Distribution of Pseudomonas fluorescens and Aeromonas hydrophila Bacteria in a Recirculating Aquaculture System during Farming of European Grayling (Thymallus thymallus L.) Broodstock

 , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
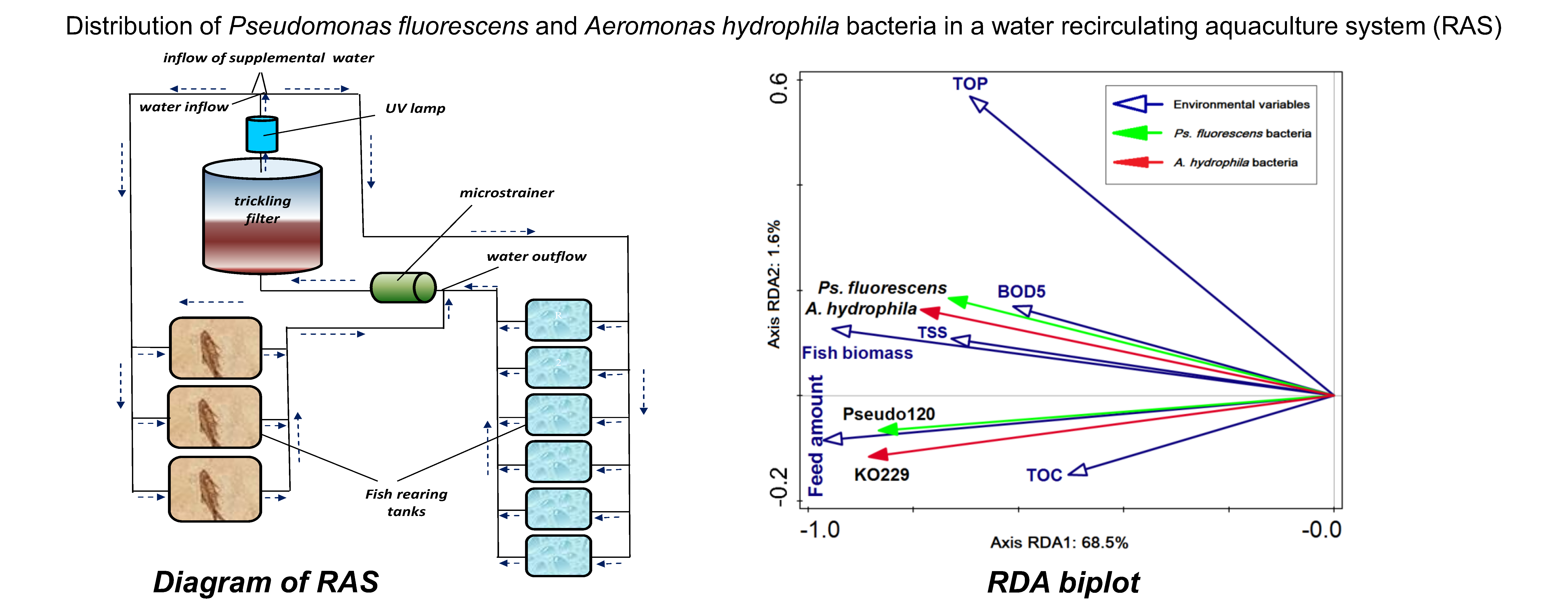
2.1. Study Area and Sampling Sites
2.2. Microbiological Analysis
2.2.1. Determination of Ps. fluorescens and A. hydrophila Counts by the Culture-Dependent Method in Samples of RAS Water and Fish Feed
2.2.2. Determination of Total Bacterial Counts in RAS Water and Fish Feed Samples by DAPI Staining and in Situ Hybridization FISH
2.3. Physicochemical Parameters of RAS Water
2.4. Statistical Analysis
3. Results and Discussion
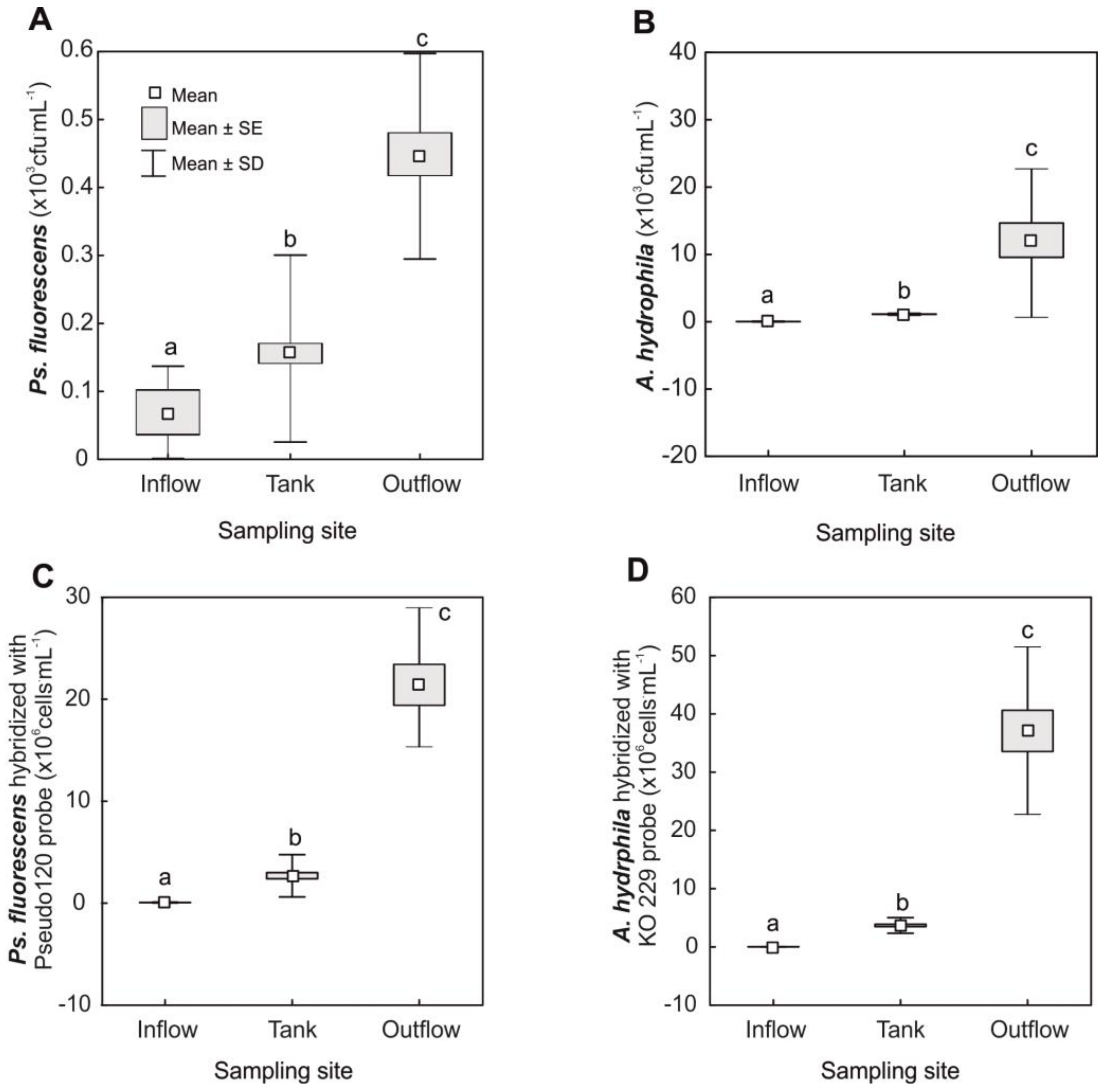
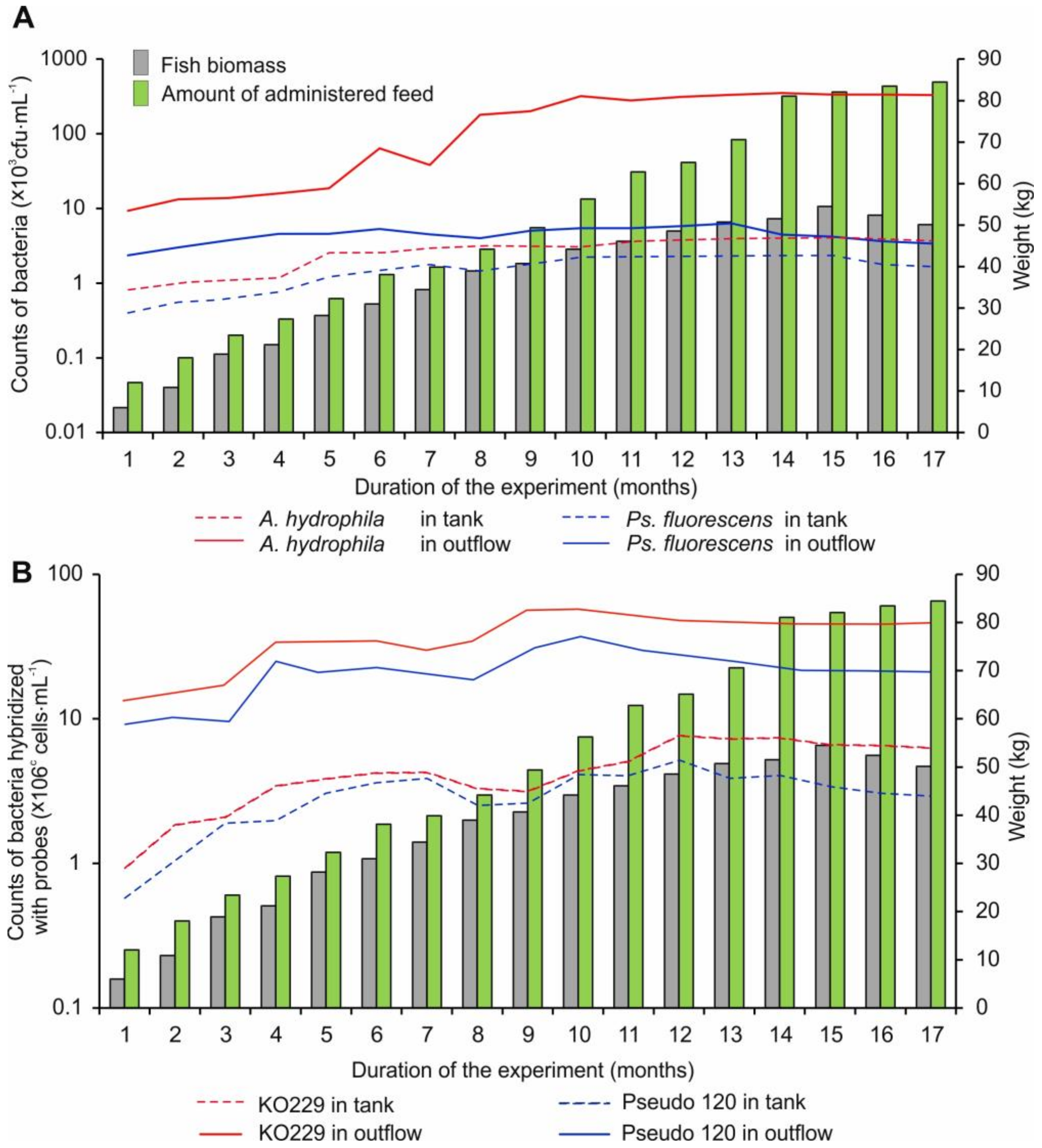
3.1. Ps. fluorescens and A. hydrophila Counts in Samples of Water and Fish Feed
3.2. Physicochemical Parameters of Water Samples
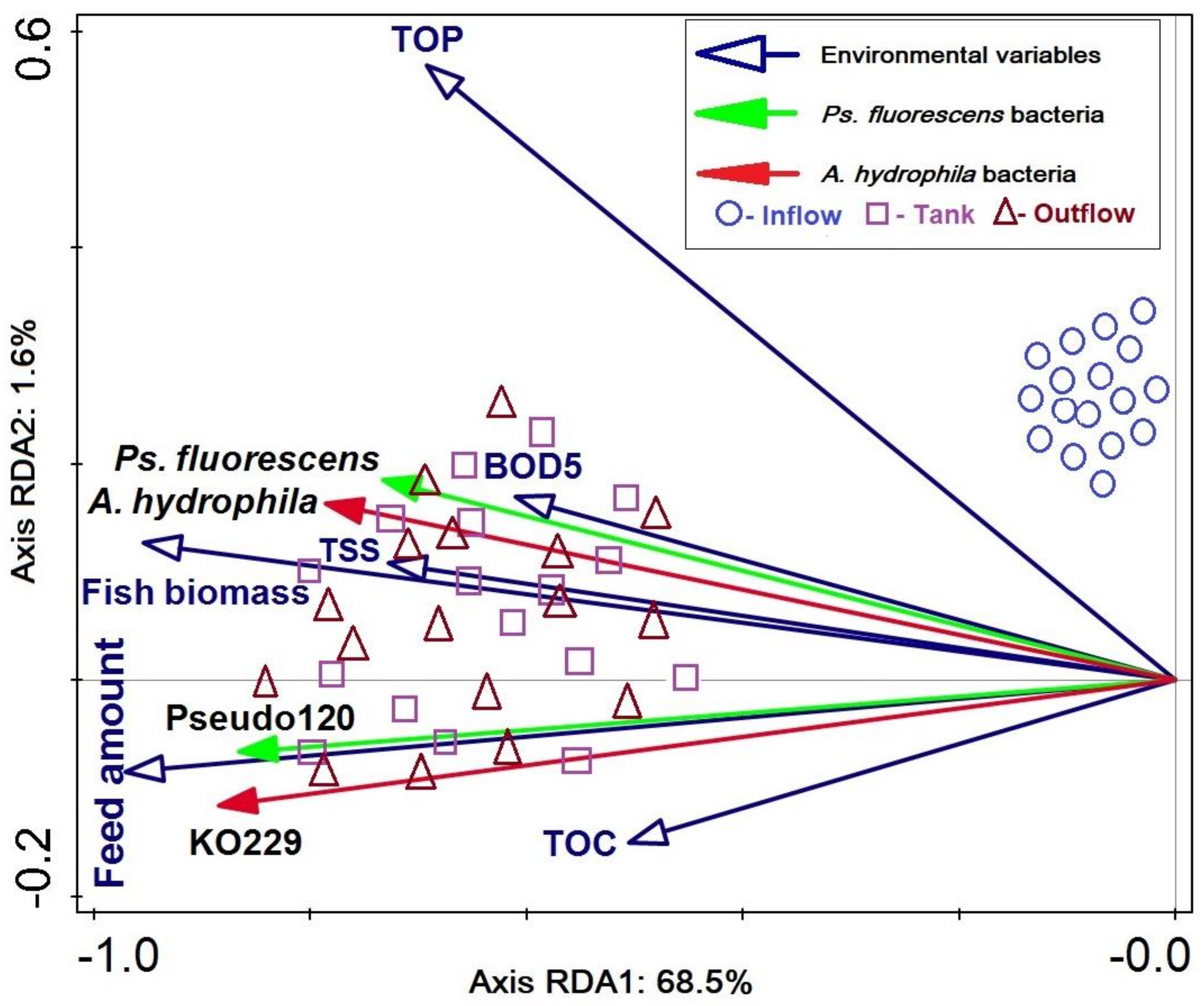
3.3. Relationships between Bacterial Counts and Environmental Variables
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bakke, I.; Skjermo, J.; Vo, T.A.; Vadstein, O. Live feed is not a major determinant of the microbiota associated with cod larvae (Gadus morhua). Environ. Microbiol. Rep. 2013, 5, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Heilig, H.; Benvenuti, G.; Verreth, J.; Verdegem, M. The impact of rearing environment on the development of gut microbiota in tilapia larvae. Sci. Rep. 2015, 5, 18206. [Google Scholar] [CrossRef] [PubMed]
- Terech-Majewska, E.; Pajdak, J.; Siwicki, A.K. Water as a source of macronutrients and micronutrients for fish, with special emphasis on the nutritional requirements of two fish species: The common carp (Cyprinus carpio) and the rainbow trout (Oncorhynchus mykiss). J. Elem. 2016, 21, 947–961. [Google Scholar]
- Harnisz, M.; Gołaś, I.; Pietruk, M. Tetracycline-resistant bacteria as indicators of antimicrobial resistance in protected waters—The example of the Drwęca River nature reserve (Poland). Ecol. Indic. 2011, 11, 663–668. [Google Scholar] [CrossRef]
- Harnisz, M.; Korzeniewska, E.; Gołaś, I. The impact of a freshwater fish farm on the community of tetracycline-resistant bacteria and the structure of tetracycline resistance genes in river water. Chemosphere 2015, 128, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Terech-Majewska, E. Improving disease prevention and treatment in controlled fish culture. Arch. Pol. Fish. 2016, 24, 115–165. [Google Scholar] [CrossRef]
