Why Do We Need to Document and Conserve Foundation Species in Freshwater Wetlands?


Abstract
:1. Introduction
1.1. The Importance of Foundation Species in Freshwater Wetlands
1.2. Drivers and Factors Affecting Foundation Species in Freshwater Wetlands
1.3. Foundation Species Changes and Loss in Freshwater Wetlands
1.4. Focus, Goal, and Objectives
2. Foundation Species Dynamics in Freshwater Wetlands
2.1. Foundation Species in Tropical and Subtropical Wetlands
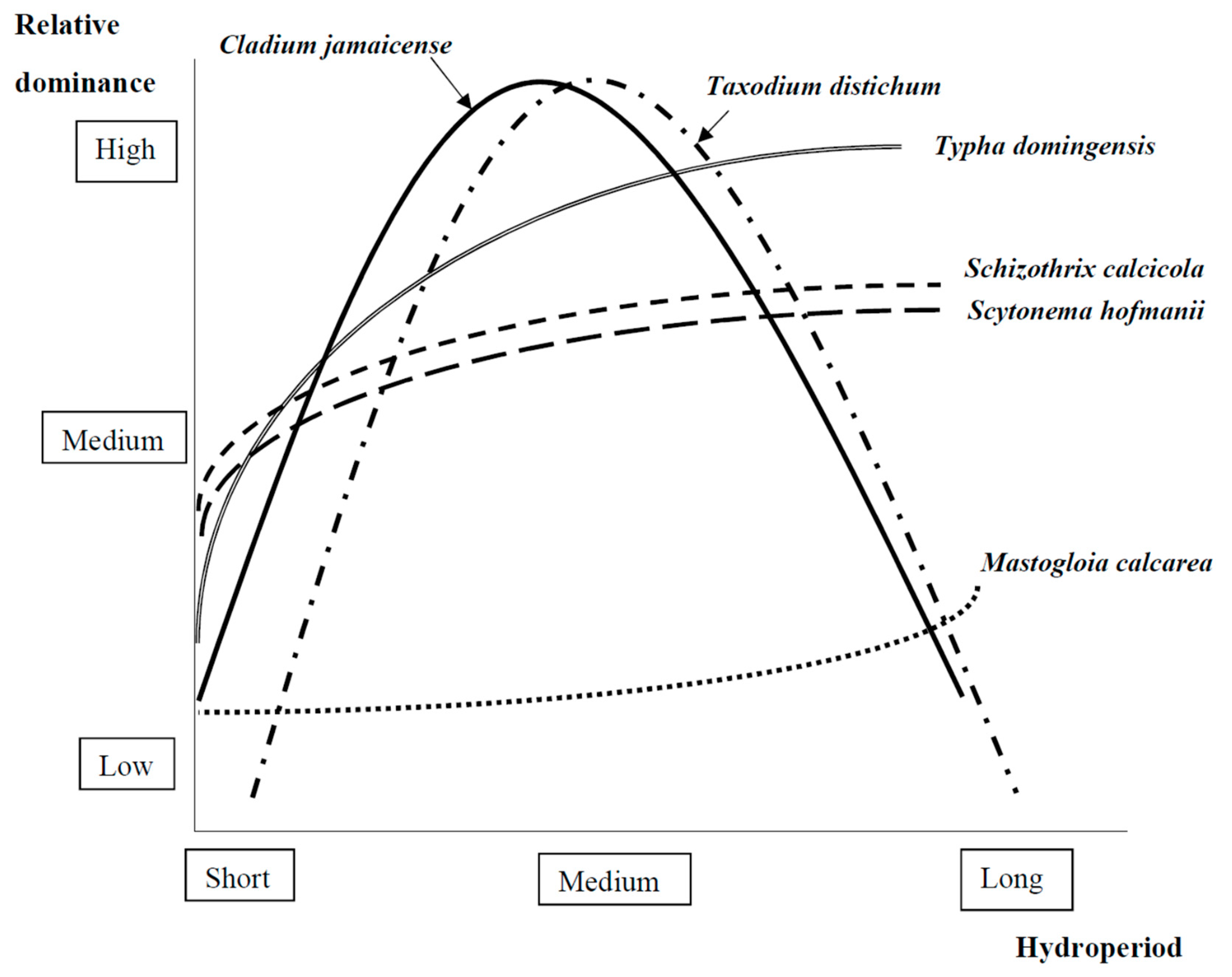
2.1.1. Foundation Species Interactions with Hydrology
Algae
Herbaceous Vascular Plants
Trees
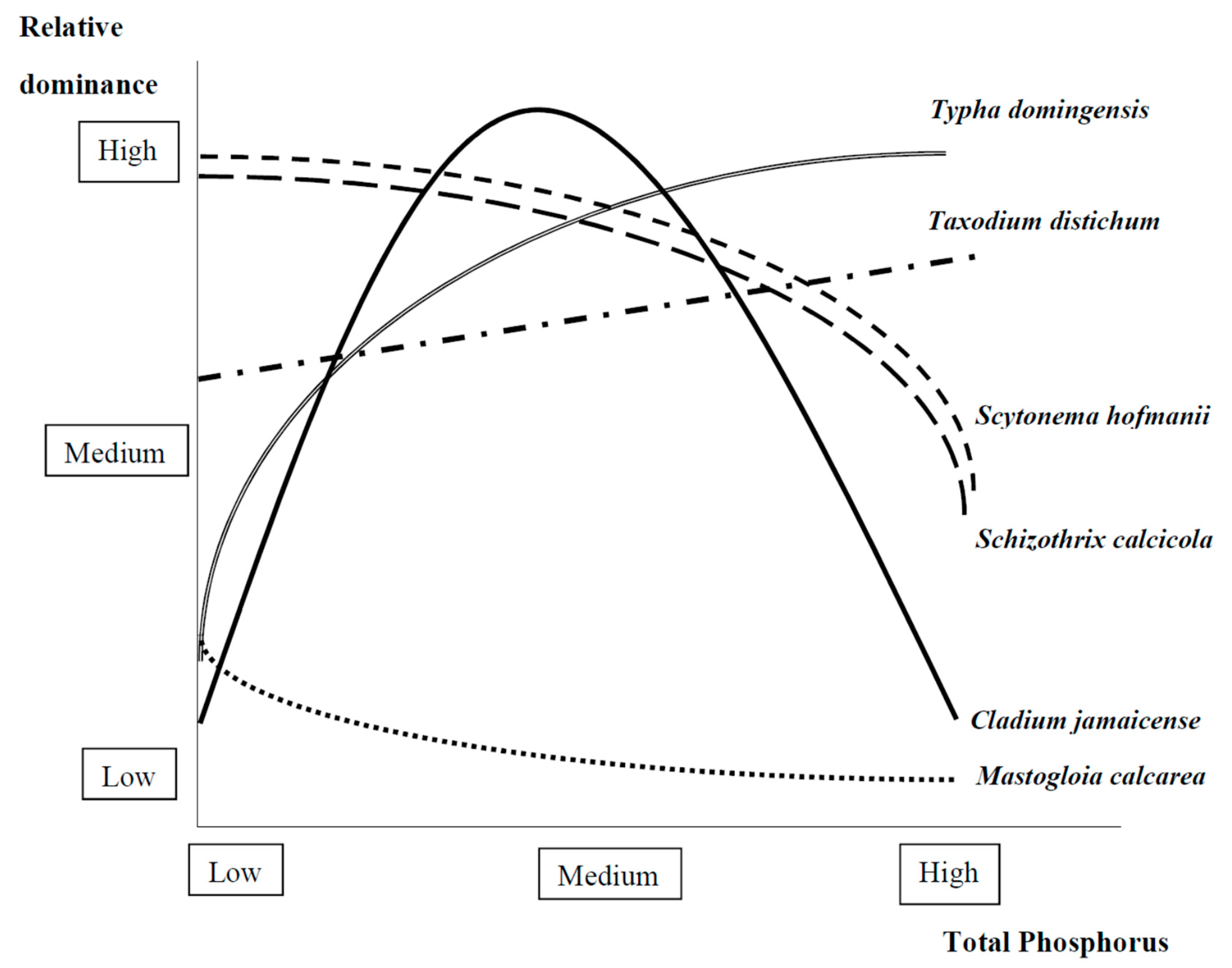
2.1.2. Foundation Species Interactions with Nutrients
Algae
Herbaceous Vascular Plants
Trees
2.1.3. Foundation Species Interactions with Other Environmental Conditions
Algae
Herbaceous Vascular Plants
Trees
2.2. Foundation Species in Boreal and Temperate Wetlands
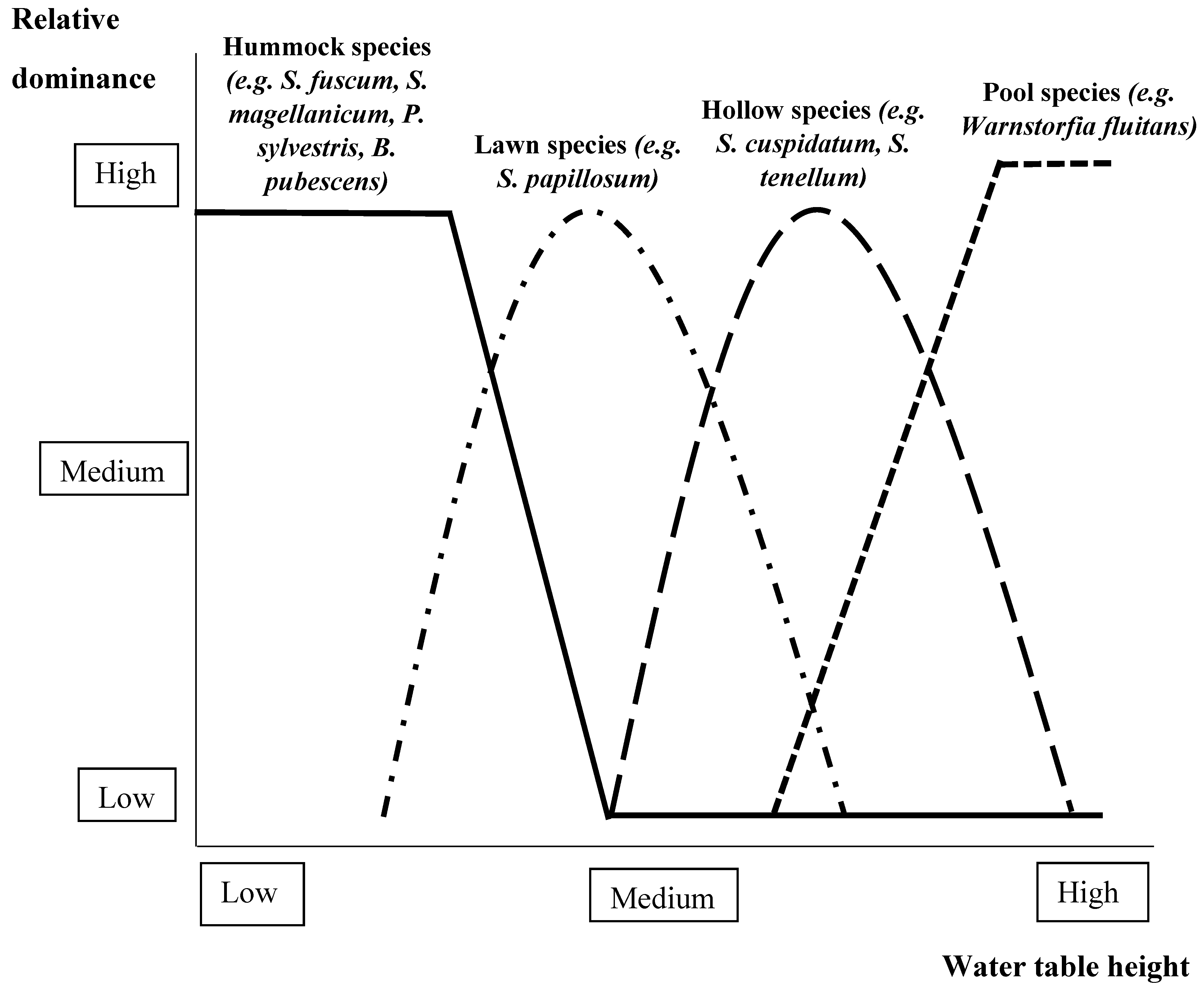
2.2.1. Foundation Species Interactions with Hydrology
Algae
Vascular and Nonvascular Plants
Trees
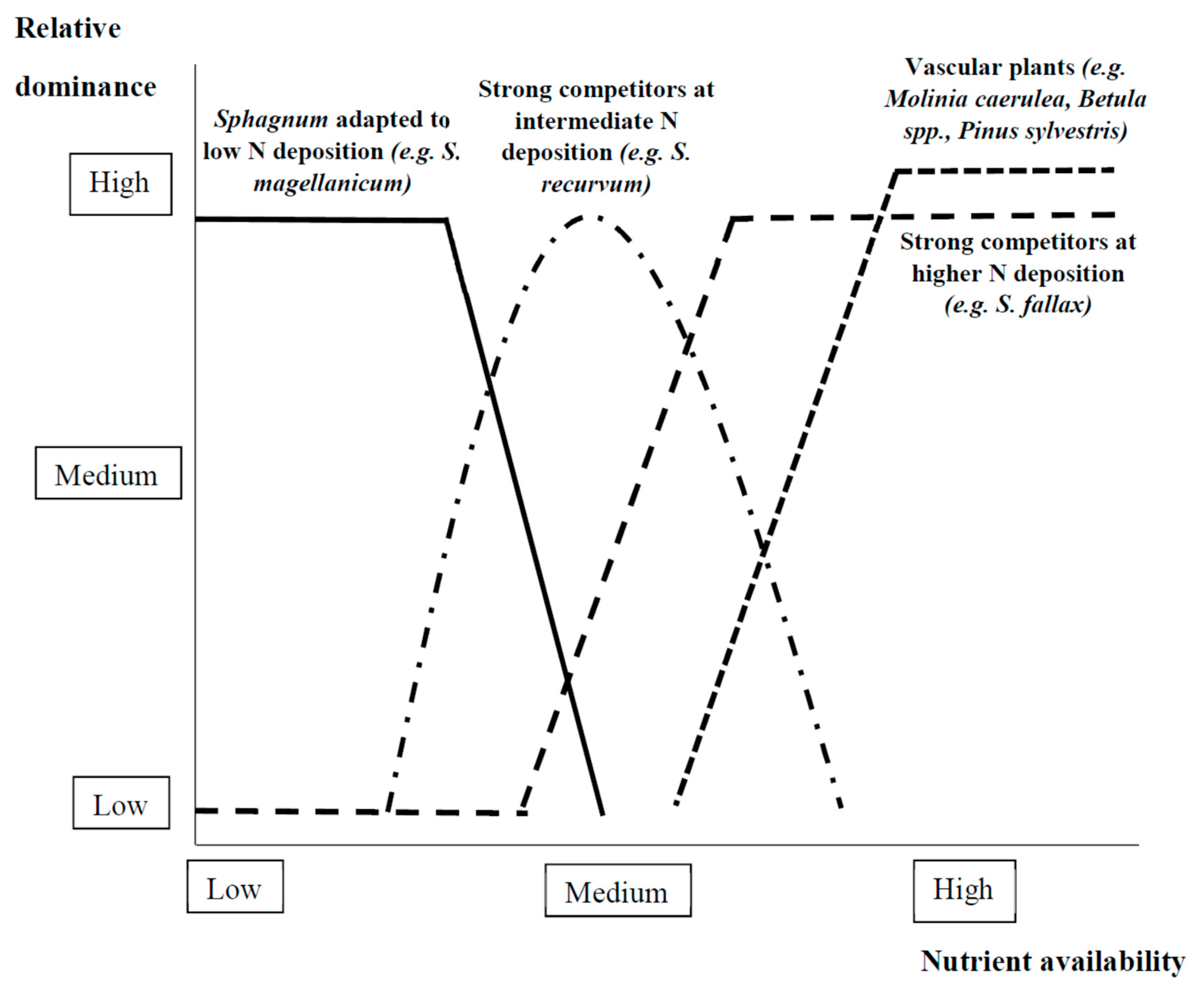
2.2.2. Foundation Species Interactions with Nutrients
Algae
Vascular and Nonvascular Plants
Trees
2.2.3. Foundation Species Interactions with Other Environmental Conditions
Algae
Vascular and Nonvascular Plants
Trees
3. Discussion
3.1. The Impacts of Foundation Species on Wetland Ecosystem Structure and Functions
3.2. Foundation Species as Targets of Conservation
3.3. Future Research Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Angelini, C.; Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. BioScience 2011, 61, 782–789. [Google Scholar] [CrossRef]
- Dayton, P.K. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo Sound, Antarctica. In Proceedings of the Colloquium on Conservation Problems in Antarctica; Parker, B.C., Ed.; Allen Press: Dublin, Ireland, 1972; pp. 81–95. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Faunce, C.H.; Serafy, J.E. Selective use of mangrove shorelines by snappers, grunts and great barracuda. Mar. Ecol. Prog. Ser. 2008, 356, 153–162. [Google Scholar] [CrossRef]
- Gaiser, E.E.; McCormick, P.V.; Hagerthey, S.E.; Gottlieb, A.D. Landscape patterns of periphyton in the Florida Everglades. Crit. Rev. Environ. Sci. Technol. 2011, 41, 92–120. [Google Scholar] [CrossRef]
- Parker, G.G.; Harmon, M.A.; Lefsky, J.Q. Three-dimensional structure of an old-growth Pseudotsuga tsuga canopy and its implications for radiation balance, microclimate, and gas exchange. Ecosystems 2004, 7, 440–453. [Google Scholar] [CrossRef]
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities. BioScience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Bruno, J.F.; Bertness, M.D. Habitat modification and facilitation in benthic marine communities. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., Hay, M.E., Eds.; Sinauer: Sunderland, MA, USA, 2001; pp. 201–218. [Google Scholar]
- Gedan, K.B.; Bertness, M.D. How will warming affect the salt marsh foundation species Spartina patens and its ecological role? Oecologia 2010, 164, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Enwright, N.; Day, R.H.; Doyle, T.W. Winter climate change and coastal wetland foundation species: Salt marshes vs. mangrove forests in the southeastern United States. Glob. Chang. Biol. 2013, 19, 1482–1494. [Google Scholar] [CrossRef]
- Thompson, D.K.; Baisley, A.S.; Waddington, J.M. Seasonal variation in albedo and radiation exchange between a burned and unburned forested peatland: Implications for peatland evaporation. Hydrol. Process. 2015, 29, 3227–3235. [Google Scholar] [CrossRef]
- Sharitz, R.; Mitsch, W.J. Southern floodplain forests. In Biodiversity of the Southeastern United States: Lowland Terrestrial Communities; Echternacht, A.C., Ed.; John Wiley and Sons: New York, NY, USA, 1993. [Google Scholar]
