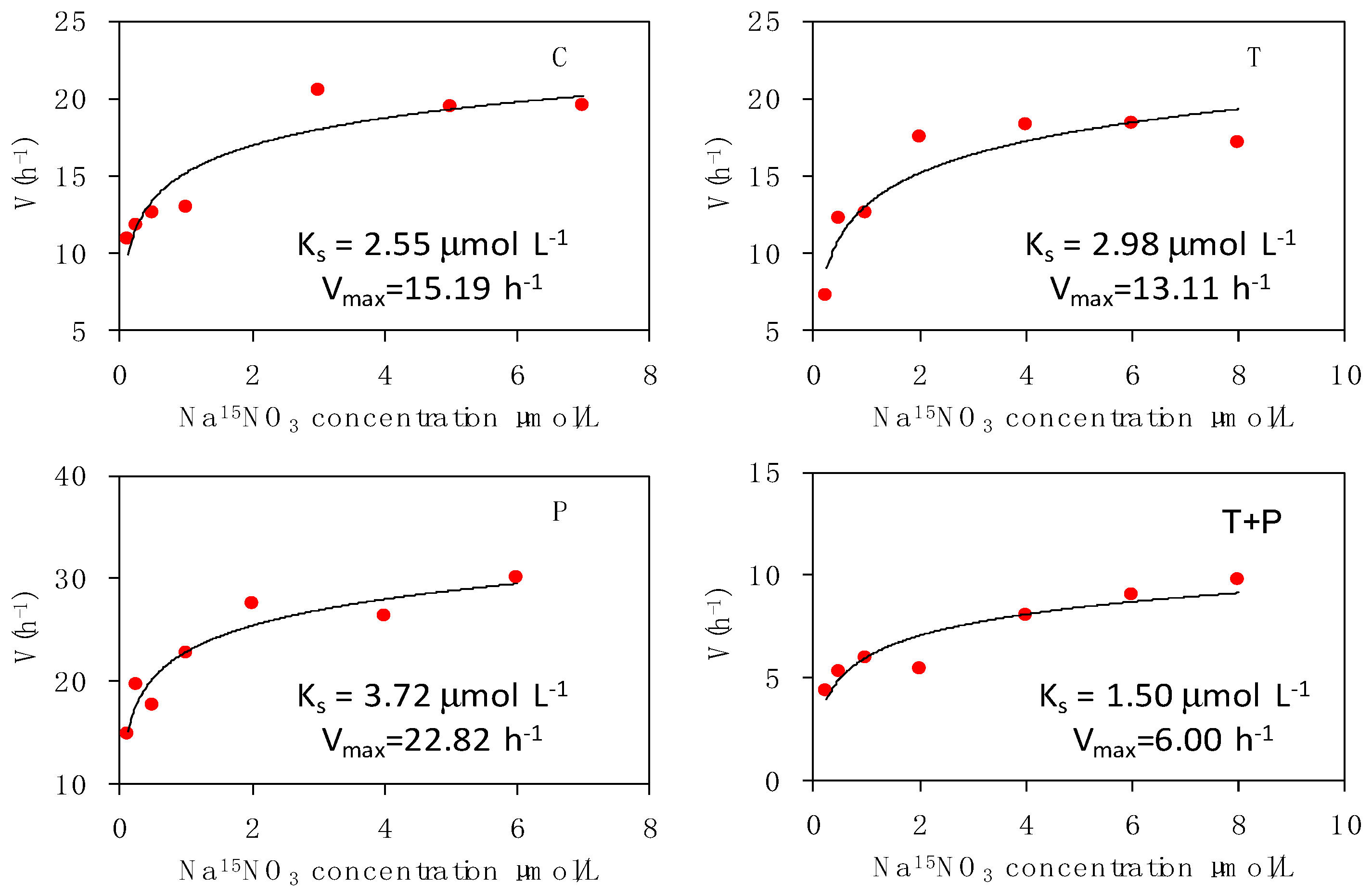
2.4. Kinetics of Nitrogen Uptake
2.5. Tracer Techniques
3. Results
3.1. Water Chemistry in Mesocosms
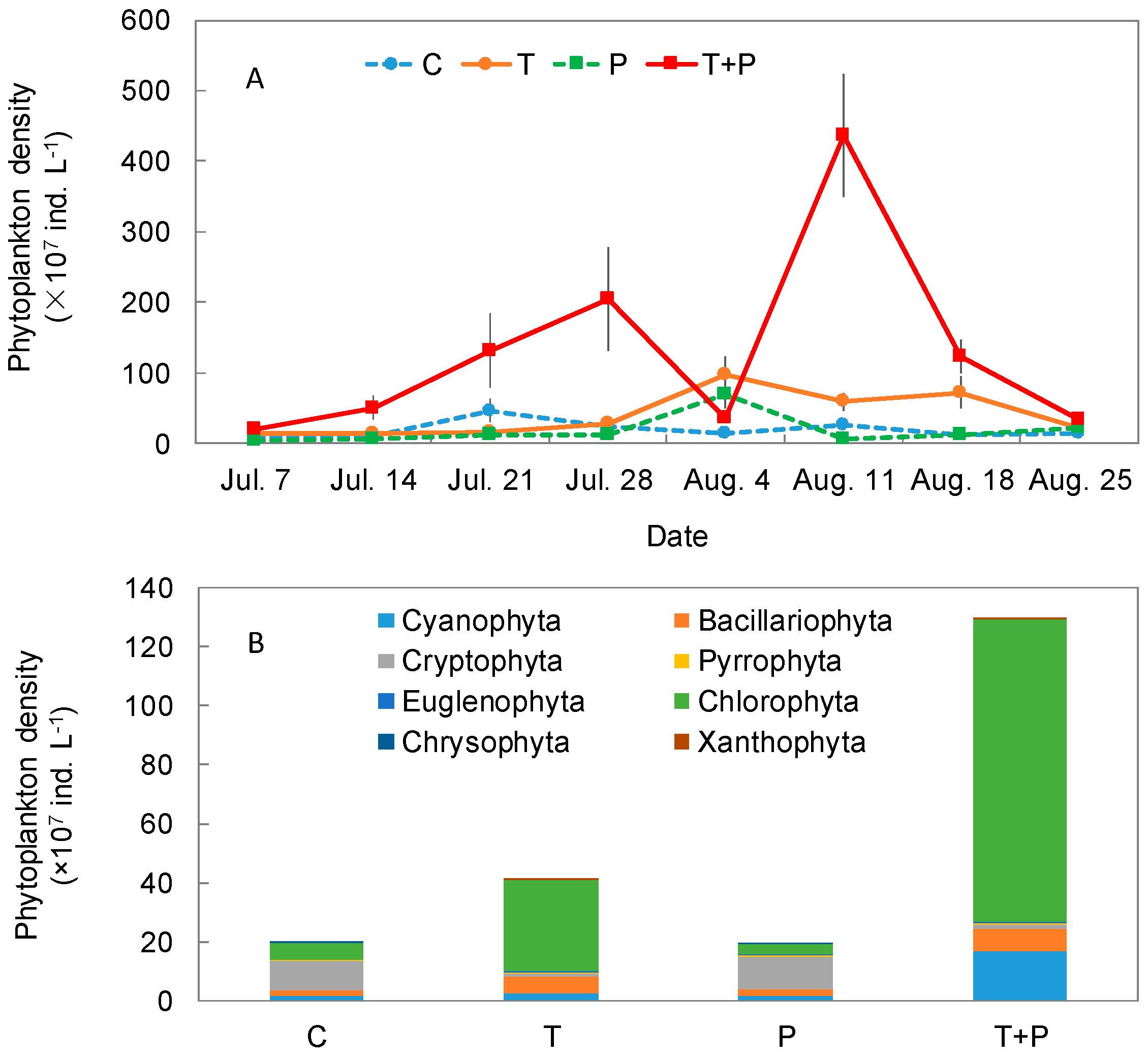
3.2. Phytoplankton Community Structure
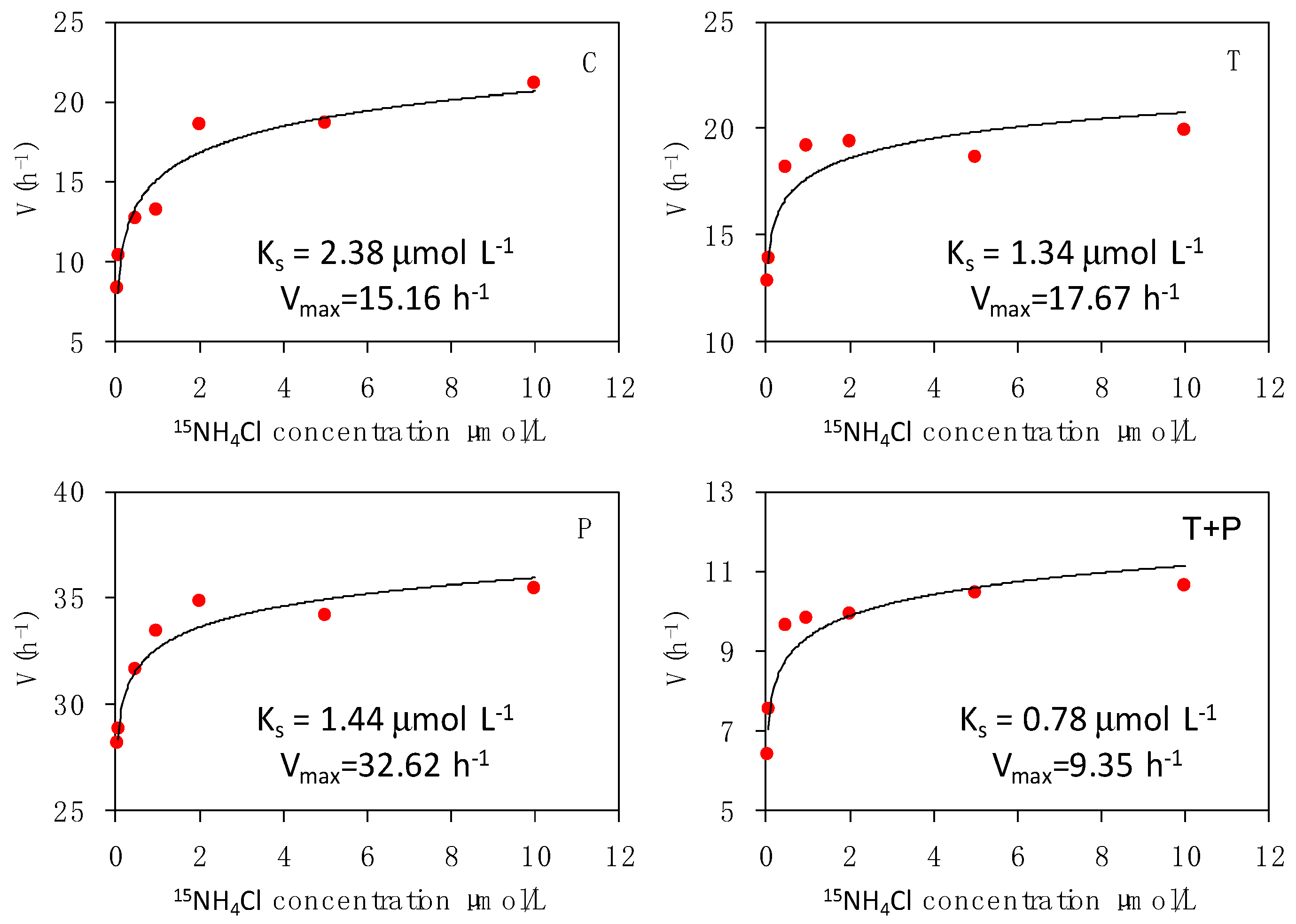
3.3. Effects on Dynamics of Phytoplankton Nitrogen Uptake
4. Discussion
4.1. Effects of Nutrients and Temperature on Phytoplankton Community Structure
4.2. Effects on Dynamics of Phytoplankton Nitrogen Uptake
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blenckner, T. A conceptual model of climate-related effects on lake ecosystems. Hydrobiologia 2005, 533, 1–14. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change (IPCC). Climate Change 2014: Synthesis Report. In Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; p. 8. [Google Scholar]
- Meerhoff, M.; Clemente, J.M.; Mello, F.T.D.; Iglesias, C.; Pedersen, A.R.; Jeppesen, E. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes? J. Glob. Chang. Biol. 2007, 13, 1888–1897. [Google Scholar] [CrossRef]
- Müren, U.; Berglund, J.; Samuelsson, K.; Andersson, A. Potential Effects of Elevated Sea-Water Temperature on Pelagic Food Webs. Hydrobiologia 2005, 545, 153–166. [Google Scholar] [CrossRef]
- O’Connor, M.I.; Piehler, M.F.; Leech, D.M.; Anton, A.; Bruno, J.F. Warming and Resource Availability Shift Food Web Structure and Metabolism. J. PLoS Biol. 2009, 7. [Google Scholar] [CrossRef]
- Markensten, H. Climate Effects on Early Phytoplankton Biomass Over Three Decades Modified by the Morphometry in Connected Lake Basins. Hydrobiologia 2006, 559, 319–329. [Google Scholar] [CrossRef]
- Blenckner, T.; Chen, D.L. Comparison of the impact of regional and North Atlantic atmospheric circulation on an aquatic ecosystem. J. Clim. Res. 2003, 23, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Lengfellner, K. Climate change and the timing, magnitude, and composition of the phytoplankton spring bloom. J. Glob. Chang. Biol. 2008, 14, 1199–1208. [Google Scholar] [CrossRef]