- Cytryn, E.; Gelfand, I.; Barak, Y.; van Rijn, J.; Minz, D. Diversity of microbial communities correlated to physiochemical parameters in a digestion basin of a zero-discharge mariculture system. Environ. Microbiol. 2003, 5, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Auffret, M.; Yergeau, E.; Pilote, A.; Proulx, E.; Proulx, D.; Greer, C.W.; Vandenberg, G.; Villemur, R. Impact of water quality on the bacterial populations and off-flavours in recirculating aquaculture systems. FEMS Microbiol. Ecol. 2013, 84, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Blancheton, J.P.; Attramadal, K.J.K.; Michaud, L.; Roque d’Orbcastel, E.; Vadstein, O. Insight into bacterial population in aquaculture systems and its implication. Aquacult. Eng. 2013, 53, 30–39. [Google Scholar] [CrossRef]
- Gołaś, I.; Zmysłowska, I.; Harnisz, M.; Teodorowicz, M. The microbiological state of fish feed, water and Silurus glanis L. skin of fry during intensive rearing. Bull. Sea Fish. Inst. 2004, 1, 3–14. [Google Scholar]
- Krause, J.; Zmysłowska, I.; Gołaś, I. Potentially pathogenic bacteria in water and Siberian sturgeon (Acipenser baeri Br.) with Russian sturgeon (Acipenser gueldenstaedti Br.) hybrids in a closed water cycle. Bull. Sea Fish. Inst. 2005, 3, 65–77. [Google Scholar]
- Krause, J.; Zmysłowska, I.; Gołaś, I.; Szarek, J. Qualitative composition of bacterial microflora of tank water and hybrid of Siberian sturgeon (Acipenser baeri Br.) with Russian sturgeon (Acipenser gueldenstaedti Br.) from intensive rearing in a closed water system. Pol. J. Natur. Sci. 2006, 20, 345–354. [Google Scholar]
- Sugita, H.; Nakamura, H.; Shimada, T. Microbial communities associated with filter material in recirculating aquaculture systems of freshwater fish. Aquaculture 2005, 243, 403–409. [Google Scholar] [CrossRef]
- Kawahara, N.; Shigematsu, K.; Miyadai, T.; Kondo, R. Comparison of bacterial communities in fish farm sediments along organic enrichment gradient. Aquaculture 2009, 287, 107–113. [Google Scholar] [CrossRef]
- Bacelar-Nicolau, P.; Nicolau, L.B.; Marques, J.C.; Morgado, F.; Pastorinho, R.; Azeiteiro, U.M. Bacterioplankton dynamics in the Mondego eustary (Portugal). Acta Oecol. 2003, 24, S67–S75. [Google Scholar] [CrossRef]
- Thayumanavan, T.; Vivekanandhan, G.; Savithamani, K.; Subashkumar, R.; Lakshmanaperumalsamy, P. Incidence of haemolysin-positive and drug-resistant Aeromonas hydrophila in freshly caught fin fish and prawn collected from major commercial fishes of coastal South India. FEMS Immunol. Med. Mic. 2003, 36, 41–45. [Google Scholar] [CrossRef]