- Kominoski, J.S.; Gaiser, E.E.; Baer, S.G. Advancing Theories of Ecosystem Development through Long-Term Ecological Research. BioScience 2018, 68, 554–562. [Google Scholar] [CrossRef]
- Junk, W.J.; An, S.; Finlayson, C.M.; Gopal, B.; Květ, J.; Mitchell, S.A.; Mitsch, W.J.; Robarts, R.D. Current state of knowledge regarding the world’s wetlands and their future under global climate change: A synthesis. Aquat. Sci. 2013, 75, 151–167. [Google Scholar] [CrossRef]
- Scrosati, R.A. Community-level facilitation by macroalgal foundation species peaks at an intermediate level of environmental stress. Algae 2017, 32, 41–46. [Google Scholar] [CrossRef]
- Smith, T.J.; Foster, A.M.; Tiling-Range, G.; Jones, J.B., Jr. Dynamics of mangrove-marsh ecotones in subtropical coastal wetlands: Fire, sea-level rise, and water levels. Fire Ecol. 2013, 9, 66–77. [Google Scholar] [CrossRef]
- Barr, J.G.; Engel, V.; Smith, T.J.; Fuentes, J.D. Hurricane disturbance and recovery of energy balance, CO2 fluxes and canopy structure in a mangrove forest of the Florida Everglades. Agric. For. Meteorol. 2012, 153, 54–66. [Google Scholar] [CrossRef]
- Davis, S.E.; Boucek, R.; Castañeda-Moya, E.; Dessu, S.; Gaiser, E.; Kominoski, J.; Sah, J.P.; Surratt, D.; Troxler, T. Episodic disturbances drive nutrient dynamics along freshwater-to-estuary gradients in a subtropical wetland. Ecosphere 2018, 9, e02296. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Michalet, R.; Broker, R.W.; Cavieres, L.A.; Kikvidze, Z.; Lortie, C.J.; Pugnaire, F.I.; Valiente-Banuet, A.; Callaway, R.M. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 2006, 9, 767–773. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M. Strong facilitation in mild environments: The stress gradient hypothesis revisited. J. Ecol. 2010, 98, 1269–1275. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altien, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [Green Version]
- Passy, S.I. Abundance inequality in freshwater communities has an ecological origin. Am. Nat. 2016, 187, 502–516. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Newman, S.; Schuette, J.; Grace, J.B.; Rutchey, K.; Fontaine, T.; Reddy, K.R.; Pietrucha, M. Factors influencing cattail abundance in the northern Everglades. Aquat. Bot. 1998, 60, 265–280. [Google Scholar] [CrossRef]
- Crain, C.M.; Bertness, M.D. Community impacts of a tussock sedge: Is ecosystem engineering important in benign habitats? Ecology 2005, 86, 2695–2704. [Google Scholar] [CrossRef]
- Costanza, R.; de Groot, R.; Sutton, P.; Van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 3rd ed.; Wiley: New York, NY, USA, 2000; 920p. [Google Scholar]
- Eppinga, M.B.; Rietkerk, M.; Wassen, M.J.; De Ruiter, P.C. Linking habitat modification to catastrophic shifts and vegetation patterns in bogs. Plant Ecol. 2009, 200, 53–68. [Google Scholar] [CrossRef]
- Larsen, L.G.; Harvey, J.W.; Crimaldi, J.P. A delicate balance: Ecohydrological feedbacks governing landscape morphology in a lotic peatland. Ecol. Monogr. 2007, 77, 591–614. [Google Scholar] [CrossRef]
- Douglas, M.S. The Everglades: River of Grass; Pineapple Press Inc.: Sarasota, FL, USA, 2007. [Google Scholar]
- Lodge, T.E. The Everglades Handbook: Understanding the Ecosystem; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Lee, S.S. Mastogloia calcarea. 2014. Available online: https://diatoms.org/species/mastogloia_calcarea (accessed on 30 October 2018).
- Gaiser, E.E.; La Hée, J.M.L.; Tobias, F.A.; Wachnicka, A.H. Mastogloia smithii var. lacustris Grun.: A structural engineer of calcareous mats in karstic subtropical wetlands. Proc. Acad. Natl. Sci. USA 2010, 160, 99–112. [Google Scholar]
- Gottlieb, A.D.; Richards, J.H.; Gaiser, E.E. Comparative study of periphyton community structure in long and short-hydroperiod Everglades marshes. Hydrobiologia 2006, 569, 195–207. [Google Scholar] [CrossRef]