- Klauschies, T.; Bauer, B.; Aberle-Malzahn, N.; Sommer, U.; Gaedke, U. Climate change effects on phytoplankton depend on cell size and food web structure. J. Mar. Biol. 2012, 159, 2455–2478. [Google Scholar] [CrossRef] [Green Version]
- Domine, L.M.; Vanni, M.J.; Renwick, W.H. New and regenerated primary production in a productive reservoir ecosystem. J. Can. J. Fish. Aquat. Sci. 2010, 67, 278–287. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Olesen, J.E.; Audet, J.; Søndergaard, M.; Hoffmann, C.C.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Larsen, S.E.; et al. Climate change effects on nitrogen loading from cultivated catchments in Europe: Implications for nitrogen retention, ecological state of lakes and adaptation. Hydrobiologia 2011, 663, 1–21. [Google Scholar] [CrossRef]
- Domis, L.N.; Van de Waal, D.B.; Helmsing, N.R.; Van Donk, D.E.; Mooij, W.M. Community stoichiometry in a changing world: Combined effects of warming and eutrophication on phytoplankton dynamics. Ecology 2014, 95, 1485–1495. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W. Recent advances in the understanding and management of eutrophication. J. Limnol. Oceanogr. 2006, 51, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Smith, V.H.; Joye, S.B.; Howarth, R.W. Eutrophication of Freshwater and Marine Ecosystems. J. Limnol. Oceanogr. 2006, 51, 351–355. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Ishikawa, K.; Sakai, Y.; Ishikawa, T.; Ichise, S.; Yamamoto, Y.; Kuo, T.C.; Park, H.D.; Yamamura, N.; Kumagai, M. Phytoplankton community reorganization driven by eutrophication and warming in Lake Biwa. J. Aquat. Sci. 2010, 72, 467–483. [Google Scholar] [CrossRef]
- Winder, M.; Hunter, D.A. Temporal organization of phytoplankton communities linked to physical forcing. Oecologia 2008, 156, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.A.; Jones, I.D.; Thackeray, S.J. Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake. Hydrobiologia 2006, 559, 401–411. [Google Scholar] [CrossRef]
- Kudela, R.M.; Cochlan, W.P.; Dugdale, R.C. Carbon and nitrogen uptake response to light by phytoplankton during an upwelling event. J. Plankton Res. 1997, 19, 609–630. [Google Scholar] [CrossRef] [Green Version]