- Zhou, Q.L.; Li, K.; Jun, X.; Bo, L. Role and functions of beneficial microorganisms in sustainable aquaculture. Bioresour. Technol. 2009, 100, 3780–3786. [Google Scholar] [CrossRef] [PubMed]
- Roca, C.; Olsson, L. Dynamic responses of Pseudomonas fluorescens DF57 to nitrogen or carbon source addition. J. Biotechnol. 2001, 86, 39–50. [Google Scholar] [CrossRef]
- Saha, R.S.; Donofrio, D.; Goeres, M.; Bagley, S.T. Rapid detection of rRNA group I pseudomonads in contaminated metalworking fluids and biofilm formation by fluorescent in situ hybridization. Appl. Microbiol. Biotechnol. 2012, 94, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Pianetti, A.; Battistelli, M.; Citterio, B.; Parlani, C.; Falcieri, E.; Bruscolini, F. Morphological changes of Aeromonas hydrophila in response to osmotic stress. Micron 2009, 40, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Picot, L.; Abdelmoula, S.M.; Mérieau, A.; Orange, N.; Feuilloley, M.G.J. The psychotropic bacterium Pseudomonas fluorescens can adapt to human physiologic temperature and behaves as a pathogen in central and peripheral nerve cells. Eur. J. Cell Biol. 2000, 52, 129–132. [Google Scholar]
- Picot, L.; Abdelmoula, S.M.; Merieau, A.; Leroux, P.; Cazin, L.; Orange, N.; Feuilloley, M.G.J. Pseudomonas fluorescens as a potential pathogen: Adherence to nerve cells. Microbes Infect. 2001, 3, 985–995. [Google Scholar] [CrossRef]
- Shayo, S.D.; Mwita, C.J.; Hosea, K.M. Virulence of Pseudomonas and Aeromonas bacteria recovered from Oreochromis niloticus (Perege) from Mtera hydropower Dam; Tanzania. Ann. Biol. Res. 2012, 3, 5157–5161. [Google Scholar]
- Holmes, P.; Sartory, D.P. An evaluation of media for the membrane filtration enumeration of Aeromonas from drinking water. Lett. Appl. Microbiol. 1993, 17, 58–60. [Google Scholar] [CrossRef]
- Johnsen, K.; Nielsen, P. Diversity of Pseudomonas strains isolated with King’s B and Gould’s S1 agar determined by repetitive extragenic palindromic-polymerase chain reaction, 16S rDNA sequencing and Fourier transform infrared spectroscopy characterisation. FEMS Microbiol. Lett. 1999, 173, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.; Snaidr, J.; Wagner, M.; Ludwig, W.; Schleifer, K.H. In situ visualization of high genetic diversity in a natural microbial community. J. Bacteriol. 1996, 178, 3496–3500. [Google Scholar] [CrossRef] [PubMed]