- La Hée, J.M.; Gaiser, E.E. Benthic diatom assemblages as indicators of water quality in the Everglades and three tropical karstic wetlands. Freshw. Sci. 2012, 31, 205–221. [Google Scholar] [CrossRef]
- Lee, S.S.; Gaiser, E.E.; Trexler, J.C. Diatom-based models for inferring hydrology and periphyton abundance in a subtropical karstic wetland: Implications for ecosystem-scale bioassessment. Wetlands 2013, 33, 157–173. [Google Scholar] [CrossRef]
- Marazzi, L.; Gaiser, E.E.; Tobias, F.A. Phosphorus scarcity and desiccation stress increase the occurrence of dominant taxa in wetland benthic primary producer communities. Aquat. Ecol. 2017, 51, 571–589. [Google Scholar] [CrossRef]
- Van Meter, N.N. Some Quantitative and Qualitative Aspects of Periphyton in the Everglades. Master’s Thesis, University of Miami, Coral Gables, FL, USA, 1965; 98p. [Google Scholar]
- Lee, S.S.; Gaiser, E.E.; Van De Vijver, B.; Edlund, M.B.; Spaulding, S.A. Morphology and typification of Mastogloia smithii and M. lacustris, with descriptions of two new species from the Florida Everglades and the Caribbean region. Diatom Res. 2014, 29, 325–350. [Google Scholar] [CrossRef]
- Hagerthey, S.E.; Bellinger, B.J.; Wheeler, K.; Gantar, M.; Gaiser, E.E. Everglades periphyton: A biogeochemical perspective. Crit. Rev. Environ. Sci. Technol. 2011, 41 (Suppl. 1), 309–343. [Google Scholar] [CrossRef]
- Trexler, J.C.; Loftus, W.F. Invertebrates of the Florida Everglades. In Invertebrates in Freshwater Wetlands: An International Perspective on their Ecology; Batzer, D., Boix, D., Eds.; Springer: New York, NY, USA, 2016; pp. 321–356. [Google Scholar]
- Gaiser, E.E.; Gottlieb, A.D.; Lee, S.S.; Trexler, J.C. The Importance of Species-Based Microbial Assessment of Water Quality in Freshwater Everglades Wetlands. In Microbiology of the Everglades Ecosystem; Entry, J., Jayachandran, K., Gottlieb, A.D., Ogram, A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 115–130. [Google Scholar]
- Gottlieb, A.; Richards, J.; Gaiser, E. Effects of desiccation duration on the community structure and nutrient retention of short and long-hydroperiod Everglades periphyton mats. Aquat. Bot. 2005, 82, 99–112. [Google Scholar] [CrossRef]
- Rejmánková, E.; Komárková, J. A function of cyanobacterial mats in phosphorus-limited tropical wetlands. Hydrobiologia 2000, 431, 135–153. [Google Scholar] [CrossRef]
- Novelo, E.; Tavera, R. The role of periphyton in the regulation and supply of nutrients in a wetland at El Edén, Quintana Roo. In The Lowland Maya Area: Three Millennia at the Human-Wildland Interface; Food Products Press: New York, NY, USA, 2003; pp. 217–236. [Google Scholar]
- Loveless, C.M. A study of the vegetation in the Florida Everglades. Ecology 1959, 40, 1–9. [Google Scholar] [CrossRef]
- Miao, S.L.; Sklar, F.H. Biomass and nutrient allocation of sawgrass and cattail along a nutrient gradient in the Florida Everglades. Wetl. Ecol. Manag. 1998, 5, 245–263. [Google Scholar] [CrossRef]
- Herndon, A.; Gunderson, L.; Stenberg, J. Sawgrass (Cladium jamaicense) survival in a regime of fire and flooding. Wetlands 1991, 11, 17–28. [Google Scholar] [CrossRef]
- Givnish, T.J.; Volin, J.C.; Owen, V.D.; Volin, V.C.; Muss, J.D.; Glaser, P.H. Vegetation differentiation in the patterned landscape of the central Everglades: Importance of local and landscape drivers. Glob. Ecol. Biogeogr. 2008, 17, 384–402. [Google Scholar] [CrossRef]
- Ross, M.S.; Reed, D.L.; Sah, J.P.; Ruiz, P.L.; Lewin, M.T. Vegetation:environment relationships and water management in Shark Slough, Everglades National Park. Wetl. Ecol. Manag. 2003, 11, 291–303. [Google Scholar] [CrossRef]
- Ross, M.S.; Sah, J.P.; Ruiz, P.L.; Jones, D.T.; Cooley, H.C.; Travieso, R.; Snyder, J.R.; Hagyari, D. Effect of Hydrology Restoration on the Habitat of the Cape Sable Seaside Sparrow; Report to Everglades National Park, SERC Research Reports. 85; South Florida Natural Resource Center: Homestead, FL, USA, 2006; pp. 1–46.