- Elskens, M.; Baeyens, W.; Goeyens, L. Contribution of nitrate to the uptake of nitrogen by phytoplankton in an ocean margin environment. Hydrobiologia 1997, 353, 139–152. [Google Scholar] [CrossRef]
- Seeyave, S.; Probyn, T.; Álvarez-Salgado, X.A.; Figueiras, F.G.; Purdie, D.A.; Barton, E.D.; Lucas, M. Nitrogen uptake of phytoplankton assemblages under contrasting upwelling and downwelling conditions: The Ría de Vigo, NW Iberia. J. Estuarine Coast. Shelf Sci. 2013, 124, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tungaraza, C.; Brion, N.; Rousseau, V.; Baeyens, W.; Goeyens, L. Influence of bacterial activities on nitrogen uptake rates determined by the application of antibiotics. Oceanologia 2003, 45, 473–489. [Google Scholar]
- Reay, D.S.; Nedwell, D.B.; Priddle, J.; Ellisevans, J.C. Temperature Dependence of Inorganic Nitrogen Uptake: Reduced Affinity for Nitrate at Suboptimal Temperatures in Both Algae and Bacteria. J. Appl. Environ. Microbiol. 1999, 65, 2577–2584. [Google Scholar]
- Berg, G.M.; Balode, M.; Purina, I.; Bekere, S.; Christian, B.; Serge, M. Plankton community composition in relation to availability and uptake of oxidized and reduced nitrogen. J. Aquat. Microb. Ecol. 2003, 30, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Rhee, G.-Y.; Gotham, I.J. The effect of environmental factors on phytoplankton growth: Temperature and the interactions of temperature with nutrient limitation. J. Limnol. Oceanogr. 1981, 26, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.C.; Tu, Q.Y. Investigation Specifications for Lake Eutrophication, 2nd ed.; China Environmental Science Press: Beijing, China, 1990; pp. 114–132. [Google Scholar]
- Wilhelm, S.W.; Carberry, M.J.; Eldridge, M.L.; Poorvin, L.; Saxton, M.A.; Doblin, M.A. Marine and freshwater cyanophages in a Laurentian Great Lake: Evidence from infectivity assays and molecular analyses of g20 genes. J. Appl. Environ. Microbiol. 2006, 72, 4957–4963. [Google Scholar] [CrossRef] [PubMed]