- Franke-Whittle, I.; Klammer, S.; Insam, H. Design and application of an oligonucleotide microarray for the investigation of compost microbial communities. J. Microbiol. Methods 2005, 62, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Gotkowska-Płachta, A.; Gołaś, I.; Korzeniewska, E.; Koc, J.; Rochwerger, A.; Solarski, K. Evaluation of the distribution of faecal indicator bacteria in a river system depending on different types of land use in the southern watershed of the Baltic Sea. ESPR 2016, 23, 4073–4085. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Olsen, R.E. The effect of diet on aerobic bacterial flora associated with intestine of Arctic charr (Salvelinus alpinus L.). J. Appl. Microbiol. 1999, 86, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, H.; Sekiguchi, Y.; Hanada, S.; Nakamura, K.; Nomura, N.; Matsumura, M.; Kamagata, Y. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques. Appl. Environ. Microbiol. 2005, 71, 2162–2169. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J.P.; Lamouroux, N.; Sagnes, P.; Persat, H. Habitat preferences of European grayling in a medium size stream, the Ain river, France. J. Fish Biol. 2000, 56, 1312–1326. [Google Scholar] [CrossRef]
- Ovidio, M.; Philippart, J.C. The impact of small physical obstacles on upstream movements of six species of fish. Hydrobiologia 2002, 483, 55–69. [Google Scholar] [CrossRef]
- Penczak, T.; Kruk, A. Threatened obligatory riverine fishes in human-modified Polish rivers. Ecol. Freshwater Fish 2000, 9, 109–117. [Google Scholar] [CrossRef]
- Wiśniewolski, W. Factors supporting and preventing the development and maintenance of fish populations in flowing waters. Suppl. Acta Hydrobiol. 2002, 3, 1–28. [Google Scholar]
- Augustyn, L.; Nowak, M. Assessment of populations of European grayling, Thymallus thymallus (L.) in the Dunajec River catchment based on recreational catch records. Kom. Ryb. 2014, 4, 9–15. [Google Scholar]
- Fjeld, E.; Haugen, T.O.; Vøllestad, L.A. Permanent impairment in the feeding behavior of grayling (Thymallus thymallus) exposed to methylmercury during embryogenesis. Sci. Total Environ. 1998, 213, 247–254. [Google Scholar] [CrossRef]