- Todd, M.J.; Muneepeerakul, R.; Pumo, D.; Azaele, S.; Miralles-Wilhelm, F.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydrological drivers of wetland vegetation community distribution within Everglades National Park, Florida. Adv. Water Resour. 2010, 33, 1279–1289. [Google Scholar] [CrossRef]
- Sah, J.P.; Ross, M.S.; Saha, S.; Minchin, P.; Sadle, J. Trajectories of vegetation response to water management in Taylor Slough, Everglades National Park, Florida. Wetlands 2014, 34, 65–79. [Google Scholar] [CrossRef]
- Toth, L.A. Effects of Hydrologic Regimes on Lifetime Production and Nutrient Dynamics of Sawgrass (Technical Publication# 87-6); Environmental Sciences Division: West Palm Beach, FL, USA, 1987. [Google Scholar]
- Busch, D.E.; Loftus, W.F.; Bass, O.L. Long-term hydrologic effects on marsh plant community structure in the southern Everglades. Wetlands 1998, 18, 230–241. [Google Scholar] [CrossRef]
- Armentano, T.V.; Sah, J.P.; Ross, M.S.; Jones, D.T.; Cooley, H.C.; Smith, C.S. Rapid responses of vegetation to hydrological changes in Taylor Slough, Everglades National Park, Florida, USA. Hydrobiologia 2006, 569, 293–309. [Google Scholar] [CrossRef]
- Gunderson, L.H. South florida: The reality of change and the prospects for sustainability: Managing surprising ecosystems in southern Florida. Ecol. Econ. 2001, 37, 371–378. [Google Scholar] [CrossRef]
- Mazzotti, F.J.; Fling, H.E.; Merediz, G.; Lazcano, M.; Lasch, C.; Barnes, T. Conceptual ecological model of the Sian Ka’an Biosphere Reserve, Quintana Roo, Mexico. Wetlands 2005, 25, 980–997. [Google Scholar] [CrossRef]
- Imbert, D.; Delbé, L. Ecology of fire-influenced Cladium jamaicense marshes in Guadeloupe, Lesser Antilles. Wetlands 2006, 26, 289–297. [Google Scholar] [CrossRef]
- Meerman, J.C. Ecological Characterization of the New River Lagoon, Orange Walk District, Belize; Report prepared for the Freshwater Program; Programme for Belize: Belize City, BZ, USA, 2006; pp. 2–26. [Google Scholar]
- Rejmánková, E.; Pope, K.O.; Post, R.; Maltby, E. Herbaceous wetlands of the Yucatan peninsula: Communities at extreme ends of environmental gradients. Int. Rev. Gesamten Hydrobiol. 1996, 81, 223–252. [Google Scholar] [CrossRef]
- Grace, J.B. The effects of nutrient additions on mixtures of Typha latifolia L. and Typha domingensis Pers. along a water depth gradient. Aquat. Bot. 1988, 31, 83–92. [Google Scholar] [CrossRef]
- Sun, G.; McNulty, S.G.; Shepard, J.P.; Amatya, D.M.; Riekerk, H.; Comerford, N.B.; Skaggs, W.; Swift, L., Jr. Effects of timber management on the hydrology of wetland forests in the southern United States. For. Ecol. Manag. 2001, 143, 227–236. [Google Scholar] [CrossRef]
- Feliciano, E.A. Multi-Scale Remote Sensing Assessments of Forested Wetlands: Applications to the Everglades National Park. Open Access Diss. 2015, p. 1417. Available online: https://scholarlyrepository.miami.edu/oa_dissertations/1417 (accessed on 1 September 2018).
- Allen, J.A.; Pezeshki, S.R.; Chambers, J.L. Interaction of flooding and salinity stress on baldcypress (Taxodium distichum). Tree Physiol. 1996, 16, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Megonigal, J.P.; Day, F.P. Effects of flooding on root and shoot production of bald cypress in large experimental enclosures. Ecology 1992, 73, 1182–1193. [Google Scholar] [CrossRef]
- Shanklin, J.; Kozlowski, T.T. Effect of flooding of soil on growth and subsequent responses of Taxodium distichum seedlings to SO2. Environ. Pollut. Ser. A Ecol. Biol. 1985, 38, 199–212. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Delaune, R.D.; Patrick, W.H., Jr. Gas exchange characteristics of bald cypress (Taxodium distichum L.): Evaluation of responses to leaf aging, flooding, and salinity. Can. J. For. Res. 1986, 16, 1394–1397. [Google Scholar] [CrossRef]
- Pezeshki, S.R. Root responses of flood-tolerant and flood-sensitive tree species to soil redox conditions. Trees 1991, 5, 180–186. [Google Scholar] [CrossRef]
- Pezeshki, S. Response of baldcypress (Taxodium distichum) seedlings to hypoxia: Leaf protein content, ribulose-1,5-bisphosphate carboxy-lase/oxygenase activity and photosynthesis. Photosynthetica 1994, 30, 59–68. [Google Scholar]
- Crawford, R.M.M. Root survival in flooded soils. In Ecosystems of the World; Gore, A.J.P., Ed.; Elsevier Scientific: Amsterdam, The Netherlands, 1983; pp. 257–283. [Google Scholar]
- Hook, D.D. Adaptations to flooding with fresh water. In Flooding and Plant Growth; Kozlowski, T.T., Ed.; Academic Press: Orlando, FL, USA, 1984; pp. 265–294. [Google Scholar]
- Wang, M.; Talbot, J.; Moore, T.R. Drainage and fertilization effects on nutrient availability in an ombrotrophic peatland. Sci. Total Environ. 2017, 121, 1255–1263. [Google Scholar] [CrossRef]
- Elcan, J.M.; Pezeshki, S.R. Effects of Flooding on Susceptibility of Taxodium distichum L. Seedlings to Drought. Photosynthetica 2002, 40, 177–182. [Google Scholar] [CrossRef]
- Lei, T.; Middleton, B.A. Repeated Drought Alters Resistance of Seed Bank Regeneration in Baldcypress Swamps of North America. Ecosystems 2018, 21, 190–201. [Google Scholar] [CrossRef]
- Gaiser, E.E.; Richards, J.H.; Trexler, J.C.; Jones, R.D.; Childers, D.L. Periphyton responses to eutrophication in the Florida Everglades: Cross-system patterns of structural and compositional change. Limnol. Oceanogr. 2006, 51, 617–630. [Google Scholar] [CrossRef] [Green Version]
- Gaiser, E.E. Periphyton as an indicator of restoration in the Florida Everglades. Ecol. Indic. 2009, 9, S37–S45. [Google Scholar] [CrossRef]
- Pinowska, A.; Francoeur, S.N.; Manoylov, K.M. Substratum-associated microbiota. Water Environ. Res. Lit. Rev. 2008, 80, 1858–1891. [Google Scholar] [CrossRef]
- Dodds, W.K. The role of periphyton in phosphorus retention in shallow freshwater aquatic systems. J. Phycol. 2003, 39, 840–849. [Google Scholar] [CrossRef]
- Lamberti, G.A. The role of periphyton in benthic food webs. In Algal Ecology, 1st ed.; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 533–564. [Google Scholar]
- Hagerthey, S.E.; Cole, J.J.; Kilbane, D. Aquatic metabolism in the Everglades: Dominance of water column heterotrophy. Limnol. Oceanogr. 2010, 55, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Rejmankova, E.; Pope, K.O.; Pohl, M.D.; Rey-Benayas, J.M. Freshwater wetland plant communities of northern Belize: Implications for paleoecological studies of Maya wetland agriculture. Biotropica 1995, 27, 28–36. [Google Scholar] [CrossRef]
- Gaiser, E.E.; Anderson, E.P.; Castañeda-Moya, E.; Collado-Vides, L.; Fourqurean, J.W.; Heithaus, M.R.; Jaffé, R.; Lagomasino, D.; Oehm, N.J.; Price, R.M.; et al. New perspectives on an iconic landscape from comparative international long-term ecological research. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef]