- John, D.M.; Whitton, B.A.; Brook, A.J. Freshwater Algal Flora of the British Isles. An. Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Dugdale, R.C.; Goering, J.J. Uptake of new and regenerated forms of nitrogen in primary productivity. J. Limnol. Oceanogr. 1967, 12, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Dugdale, R.C.; Wilkerson, F.P. The Use of 15N to Measure Nitrogen Uptake in Eutrophic Oceans; Experimental Considerations. J. Limnol. Oceanogr. 1986, 31, 673–689. [Google Scholar] [CrossRef]
- Gu, B.; Alexander, V. Dissolved Nitrogen Uptake by a Cyanobacterial Bloom (Anabaena flos-aquae) in a Subarctic Lake. J. Appl. Environ. Microbiol. 1993, 59, 422–430. [Google Scholar] [CrossRef]
- Gavis, J.; Ferguson, J.F. Kinetics of Carbon Dioxide Uptake by Phytoplankton at High pH. J. Limnol. Oceanogr. 1975, 20, 211–221. [Google Scholar] [CrossRef]
- Cermeño, P.; Dutkiewicz, S.; Harris, R.P.; Follows, M.; Schofield, O.; Falkowski, P.G. The role of nutricline depth in regulating the ocean carbon cycle. Proc. Natl. Acad. Sci. USA 2008, 105, 20344–20349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. J. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef]
- Hays, G.C.; Richardson, A.J.; Robinson, C. Climate change and marine plankton. J. Trends Ecol. Evol. 2005, 20, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Daufresne, M.; Lengfellner, K.; Sommer, U. Global warming benefits the small aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12788–12793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morán, X.A.G.; Lopez-Urrutia, A.; Calvo-Diaz, A.; Li, W.K.W. Increasing importance of small phytoplankton in a warmer ocean. J. Glob. Chang. Biol. 2010, 16, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Mooij, W.M.; Hülsmann, S.; Domis, L.N.D.S.; Nolet, B.A.; Bodelier, P.L.E.; Boers, P.C.M.; Pires, L.M.D.; Gons, H.J.; Ibelings, B.W.; Noordhuis, R.; et al. The impact of climate change on lakes in The Netherlands: A review. J. Aquat. Ecol. 2005, 39, 381–400. [Google Scholar] [CrossRef]