- Honkanen, J.O.; Kostamo, A.; Kukkonen, J.V.K. Toxicity of a phytosterol mixture to grayling (Thymallus thymallus) during early developmental stages. Arch. Environ. Contam. Toxicol. 2005, 48, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Vuorinen, P.J.; Peuranen, S.; Tigerstedt, C. Effects of iron, aluminium, dissolved humic material and acidity of grayling (Thymallus thymallus) in laboratory exposures, and a comparison of sensitivity with brown trout (Salmo trutta). Boreal Environ. Res. 1998, 3, 405–419. [Google Scholar]
- Horka, P.; Ibbotson, A.; Jones, J.I.; Cove, R.J.; Scott, L.J. Validation of scale-age determination in European grayling Thymallus thymallus using tag-recapture analysis. J. Fish Biol. 2010, 77, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, A.; Błachuta, J.; Kokurewicz, B.; Kowalewski, M. Changes of gonadosomatic index (GSI) and egg diameter in the Thymallus thymallus (L.) in annual cycle. Acta Ichtyol. Pisc. 1989, 19, 21–28. [Google Scholar] [CrossRef]
- Witkowski, A.; Kotusz, J.; Przybylski, M. The degree of threat to the freshwater ichthyofauna of Poland: 2009 Red List of Fish and Lamprey Species. Chrońmy Przyr. Ojcz. 2009, 65, 33–52. (In Polish) [Google Scholar]
- Szmyt, M.; Niewiadomski, P. Evaluation of European grayling (Thymallus thymallus L.) farming in recirculating systems in view of active protection measures. In Proceedings of the 24th National Symposium on Comprehensive Research and Protection of the Natural Environment, Tleń, Poland, 26–27 June 2017; 2017; pp. 237–249. (In Polish). [Google Scholar]
- Szmyt, M.; Lejk, A.M.; Ciereszko, A.; Woźniak, M.; Dietrich, G.J.; Judycka, S.; Roszuk, J.; Polewacz, A.; Lampkowski, M.; Pasturczak, F.; et al. Broodstock selection and an evaluation of controlled reproduction in the European grayling (Thymallus thymallus L.) in a recirculating aquaculture system (RAS). In Proceedings of the 60th Conference of Salmonid Breeders, Gdynia, Poland, 8–9 October 2015. [Google Scholar]
- Gołaś, I.; Korzekwa, K.; Harnisz, M.; Zmysłowska, I.; Teodorowicz, M.; Terech-Majewska, E.; Rodziewicz, W.; Bieńkowska, M. Influence of fishery management and environmental factors on occurrence of heterotrophic, hemolytic and mesophilic bacteria and Aeromonas hydrophila in waters of Drwęca River, Poland. Arch. Environ. Prot. 2009, 35, 27–40. [Google Scholar]
- Michaud, L.; Lo Giudice, A.; Troussellier, M.; Smedile, F.; Bruni, V.; Blancheton, J.P. Phylogenetic characterization of the heterotrophic bacterial communities inhabiting a marine recirculating aquaculture system. J. Appl. Microbiol. 2009, 107, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- APHA. Standard Methods for the Examination of Water and Wastewater, 15th ed.; Water Pollution Control Federation: New York, NY, USA, 1992. [Google Scholar]