- Steward, K.K.; Ornes, W.H. The autecology of sawgrass in the Florida Everglades. Ecology 1975, 56, 162–171. [Google Scholar] [CrossRef]
- Steward, K.K.; Ornes, W.H. Mineral nutrition of sawgrass (Cladium jamaicense Crantz) in relation to nutrient supply. Aquat. Bot. 1983, 16, 349–359. [Google Scholar] [CrossRef]
- Davis, S.M. Sawgrass and cattail nutrient flux: Leaf turnover, decomposition, and nutrient flux of sawgrass and cattail in the Everglades. Aquat. Bot. 1991, 40, 203–224. [Google Scholar] [CrossRef]
- Noe, G.B.; Childers, D.L.; Jones, R.D. Phosphorus Biogeochemistry and the Impact of Phosphorus Enrichment: Why Is the Everglades so Unique? Ecosystems 2001, 4, 603–624. [Google Scholar] [CrossRef]
- Richardson, C.J.; Ferrell, G.M.; Vaithiyanathan, P. Nutrient effects on stand structure, resorption efficiency, and secondary compounds in Everglades sawgrass. Ecology 1999, 80, 2182–2192. [Google Scholar] [CrossRef]
- Brix, H.; Lorenzen, B.; Mendelssohn, I.A.; McKee, K.L.; Miao, S.L. Can differences in phosphorus uptake kinetics explain the distribution of cattail and sawgrass in the Florida Everglades? Plant Biol. 2010, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beare, P.A.; Zedler, J.B. Cattail invasion and persistence in a coastal salt marsh: The role of salinity. Estuaries 1987, 10, 165–170. [Google Scholar] [CrossRef]
- Thomas, S.E.; Gaiser, E.E.; Tobias, F.A.C. Effects of shading on calcareous benthic periphyton in a short-hydroperiod oligotrophic alkaline wetland (Everglades, FL, USA). Hydrobiologia 2006, 569, 209–221. [Google Scholar] [CrossRef]
- Grimshaw, H.J.; Rosen, M.; Swift, D.R.; Rodberg, K.; Noel, J.M. Marsh phosphorus concentrations, phosphorus content and species composition of Everglades periphyton communities. Arch. Hydrobiol. 1993, 139, 17–27. [Google Scholar]
- Chapin, F.S., III. The mineral nutrition of wild plants. Ann. Rev. Ecol. System. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Newman, S.; Grace, J.B.; Koebel, J.W. Effects of nutrients and hydroperiod on Typha, Cladium, and Eleocharis: Implications for Everglades restoration. Ecol. Appl. 1996, 6, 774–783. [Google Scholar] [CrossRef]
- Davis, S.M.; Ogden, J.C. Towards ecosystem restoration. In Everglades, The Ecosystem and Its Restoration; Davis, S.M., Ogden, J.C., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1994; pp. 769–796. [Google Scholar]
- Hagerthey, S.E.; Newman, S.; Rutchey, K.; Smith, E.P.; Godin, J. Multiple regime shifts in a subtropical peatland: Community-specific thresholds to eutrophication. Ecol. Monogr. 2008, 78, 547–565. [Google Scholar] [CrossRef]
- Effler, R.S.; Goyer, R.A.; Lenhard, G.J. Baldcypress and water tupelo responses to insect defoliation and nutrient augmentation in Maurepas Swamp, Louisiana, USA. For. Ecol. Manag. 2006, 236, 295–304. [Google Scholar] [CrossRef]
- Krauss, K.W.; Duberstein, J.A.; Doyle, T.W.; Conner, W.H.; Day, R.H.; Inabinette, L.W.; Whitbeck, J.L. Site condition, structure, and growth of baldcypress along tidal/non-tidal salinity gradients. Wetlands 2009, 29, 505–519. [Google Scholar] [CrossRef]
- Nyman, J.A.; Lindau, C.W. Nutrient availability and flooding stress interact to affect growth and mercury concentration in Taxodium distichum (L.) Rich. seedlings. Environ. Exp. Bot. 2016, 125, 77–86. [Google Scholar] [CrossRef]
- Dickson, R.E.; Broyer, T.C. Effects of aeration, water supply, and nitrogen source on growth and development of tupelo gum and bald cypress. Ecology 1972, 53, 626–634. [Google Scholar] [CrossRef]
- Dickson, R.E.; Broyer, T.C.; Johnson, C.M. Nutrient uptake by tupelo gum and bald cypress from saturated or unsaturated soil. Plant Soil 1972, 37, 297–308. [Google Scholar] [CrossRef]
- Effler, R.S.; Goyer, R.A. Baldcypress and water tupelo sapling response to multiple stress agents and reforestation implications for Louisiana swamps. For. Ecol. Manag. 2006, 226, 330–340. [Google Scholar] [CrossRef]
- Day, R.H.; Doyle, T.W.; Draugelis-Dale, R.O. Interactive effects of substrate, hydroperiod, and nutrients on seedling growth of Salix nigra and Taxodium distichum. Environ. Exp. Bot. 2006, 55, 163–174. [Google Scholar] [CrossRef]
- Hesse, I.D.; Day, J.W.; Doyle, T.W. Long-Term Growth Enhancement of Baldcypress (Taxodium distichum) from Municipal Wastewater Application. Environ. Manag. 1998, 22, 119–127. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Westphal, A.; Pratt, R.; Hyfield, E.; Rybczyk, J.; Kemp, G.P.; Day, J.N.; Marx, B. Effects of long-term municipal effluent discharge on the nutrient dynamics, productivity, and benthic community structure of a tidal freshwater forested wetland in Louisiana. Ecol. Eng. 2006, 27, 242–257. [Google Scholar] [CrossRef]
- Keim, R.F.; Izdepski, C.W.; Day, J.W. Growth Responses of Baldcypress to Wastewater Nutrient Additions and Changing Hydrologic Regime. Wetlands 2012, 32, 95–103. [Google Scholar] [CrossRef]
- Mazzei, V.; Gaiser, E. Scale and spatial consistency of specialization in an endemic and abundant freshwater diatom from the Caribbean Basin. Freshw. Sci. 2017, 36, 542–554. [Google Scholar] [CrossRef]
- Mazzei, V.; Gaiser, E.E.; Kominoski, J.S.; Troxler, T.; Wilson, B.; Servais, S.; Bauman, L.; Davis, S.; Kelly, S.; Sklar, F.; et al. Functional and compositional responses of periphyton mats to simulated saltwater intrusion in the southern Everglades. Estuaries Coasts 2018, 41, 2105–2119. [Google Scholar] [CrossRef]
- Obeysekera, J.; Barnes, J.; Nungesser, M. Climate sensitivity runs and regional hydrologic modeling for predicting the response of the greater Florida Everglades ecosystem to climate change. Environ. Manag. 2015, 55, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Sah, J.P.; Ross, M.S.; Snyder, J.R.; Stoffella, S.; Blanco, J.; Freixa, J. Re-Sampling of Vegetation Survey Sites within Cape Sable Seaside Sparrow Habitat; South Florida Natural Resources Center, Everglades & Dry Tortugas National Parks: Homestead, FL, USA, 2015; p. 53.
- Alexander, T.R. Sawgrass biology related to the future of the Everglades ecosystem. Proc. Soil Crop Sci. Soc. Fla. 1971, 31, 72–74. [Google Scholar]
- Doyle, T.W.; Krauss, K.W.; Conner, W.H. Predicting the retreat and migration of tidal forests along the northern Gulf of Mexico under sea-level rise. For. Ecol. Manag. 2010, 259, 770–777. [Google Scholar] [CrossRef]
- Ross, M.S.; Meeder, J.F.; Sah, J.P.; Ruiz, P.L.; Telesnicki, G.J. The southeast saline Everglades revisited: 50 years of coastal vegetation change. J. Veg. Sci. 2000, 11, 101–112. [Google Scholar] [CrossRef]
- Furtado, J.I.; Verghese, S.; Liew, K.S.; Lee, T.H. Litter production in a freshwater swamp forest Tasek Bera, Malaysia. In Tropical Ecology and Development, Proceedings of the 5th International Symposium of Tropical Ecology, Kuala Lumpur, Malaysia, 16–21 April 1979; Furtado, J.I., Ed.; International Society of Tropical Ecology: Kuala Lumpur, Malaysia; pp. 815–822.