- Moss, B.; Mckee, D.; Atkinson, D.; Collings, S.E.; Eaton, J.W.; Gill, A.B.; Harvey, I.; Hatton, K.; Heyes, T.; Wilson, D. How important is climate? Effects of warming, nutrient addition and fish on phytoplankton in shallow lake microcosms. J. Appl. Ecol. 2003, 40, 782–792. [Google Scholar] [CrossRef] [Green Version]
- Mckee, D.; Atkinson, D.; Collings, S.E.; Eaton, J.W.; Gill, A.B.; Harvey, I.; Hatton, K.; Heyes, T.; Wilson, D.; Moss, B. Response of freshwater microcosm communities to nutrients, fish, and elevated temperature during winter and summer. J. Limnol. Oceanogr. 2003, 48, 707–722. [Google Scholar] [CrossRef] [Green Version]
- Hanson, C.E.; Waite, A.M.; Thompson, P.A.; Pattiaratchi, C.B. Phytoplankton community structure and nitrogen nutrition in Leeuwin Current and coastal waters off the Gascoyne region of Western Australia. Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 902–924. [Google Scholar] [CrossRef]
- Sánchez, J.; Fernández-Sevilla, J.M.; Acién, F.G.; Cerón, M.C.; Pérez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. J. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Staehr, P.; Birkeland, M. Temperature acclimation of growth, photosynthesis and respiration in two mesophilic phytoplankton species. Phycologia 2006, 45, 648–656. [Google Scholar] [CrossRef]
- Costas, E.; Floresmoya, A.; Lópezrodas, V. Rapid adaptation of phytoplankters to geothermal waters is achieved by single mutations: Were extreme environments ‘Noah’s Arks’ for photosynthesizers during the Neoproterozoic ‘snowball Earth’? J. New Phytol. 2008, 180, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Huertas, I.E.; Rouco, M.; Lópezrodas, V.; Costas, E. Warming will affect phytoplankton differently: Evidence through a mechanistic approach. J. Proc. R. Soc. B Biol. 2011, 278, 3534–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffer, M.; Rinaldi, S.; Kuznetsov, Y.A.; Van Nes, E.H. Seasonal dynamics of Daphnia and algae explained as a periodically forced predator-prey system. Oikos 1997, 80, 519–532. [Google Scholar] [CrossRef]