- Loy, A.; Maixner, F.; Wagner, M.; Horn, M. ProbeBase—An online resource for rRNA-targeted oligonucleotide probes: New features 2007. Nucleic Acids Res. 2007, 3, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, J.F.; Glöckner, O.; Schönhuber, W.; Amann, R. Fluorescence in situ hybridization with rRNA-targeted oligonucleotide probes. In Fluorescence In Situ Hybridization, Methods in Microbiology: Marine Microbiology; Pernthaler, J.F., Glöckner, O., Schönhuber, W., Amann, R., Paul, J., Eds.; Academic Press Ltd.: London, UK, 2001; Volume 30, pp. 207–226. [Google Scholar]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of Proteobacteria: Problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16SrRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [PubMed]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization with rRNA-targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Brosius, J.; Dull, T.J.; Sleeter, D.D.; Noller, H.F. Gene organization and primary structure of a ribosomal RNA operon from Escherichia coli. J. Mol. Biol. 1981, 148, 107–127. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: New York, NY, USA, 2002. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and Canodraw for Windows User’s Guide, Software for Canonical Community Ordination, version 4.5; Microcomputer Power: Ithaca, NY, USA, 2002; p. 500. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using Canoco; Cambridge University Press: Cambridge, UK, 2003; p. 269. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012; 496p. [Google Scholar]
- Leonard, N.; Blancheton, J.P.; Guiraud, J.P. Populations of heterotrophic bacteria in an experimental recirculating aquaculture system. Aquacult. Eng. 2000, 22, 109–120. [Google Scholar] [CrossRef]
- Wietz, M.; Hall, M.R.; Hoj, L. Effects of seawater ozonation on biofilm development in aquaculture tanks. Syst. Appl. Microbiol. 2009, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.; Borges, M.-T.; Peixe, C.; Henriques, I.; Pereira, C.M.; Castro, P.M.L. A molecular and multivariate approach to the microbial community of a commercial shallow raceway marine recirculation system operating with a moving bed biofilter. Aquac. Res. 2011, 42, 1308–1322. [Google Scholar] [CrossRef]