- Alvarez-López, M. Ecology of Pterocarpus officinalis forested wetlands in Puerto Rico. In Forested Wetlands; Elsevier: Amsterdam, The Netherlands, 1990; pp. 251–265. [Google Scholar]
- Frangi, J.L.; Lugo, A.E. Ecosystem dynamics of a subtropical floodplain forest. Ecol. Monogr. 1985, 55, 351–369. [Google Scholar] [CrossRef]
- Lugo, A.E.; Brown, S.; Brinson, M.M. Forested wetlands in freshwater and salt-water environments. Limnol. Oceanogr. 1988, 33, 894–909. [Google Scholar] [Green Version]
- Liu, X.; Conner, W.H.; Song, B.; Jayakaran, A.D. Forest composition and growth in a freshwater forested wetland community across a salinity gradient in South Carolina, USA. For. Ecol. Manag. 2017, 389, 211–219. [Google Scholar] [CrossRef]
- Pierfelice, K.N.; Lockaby, B.G.; Krauss, K.W.; Conner, W.H.; Noe, G.B.; Ricker, M.C. Salinity Influences on Aboveground and Belowground Net Primary Productivity in Tidal Wetlands. J. Hydrol. Eng. 2017, 22, D5015002. [Google Scholar] [CrossRef]
- Thomas, B.L.; Doyle, T.; Krauss, K. Annual Growth Patterns of Baldcypress (Taxodium distichum) Along Salinity Gradients. Wetlands 2015, 35, 831–839. [Google Scholar] [CrossRef]
- Krauss, K.W.; Chambers, J.L.; Creech, D. Selection for salt tolerance in tidal freshwater swamp species: Advances using baldcypress as a model for restoration. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Conner, W.H., Doyle, T.W., Krauss, K.W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 385–410. [Google Scholar]
- Allen, J.A.; Chambers, J.L.; Pezeshki, S.R. Effects of salinity on baldcypress seedlings: Physiological responses and their relation to salinity tolerance. Wetlands 1997, 17, 310–320. [Google Scholar] [CrossRef]
- Duever, M.J.; Meeder, J.F.; Duever, L.C. Ecosystems of the Big Cypress Swamp. In Cypress Swamps; Ewel, K.C., Odum, H.T., Eds.; University Presses of Florida: Gainesville, FL, USA, 1984; pp. 294–303. [Google Scholar]
- Krauss, K.W.; Duberstein, J.A. Sapflow and water use of freshwater wetland trees exposed to saltwater incursion in a tidally influenced South Carolina watershed. Can. J. For. Res. 2010, 40, 525–535. [Google Scholar] [CrossRef]
- Fry, B.; Bern, A.L.; Ross, M.S.; Meeder, J.F. δ15N Studies of Nitrogen Use by the Red Mangrove, Rhizophora mangle L. in South Florida. Estuar. Coast. Shelf Sci. 2000, 50, 291–296. [Google Scholar] [CrossRef]
- McKee, K.L.; Feller, I.C.; Popp, M.; Wanek, W. Mangrove isotopic (δ15N and δ13C) fractionation across a nitrogen vs. phosphorus limitation gradient. Ecology 2002, 83, 1065–1075. [Google Scholar] [CrossRef]
- McWilliams, W.H.; Rosson, J.F., Jr. Composition and vulnerability of bottomland hardwood forests of the coastal plain province in the south central United States. For. Ecol. Manag. 1990, 33/34, 485–501. [Google Scholar] [CrossRef]
- Zhai, L.; Krauss, K.W.; Liu, X.; Duberstein, J.A.; Conner, W.H.; DeAngelis, D.L.; Sternberg, L.d.S.L. Growth stress response to sea level rise in species with contrasting functional traits: A case study in tidal freshwater forested wetlands. Environ. Exp. Bot. 2018, 155, 378–386. [Google Scholar] [CrossRef]
- Krauss, K.W.; Duberstein, J.A.; Conner, W.H. Assessing stand water use in four coastal wetland forests using sapflow techniques: Annual estimates, errors and associated uncertainties. Hydrol. Process. 2015, 29, 112–127. [Google Scholar] [CrossRef]
- Kemp, P.G.; Conner, W.H.; Day, J.W., Jr. Effects of flooding on decomposition and nutrient cycling in a Louisiana swamp forest. Wetlands 1985, 5, 35–50. [Google Scholar] [CrossRef]
- Schumann, M.; Joosten, H. Global Peatland Restoration Manual. Institute of Botany and Landscape Ecology; Greifswald University: Greifswald, Germany, 2008; p. 103. [Google Scholar]
- Potapova, M. Navicymbula pusilla. 2011. Available online: https://diatoms.org/species/navicymbula_pusilla (accessed on 30 October 2018).
- Cooper, M.J.; Costello, G.M.; Francoeur, S.N.; Lamberti, G.A. Nitrogen limitation of algal biofilms in coastal wetlands of Lakes Michigan and Huron. Freshw. Sci. 2016, 35, 25–40. [Google Scholar] [CrossRef]
- Timoney, K.; Zoltai, S.C.; Goldsborough, L.G. Boreal diatom ponds: A rare wetland associated with nesting whooping cranes. Wetlands 1997, 17, 539–551. [Google Scholar] [CrossRef]
- Johnson, R.E.; Tuchman, N.C.; Peterson, C.G. Changes in the vertical microdistribution of diatoms within a developing periphyton mat. J. N. Am. Benthol. Soc. 1997, 16, 503–519. [Google Scholar] [CrossRef]
- Kilroy, C.; Larned, S.T.; Biggs, B.J.F. The non-indigenous diatom Didymosphenia geminata alters benthic communities in New Zealand rivers. Freshw. Biol. 2009, 54, 1990–2002. [Google Scholar] [CrossRef]
- Belyea, L.R.; Clymo, R.S. Feedback control of the rate of peat formation. Proc. R. Soc. B 2001, 268, 1315–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkman, J.J. Plant communities and synecology of bogs and heath pools in the Netherlands. In Fens and Bogs in the Netherlands: Vegetation, History, Nutrient Dynamics and Conservation; Verhoeven, J.T.A., Ed.; Springer: Dordrecht, The Netherlands, 1992; pp. 173–236. [Google Scholar]
- Robroek, B.J.M.; Limpens, J.; Breeuwer, A.; van Ruijven, J.; Schouten, M.G.C. Precipitation determines the persistence of hollow Sphagnum species on hummocks. Wetlands 2007, 27, 976–986. [Google Scholar] [CrossRef]
- Hájek, T.; Beckett, R.P. Effect of water content components on desiccation and recovery in Sphagnum mosses. Ann. Bot. 2007, 101, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Vitt, D.H. Functional Characteristics and Indicators of Boreal Peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Ecological Studies (Analysis and Synthesis); Springer: Berlin/Heidelberg, Germany, 2006; Volume 188. [Google Scholar]
- Bengtsson, F.; Granath, G.; Rydin, H. Photosynthesis, growth, and decay traits in Sphagnum—A multispecies comparison. Ecol. Evol. 2016, 6, 3325–3341. [Google Scholar] [CrossRef]
- Van Breemen, N. How Sphagnum bogs down other plants. Trends Ecol. Evol. 1995, 10, 270–275. [Google Scholar] [CrossRef]
- Więcek, M.; Martin, P.; Gąbka, M. Distribution patterns and environmental correlates of water mites (Hydrachnidia, Acari) in peatland microhabitats. Exp. Appl. Acarol. 2013, 61, 147–160. [Google Scholar] [CrossRef] [Green Version]
- Eppinga, M.B.; Rietkerk, M.; Borren, W.; Lapshina, E.D.; Bleuten, W.; Wassen, M.J. Regular surface patterning of peatlands: Confronting theory with field data. Ecosystems 2008, 11, 520–536. [Google Scholar] [CrossRef]
- Malmer, N.; Albinsson, C.; Svensson, B.M.; Wallén, B. Interferences between Sphagnum and vascular plants: Effects on plant community structure and peat formation. Oikos 2003, 100, 469–482. [Google Scholar] [CrossRef]
- Frankl, R.; Schmeidl, H. Vegetation change in a South German raised bog: Ecosystem engineering by plant species, vegetation switch or ecosystem level feedback mechanisms? Flora 2000, 195, 267–276. [Google Scholar] [CrossRef]
- Hirano, T.; Suzuki, K.; Hirata, R. Energy balance and evapotranspiration changes in a larch forest caused by severe disturbance during an early secondary succession. Agric. For. Meteorol. 2017, 232, 457–468. [Google Scholar] [CrossRef]
- Takagi, K.; Tsuboya, T.; Takahashi, H.; Inoue, T. Effect of the invasion of vascular plants on heat and water balance in the Sarobetsu mire, northern Japan. Wetlands 1999, 19, 246–254. [Google Scholar] [CrossRef]
- Lamers, L.P.M.; Bobbink, R.; Roelofs, J.G.M. Natural nitrogen filter fails in polluted raised bogs. Glob. Chang. Biol. 2007, 6, 583–586. [Google Scholar] [CrossRef]