- Post, D.M.; Palkovacs, E.P.; Schielke, E.G.; Dodson, S.I. Intraspecific variation in a predator affects community structure and cascading trophic interactions. Ecology 2008, 89, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- Ahlgren, G.; Lundstedt, L.; Brett, M.; Forsberg, C. Lipid composition and food quality of some freshwater phytoplankton for cladoceran zooplankters. J. Plankton Res. 1990, 12, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.E.; Hogue, V.E.; Wilkerson, F.P.; Dugdale, R.C. The effect of inorganic nitrogen speciation on primary production in the San Francisco Estuary. Estuarine Coast. Shelf Sci. 2012, 104–105, 91–101. [Google Scholar] [CrossRef]
- Jampeetong, A.; Brix, H.; Kantawanichkul, S. Effects of inorganic nitrogen forms on growth, morphology, nitrogen uptake capacity and nutrient allocation of four tropical aquatic macrophytes (Salvinia cucullata, Ipomoea aquatica, Cyperus involucratus and Vetiveria zizanioides). J. Aquat. Bot. 2012, 97, 10–16. [Google Scholar] [CrossRef]
- Blasco, D.; Maclsaac, J.J.; Packard, T.T.; Dugdale, R.C. Relationship between nitrate reductase and nitrate uptake in phytoplankton in the Peru upwelling region. J. Limnol. Oceanogr. 1984, 29, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Doyle, R.W. Upwelling, clone selection, and the characteristic shape of nutrient uptake curves. J. Limnol. Oceanogr. 1975, 20, 487–489. [Google Scholar] [CrossRef] [Green Version]
- Button, D.K. On the theory of control of microbial growth kinetics by limiting nutrient concentrations. J. Deep Sea Res. 1978, 25, 1163–1177. [Google Scholar] [CrossRef]
- Cochlan, W.P.; Harrison, P.J. Kinetics of nitrogen (nitrate, ammonium and urea) uptake by the picoflagellate Micromonas pusilla (Prasinophyceae). J. Exp. Mar. Biol. Ecol. 1991, 153, 129–141. [Google Scholar] [CrossRef]
- Gu, B.; Havens, K.E.; Schelske, C.L.; Rosen, B.H. Uptake of dissolved nitrogen by phytoplankton in a eutrophic subtropical lake. J. Plankton Res. 1997, 19, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Healey, F.P. Slope of the Monod equation as an indicator of advantage in nutrient competition. J. Microb. Ecol. 1980, 5, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Glibert, P.M.; Zhou, M.J. Temporal and spatial variability in nitrogen uptake kinetics during harmful dinoflagellate blooms in the East China Sea. J. Harmful Algae 2010, 9, 531–539. [Google Scholar] [CrossRef]