- Keller, M.; Zengler, K. Tapping into microbial diversity. Nat. Rev. Microbiol. 2004, 2, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Kisand, V.; Wikner, J. Combining culture dependent and -independent methodologies for estimation of richness of estuarine bacterioplankton consuming riverine dissolved organic matter. Appl. Environ. Microbiol. 2003, 69, 3607–3616. [Google Scholar] [CrossRef] [PubMed]
- Avnimelech, Y. Carbon and nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Beveridge, M.C.M.; Philips, M.J.; Macintosh, D.C. Aquaculture and environment: The supply and demand for environment goods and services by Asian aquaculture and the implications for sustainability. Aquac. Res. 1997, 28, 101–111. [Google Scholar] [CrossRef]
- Sidoruk, M.; Cymes, I. Effect of water management technology used in trout culture on water quality in fish ponds. Water 2018, 10, 1264. [Google Scholar] [CrossRef]
- Boyd, C.E. Guidelines for aquaculture effluent management at the farm-level. Aquaculture 2003, 226, 101–112. [Google Scholar] [CrossRef]
- Michaud, L.; Blancheton, J.P.; Bruni, V.; Piedrahita, R. Effect of particulate organic carbon on heterotrophic bacterial populations and nitrification efficiency in biological filters. Aquacult. Eng. 2006, 34, 224–233. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe | Target Species | Target1 Site (rRNA Positions) | % FA2 in situ | TF3 | Reference |
|---|---|---|---|---|---|
| EUB338 | Bacteria | 16S (338–355) | 35 | 48 | [50] |
| NON338 | Control probe complementary to EUB338 | 16S (338–355) | 35 | 48 | [51] |
| GAM42a | Gammaproteobacteria | 23S (1024–1043) | 35 | 47 | [49] |
| GAM42a’ | Gamma competitor | 23S (1024–1043) | 35 | 47 | [49] |
| Pseudo120 | Ps. fluorescens | 16 S (120–142) | 30 | 57 | [19] |
| KO 229 | A. hydrophila | 16S (454–474) | 30 | 49 | [27] |
| Bacteria | Inflow 1 | Tank 1 | Outflow 1 | |||
|---|---|---|---|---|---|---|
| Mean | Range | Mean | Range | Mean | Range | |
| Ps. fluorescens2 | 65 | 20–145 | 141 | 40–250 | 446 | 220–930 |
| A. hydrophila2 | 26 | 1–76 | 244 | 90–342 | 11,000 | 930–35,000 |
| Total bacterial counts 3 | 39.4 | 12.3–87.2 | 85.6 | 42.4–202.1 | 303.4 | 152.3–559.1 |
| EUB338 (%) 4 | 66.0 | 27.1–69.2 | 61.7 | 16.6–69.0 | 67.2 | 63.8–79.1 |
| GAM42a (%) 4 | 8.2 | 6.3–18.5 | 10.3 | 9.8–21.2 | 15.1 | 9.4–28.6 |
| Pseudo120 (%) 4 | 0.17 | 0.1–0.3 | 2.6 | 1.5–3.9 | 4.6 | 3.3–6.4 |
| KO229 (%) 4 | 0.07 | 0.05–0.10 | 5.1 | 1.8–5.6 | 9.9 | 7.3–11.1 |
| Parameter (Unit) | Sampling Sites | ||
|---|---|---|---|