- Fay, E.; Lavoie, C. The impact of birch seedlings on evapotranspiration from a mined peatland: An experimental study in southern Quebec, Canada. Mires Peat 2009, 5, 3. [Google Scholar]
- Ohlson, M.; Økland, R.H.; Nordbakken, J.-F.; Dahlberg, B. Fatal interactions between Scots pine and Sphagnum mosses in bog ecosystems. Oikos 2001, 94, 425–432. [Google Scholar] [CrossRef]
- Gunnarsson, U.; Rydin, H. Demography and recruitment of Scots pine on raised bogs in eastern Sweden and relationships to microhabitat differentiation. Wetlands 1998, 18, 133–141. [Google Scholar] [CrossRef]
- Iremonger, S.F.; Kelly, D.L. The responses of four Irish wetland tree species to raised soil water levels. New Phytol. 1988, 109, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Rutter, A.J. Studies in the water relations of Pinus sylvestris in plantation conditions. I. Measurements of rainfall and interception. J. Ecol. 1963, 51, 191–203. [Google Scholar] [CrossRef]
- Limpens, J.; Berendse, F.; Klees, H. How phosphorus availability affects the impact of nitrogen deposition on Sphagnum and vascular plants in bogs. Ecosystems 2004, 7, 793–804. [Google Scholar] [CrossRef]
- Kokfelt, U.; Struyf, E.; Randsalu, L. Diatoms in peat–Dominant producers in a changing environment? Soil Biol. Biochem. 2009, 41, 1764–1766. [Google Scholar] [CrossRef]
- Rober, A.R.; Stevenson, R.J.; Wyatt, K.H. The role of light availability and herbivory on algal responses to nutrient enrichment in a riparian wetland, Alaska. J. Phycol. 2015, 51, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, M.B.; Rietkerk, M.; Belyea, L.R.; Nilsson, M.B.; Ruiter, P.C.D.; Wassen, M.J. Resource contrast in patterned peatlands increases along a climatic gradient. Ecology 2010, 91, 2344–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, B.S. Temporal and spatial patterns of extreme low flows and effects on stream ecosystems in Otago, New Zealand. J. Hydrol. 2002, 257, 115–133. [Google Scholar] [CrossRef]
- Mosley, L.M. Drought impacts on the water quality of freshwater systems; review and integration. Earth-Sci. Rev. 2015, 140, 203–214. [Google Scholar] [CrossRef]
- Pastor, J.; Peckham, B.; Bridgham, S.; Weltzin, J.; Chen, J. Plant community dynamics, nutrient cycling, and alternative stable equilibria in peatlands. Am. Nat. 2002, 160, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, M.M.; Klees, H.; Berendse, F. Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised CO2 and increased N deposition. Oikos 2002, 97, 415–425. [Google Scholar] [CrossRef]
- Limpens, J.; Bohlin, E.; Nilsson, M.B. Phylogenetic or environmental control on the elemental and organo-chemical composition of Sphagnum mosses? Plant Soil 2017, 417, 69–85. [Google Scholar] [CrossRef]
- Twenhöven, F.L. Competition between two Sphagnum species under different deposition levels. J. Bryol. 1992, 17, 71–80. [Google Scholar] [CrossRef]
- Bobbink, R.; Heil, G.W.; Raessen, M.B.A.G. Atmospheric deposition and canopy exchange processes in heathland ecosystems. Environ. Pollut. 1992, 75, 29–37. [Google Scholar] [CrossRef]
- Heil, G.W.; Werger, M.J.A.; De Mol, W.; Van Dam, D.; Heijne, B. Capture of atmospheric ammonium by grassland canopies. Science 1988, 239, 764–765. [Google Scholar] [CrossRef] [PubMed]
- Malmer, N.; Svensson, B.M.; Wallén, B. Interactions between Sphagnum mosses and field layer vascular plants in the development of peat-forming systems. Folia Geobot. Phytotaxon. 1994, 29, 483–496. [Google Scholar] [CrossRef]
- Tomassen, H.B.M.; Smolders, A.J.P.; Lamers, L.P.M.; Roelofs, J.G.M. Stimulated growth of Betula pubescens and Molinia caerulea on ombrotrophic bogs: Role of high levels of atmospheric nitrogen deposition. J. Ecol. 2003, 91, 357–370. [Google Scholar] [CrossRef]
- Eppinga, M.B.; De Ruiter, P.C.; Wassen, M.J.; Rietkerk, M. Nutrients and hydrology indicate the driving mechanisms of peatland surface patterning. Am. Nat. 2009, 173, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 1995. [Google Scholar]
- Rietkerk, M.; Dekker, S.C.; De Ruiter, P.C.; van de Koppel, J. Self-organized patchiness and catastrophic shifts in ecosystems. Science 2004, 305, 1926–1929. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, P.R.; Van Der Valk, A.G.; Newman, S.; Gawlik, D.E.; Troxler Gann, T.; Coronado-Molina, C.A.; Childers, D.L.; Sklar, F.H. Maintaining tree islands in the Florida Everglades: Nutrient redistribution is the key. Front. Ecol. Environ. 2005, 3, 370–376. [Google Scholar] [CrossRef]
- Agnew, A.D.; Collins, S.L.; van der Maarel, E. Mechanisms and processes in vegetation dynamics: Introduction. J. Veg. Sci. 1993, 4, 146–148. [Google Scholar] [CrossRef]
- Hörnberg, G.; Ohlson, M.; Zackrisson, O. Influence of bryophytes and microrelief conditions on Picea abies seed regeneration patterns in boreal old-growth swamp forests. Can. J. For. Res. 1997, 27, 1015–1023. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Abbott, B.W.; Bowden, W.B.; Brovkin, V.; Camill, P.; Canadell, J.G.; Chanton, J.P.; Chapin, F.S.; Christensen, T.R.; Ciais, P.; et al. Expert assessment of vulnerability of permafrost carbon to climate change. Clim. Chang. 2013, 119, 359–374. [Google Scholar] [CrossRef] [Green Version]
- DeColibus, D.T.; Rober, A.R.; Sampson, A.M.; Shurzinske, A.C.; Walls, J.T.; Turetsky, M.R.; Wyatt, K.H. Legacy effects of drought alters the aquatic food web of a northern boreal peatland. Freshw. Biol. 2017, 62, 1377–1388. [Google Scholar] [CrossRef]
- Gentili, F.; Nilsson, M.C.; Zackrisson, O.; DeLuca, T.H.; Sellstedt, A. Physiological and molecular diversity of feather moss associative N2-fixing cyanobacteria. J. Exp. Bot. 2005, 56, 3121–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Eur. J. Phycol. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Angelini, C.; Silliman, B.R. Secondary foundation species as drivers of trophic and functional diversity: Evidence from a tree–epiphyte system. Ecology 2014, 95, 85–196. [Google Scholar] [CrossRef]
- Angelini, C.; van der Heide, T.; Griffin, J.N.; Morton, J.P.; Derksen-Hooijberg, M.; Lamers, L.P.; Smolders, A.J.; Silliman, B.R. Foundation species’ overlap enhances biodiversity and multifunctionality from the patch to landscape scale in southeastern United States salt marshes. Proc. R. Soc. B 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Siegel, D.I.; Zhu, C.; Chanton, J.P.; Glaser, P.H. Geochemical mixing in peatland waters: The role of organic acids. Wetlands 2015, 35, 567–575. [Google Scholar] [CrossRef]
- Siegel, D.I.; Glaser, P.H.; So, J.; Janecky, D.R. The dynamic balance between organic acids and circumneutral groundwater in a large boreal peat basin. J. Hydrol. 2006, 320, 421–431. [Google Scholar] [CrossRef]
- Glaser, P.H.; Hansen, B.; Siegel, D.I.; Reeve, A.S.; Morin, P.J. Rates, pathways and drivers for peatland development in the Hudson Bay Lowlands, northern Ontario, Canada. J. Ecol. 2004, 92, 1036–1053. [Google Scholar] [CrossRef] [Green Version]
- Bragazza, L.; Alber, R.; Gerdol, R. Seasonal chemistry of pore water in hummocks and hollows in a poor mire in the southern Alps (Italy). Wetlands 1998, 18, 320–328. [Google Scholar] [CrossRef]
- Hayward, P.M.; Clymo, R.S. The growth of Sphagnum: Experiments on, and simulation of some effects of light flux and water-table depth. J. Ecol. 1982, 71, 845–863. [Google Scholar] [CrossRef]
- Backéus, I. Production and depth distribution of fine roots in a boreal open bog. Ann. Bot. Fenn. 1986, 27, 261–265. [Google Scholar]
- Laiho, R.; Finer, L. Change in root biomass after water level drawdown on pine mires in southern Finland. Scand. J. For. Res. 1996, 11, 251–260. [Google Scholar] [CrossRef]