- Li, J.; Glibert, P.M.; Zhou, M.J.; Lu, S.H.; Lu, D.D. Relationships between nitrogen and phosphorus forms and ratios and the development of dinoflagellate blooms in the East China Sea. J. Mar. Ecol. Prog. Ser. 2009, 383, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Donald, D.B.; Bogard, M.J.; Kerri, F.; Leavitt, P.R. Comparative effects of urea, ammonium, and nitrate on phytoplankton abundance, community composition, and toxicity in hypereutrophic freshwaters. J. Limnol. Oceanogr. 2011, 56, 2161–2175. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, E.; Galvan, A. Inorganic nitrogen assimilation in Chlamydomonas. J. Exp. Bot. 2007, 58, 2279–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Glibert, P.M.; Burkholder, J.A.M. Characterization of the affinity for nitrogen, uptake kinetics, and environmental relationships for Prorocentrum minimum in natural blooms and laboratory cultures. J. Harmful Algae 2003, 2, 283–299. [Google Scholar] [CrossRef]
- Yin, K.; Harrison, P.J.; Dortch, Q. Lack of ammonium inhibition of nitrate uptake for a diatom grown under low light conditions. J. Exp. Mar. Biol. Ecol. 1998, 228, 151–165. [Google Scholar] [CrossRef]
- Bhavya, P.S.; Kumar, S.; Gupta, G.V.M.; Sudheesh, V.; Sudharma, K.V.; Varrier, D.S.; Dhanya, K.R.; Saravanane, N. Nitrogen Uptake Dynamics in a Tropical Eutrophic Estuary (Cochin, India) and Adjacent Coastal Waters. J. Estuaries Coasts 2016, 39, 54–67. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
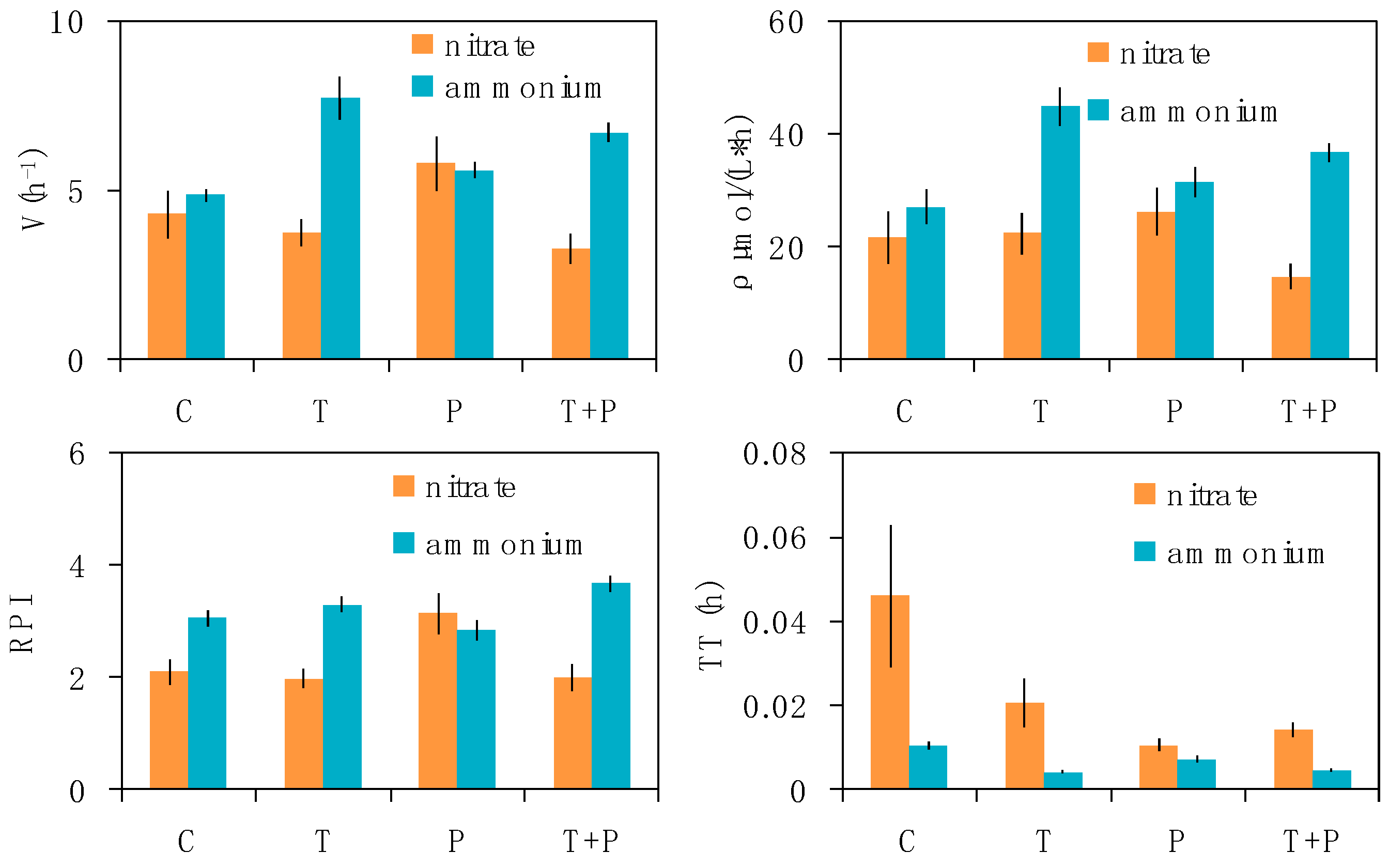{kind=link}
{kind=link}
{kind=link}
| C | T | P | T + P | |
|---|---|---|---|---|
| DO (mg L−1) | 7.61 ± 1.67 | 7.34 ± 1.85 | 7.84 ± 1.95 | 7.11 ± 2.36 |
| pH | 8.48 ± 0.55 | 8.56 ± 0.36 | 8.54 ± 0.56 | 8.56 ± 0.54 |
| Conductivity (µS cm−1) | 233.9 ± 14.7 | 308.2 ± 11.4 | 225.3 ± 15.7 | 303.4 ± 14.0 |