| Inflow | Tank | Outflow | |
| T (°C) | 10.0 ± 2.5 | 10.6 ± 2.4 | 10.6 ± 2.3 |
| DO (mg·L−1) | 10.03 ± 0.53 | 9.9 ± 0.6 | 9.01 ± 0.61 |
| OS (% O2) | 90.9 ± 3.7 | 84.6 ± 21.4 | 80.1 ± 3.2 |
| pH | 7.9 ± 0.14 | 8.0 ± 0.15 | 8.20 ± 0.15 |
| PO4-P (mg P·L−1) | 0.545 ± 0.152 | 0.554 ± 0.161 | 0.596 ± 0.121 |
| TOP (mg P·L−1) | 0.164 ± 0.051a | 0.233 ± 0.059b | 0.281 ± 0.064b |
| TP (mg P·L−1) | 0.710 ± 0.164 | 0.784 ± 0.188 | 0.910 ± 0.191 |
| NH4-N (mg N·L−1) | 0.038 ± 0.017 | 0.065 ± 0026 | 0.075 ± 0.070 |
| NH3-N (mg N·L−1) | 0.002 ± 0.001 | 0.003 ± 0.001 | 0.003 ± 0.001 |
| NO2-N (mg N·L−1) | 0.004 ± 0.003 | 0.018 ± 0.009 | 0.011 ± 0.001 |
| NO3-N (mg N·L−1) | 8.848 ± 2.385 | 8.455 ± 2.727 | 8.152 ± 2.837 |
| TON (mg N·L−1) | 3.041 ± 1.147 | 4.576 ± 1.203 | 4.733 ± 1.282 |
| TN (mg N·L−1) | 11.83 ± 2.93 | 13.10 ± 2.95 | 13.17 ± 2.95 |
| BOD5 (mg O2·L−1) | 1.95 ± 0.47a | 2.64 ± 0.57b | 3.82 ± 0.67b |
| TSS (mg·L−1) | 1.94 ± 0.78a | 4.29 ± 1.69b | 4.86 ± 2.02b |
| TOC (mg C·L−1) | 3.01 ± 0.33a | 4.31 ± 0.62b | 4.45 ± 0.71b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gołaś, I.; Szmyt, M.; Potorski, J.; Łopata, M.; Gotkowska-Płachta, A.; Glińska-Lewczuk, K. Distribution of Pseudomonas fluorescens and Aeromonas hydrophila Bacteria in a Recirculating Aquaculture System during Farming of European Grayling (Thymallus thymallus L.) Broodstock. Water 2019, 11, 376. https://doi.org/10.3390/w11020376
Gołaś I, Szmyt M, Potorski J, Łopata M, Gotkowska-Płachta A, Glińska-Lewczuk K. Distribution of Pseudomonas fluorescens and Aeromonas hydrophila Bacteria in a Recirculating Aquaculture System during Farming of European Grayling (Thymallus thymallus L.) Broodstock. Water. 2019; 11(2):376. https://doi.org/10.3390/w11020376
Chicago/Turabian StyleGołaś, Iwona, Mariusz Szmyt, Jacek Potorski, Michał Łopata, Anna Gotkowska-Płachta, and Katarzyna Glińska-Lewczuk. 2019. "Distribution of Pseudomonas fluorescens and Aeromonas hydrophila Bacteria in a Recirculating Aquaculture System during Farming of European Grayling (Thymallus thymallus L.) Broodstock" Water 11, no. 2: 376. https://doi.org/10.3390/w11020376
APA StyleGołaś, I., Szmyt, M., Potorski, J., Łopata, M., Gotkowska-Płachta, A., & Glińska-Lewczuk, K. (2019). Distribution of Pseudomonas fluorescens and Aeromonas hydrophila Bacteria in a Recirculating Aquaculture System during Farming of European Grayling (Thymallus thymallus L.) Broodstock. Water, 11(2), 376. https://doi.org/10.3390/w11020376