- Shetler, G.; Turetsky, M.R.; Kane, E.; Kasischke, E. Sphagnum mosses limit total carbon consumption during fire in Alaskan black spruce forests. Can. J. For. Res. 2008, 38, 2328–2336. [Google Scholar] [CrossRef]
- Thompson, D.K.; Waddington, J.M. Wildfire effects on vadose zone hydrology in forested boreal peatland microforms. J. Hydrol. 2013, 486, 48–56. [Google Scholar] [CrossRef]
- Zoltai, S.C.; Morissey, L.A.; Livingston, G.P.; Groot, W.J. Effects of fire on carbon cycling in North American boreal peatlands. Environ. Rev. 1998, 6, 13–24. [Google Scholar] [CrossRef]
- Hörnberg, G.; Zackrisson, O.; Segerström, U.; Svensson, B.W.; Ohlson, M.; Bradshaw, R.H.W. Boreal swamp forests: Biodiversity “hotspots” in an impoverished landscape. Bioscience 1998, 48, 795–802. [Google Scholar]
- Zoltai, S.C.; Taylor, S.J.; Jeglum, J.K.; Mills, G.F.; Johnson, J.D. Wetlands of Canada; National Wetlands Working Group, Polyscience Publictions Inc.: Montreal, QC, Canada, 1988; pp. 97–154. [Google Scholar]
- Lynch, J.A.; Clark, J.S.; Bigelow, N.H.; Edwards, M.E.; Finney, B.P. Geographic and temporal variations in fire history in boreal ecosystems of Alaska. J. Geophys. Res.-Atmos. 2003, 107, 8152. [Google Scholar] [CrossRef]
- Couwenberg, J.; Joosten, H. Self-organization in raised bog patterning: The origin of microtope zonation and mesotope diversity. J. Ecol. 2005, 93, 1238–1248. [Google Scholar] [CrossRef]
- Rietkerk, M.; Van de Koppel, J. Regular pattern formation in real ecosystems. Trends Ecol. Evol. 2008, 23, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Iwaniec, D.M. Regulation and Organization of Periphyton from the Florida Everglades, U.S.A. Master’s Thesis, Florida International University, Miami, FL, USA, 2008. [Google Scholar]
- Wilson, J.B.; Agnew, A.D. Positive-feedback switches in plant communities. Adv. Ecol. Res. 1992, 23, 263–336. [Google Scholar]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Gribben, P.E.; Lear, G.; He, Q.; Schiel, D.R.; Silliman, B.R.; South, P.M.; et al. Secondary foundation species enhance biodiversity. Nat. Ecol. Evol. 2018, 2, 634–639. [Google Scholar] [CrossRef] [PubMed]
- van der Zee, E.M.; Angelini, C.; Govers, L.L.; Christianen, M.J.; Altieri, A.H.; van der Reijden, K.J.; Silliman, B.R.; van de Koppel, J.; van der Geest, M.; van Gils, J.A.; et al. How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B 2016, 283, 20152326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heywood, V.H. (Ed.) Global biodiversity assessment. In United Nations Environment Program; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passé in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Byers, J.E.; Cuddington, K.; Jones, C.G.; Talley, T.S.; Hastings, A.; Lambrinos, J.G.; Crooks, J.A.; Wilson, W.G. Using ecosystem engineers to restore ecological systems. Trends Ecol. Evol. 2006, 21, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, D.J.; Zaady, E.; Shachak, M. Infiltration through three contrasting biological soil crusts in patterned landscapes in the Negev, Israel. CATENA 2000, 40, 323–336. [Google Scholar] [CrossRef]
- Rochefort, L. Sphagnum—A keystone genus in habitat restoration. Bryologist 2000, 103, 503–508. [Google Scholar] [CrossRef]
- Dudgeon, S.; Petraitis, P.S. First year demography of the foundation species, Ascophyllum nodosum, and its community implications. Oikos 2005, 109, 405–415. [Google Scholar] [CrossRef]
- Boers, A.M.; Zedler, J.B. Stabilized Water Levels and Typha Invasiveness. Wetlands 2008, 28, 676–685. [Google Scholar] [CrossRef]
- Lagerwall, G.; Kiker, G.; Muñoz-Carpena, R.; Wang, N. Accounting for the Impact of Management Scenarios on Typha domingensis (Cattail) in an Everglades Wetland. Environ. Manag. 2017, 59, 129–140. [Google Scholar] [CrossRef]
- McCormick, P.V.; Newman, S.; Vilchek, L.W. Landscape responses to wetland eutrophication: Loss of slough habitat in the Florida Everglades, USA. Hydrobiologia 2009, 621, 105–114. [Google Scholar] [CrossRef]
- Richardson, C.J.; King, R.S.; Qian, S.S.; Vaithiyanathan, P.; Qualls, R.G.; Stow, C.A. Estimating ecological thresholds for phosphorus in the Everglades. Environ. Sci. Technol. 2007, 41, 8084–8091. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, P.R.; Davis, S.E., III; van Lent, T.; Davis, S.M.; Henriquez, H. Science synthesis for management as a way to advance ecosystem restoration: Evaluation of restoration scenarios for the Florida Everglades. Restor. Ecol. 2017, 25, S4–S17. [Google Scholar] [CrossRef]
- Naja, M.; Childers, D.L.; Gaiser, E.E. Water quality implications of hydrologic restoration alternatives in the Florida Everglades, United States. Restor. Ecol. 2017, 25 (Suppl. 1), S48–S58. [Google Scholar] [CrossRef]
- Larsen, L.G.; Eppinga, M.B.; Passalacqua, P.; Getz, W.M.; Rose, K.A.; Liang, M. Appropriate complexity landscape modeling. Earth Sci. Rev. 2016, 160, 111–130. [Google Scholar] [CrossRef] [Green Version]
- Larsen, L.G.; Eppinga, M.; Coulthard, T.; Thomas, C. Exploratory modelling: Extracting causality from complexity. EOS 2014, 95, 285–292. [Google Scholar] [CrossRef]
- Zhai, L.; Jiang, J.; DeAngelis, D.; Sternberg, L.D.S.L. Prediction of plant vulnerability to salinity increase in a coastal ecosystem by stable isotope composition (δ 18 O) of plant stem water: A model study. Ecosystems 2016, 19, 32–49. [Google Scholar] [CrossRef]
- Pearlstine, L.; Friedman, S.; Supernaw, M.; Team, E.M. Everglades Landscape Vegetation Succession Model (ELVeS) Ecological and Design Document: Freshwater Marsh & Prairie Component Version 1.1; South Florida Natural Resources Center, Everglades National Park, National Park Service: Homestead, FL, USA, 2011.
- Twilley, R.R.; Rivera-Monroy, V.H. Developing Performance Measures of Mangrove Wetlands Using Simulation Models of Hydrology, Nutrient Biogeochemistry, and Community Dynamics. J. Coast. Res. 2005, 40, 79–93. [Google Scholar]
- Twilley, R.R.; Rivera-Monroy, V.H.; Chen, R.; Botero, L. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Mar. Pollut. Bull. 1999, 37, 404–419. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foundation Species | Roles Performed | |||
|---|---|---|---|---|
| Lowering Water Table | Nutrient Cycling | Habitat Provision | Erosion Control | |
| Benthic algae | x | x | ||
| Sphagnum mosses | x | x | ||
| Sasa palmata (grass) | x | x | x | |
| Betula nana (shrub) | x | (x) | x | |
| Sawgrass | x | x | x | |
| Bald cypress | x | x | x | |
| Scots pine | x | (x) | x | |
| Birch spp. | x | (x) | x | |
| Spruce spp. | x | (x) | x | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marazzi, L.; Gaiser, E.E.; Eppinga, M.B.; Sah, J.P.; Zhai, L.; Castañeda-Moya, E.; Angelini, C. Why Do We Need to Document and Conserve Foundation Species in Freshwater Wetlands? Water 2019, 11, 265. https://doi.org/10.3390/w11020265
Marazzi L, Gaiser EE, Eppinga MB, Sah JP, Zhai L, Castañeda-Moya E, Angelini C. Why Do We Need to Document and Conserve Foundation Species in Freshwater Wetlands? Water. 2019; 11(2):265. https://doi.org/10.3390/w11020265
Chicago/Turabian StyleMarazzi, Luca, Evelyn E. Gaiser, Maarten B. Eppinga, Jay P. Sah, Lu Zhai, Edward Castañeda-Moya, and Christine Angelini. 2019. "Why Do We Need to Document and Conserve Foundation Species in Freshwater Wetlands?" Water 11, no. 2: 265. https://doi.org/10.3390/w11020265
APA StyleMarazzi, L., Gaiser, E. E., Eppinga, M. B., Sah, J. P., Zhai, L., Castañeda-Moya, E., & Angelini, C. (2019). Why Do We Need to Document and Conserve Foundation Species in Freshwater Wetlands? Water, 11(2), 265. https://doi.org/10.3390/w11020265