| TP (mg L−1) | 0.059 ± 0.018 | 0.076 ± 0.031 | 0.155 ± 0.041 | 0.275 ± 0.074 |
| SRP (mg L−1) | 0.029 ± 0.009 | 0.028 ± 0.015 | 0.115 ± 0.034 | 0.227 ± 0.059 |
| TN (mg L−1) | 0.464 ± 0.077 | 0.508 ± 0.156 | 0.470 ± 0.093 | 0.586 ± 0.099 |
| NO3−-N (mg L−1) | 0.114 ± 0.042 | 0.142 ± 0.046 | 0.112 ± 0.037 | 0.124 ± 0.033 |
| NH4+-N (mg L−1) | 0.150 ± 0.071 | 0.164 ± 0.081 | 0.198 ± 0.083 | 0.208 ± 0.098 |
| Chl-a (mg L−1) | 2.75 ± 1.21 | 5.31 ± 2.59 | 2.84 ± 1.66 | 6.53 ± 4.37 |
| Cladocera (ind. L−1) | 15.45 ± 5.10 | 23.68 ± 4.24 | 61.27 ± 21.39 | 45.18 ± 9.97 |
| Copepoda (ind. L−1) | 46.96 ± 9.46 | 31.58 ± 7.03 | 83.08 ± 14.57 | 28.13 ± 4.61 |
| Rotifera (ind. L−1) | 26.25 ± 10.02 | 49.35 ± 11.51 | 17.23 ± 3.56 | 43.83 ± 9.71 |
| N | p | Warming | P + Warming | |
|---|---|---|---|---|
| DO (mg L−1) | 48 | 0.823 | 0.784 | 0.613 |
| pH | 48 | 0.803 | 0.759 | 0.741 |
| Conductivity (µS cm−1) | 48 | 0.229 | <0.001 | <0.001 |
| TP (mg L−1) | 48 | <0.001 | 0.475 | <0.001 |
| SRP (mg L−1) | 48 | <0.001 | 0.985 | <0.001 |
| TN (mg L−1) | 48 | 0.911 | 0.427 | <0.05 |
| NO3−-N (mg L−1) | 48 | 0.927 | 0.167 | 0.620 |
| NH4+-N (mg L−1) | 48 | 0.263 | 0.745 | 0.179 |
| Chl. a (mg L−1) | 48 | 0.951 | 0.072 | <0.01 |
| Cladocera (ind. L−1) | 48 | <0.001 | <0.05 | <0.001 |
| Copepoda (ind. L−1) | 48 | <0.05 | 0.119 | 0.140 |
| Rotifera (ind. L−1) | 48 | 1.000 | 0.226 | 0.254 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.; Li, C.; Wang, T.; Zhang, M.; Xu, J. Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake. Water 2018, 10, 1057. https://doi.org/10.3390/w10081057
Yu C, Li C, Wang T, Zhang M, Xu J. Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake. Water. 2018; 10(8):1057. https://doi.org/10.3390/w10081057
Chicago/Turabian StyleYu, Chen, Chao Li, Tao Wang, Min Zhang, and Jun Xu. 2018. "Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake" Water 10, no. 8: 1057. https://doi.org/10.3390/w10081057
APA StyleYu, C., Li, C., Wang, T., Zhang, M., & Xu, J. (2018). Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake. Water, 10(8), 1057. https://doi.org/10.3390/w10081057