Expected Shifts in Nekton Community Following Salinity Reduction: Insights into Restoration and Management of Transitional Water Habitats

 , ,
, , 

 and
and
Abstract
:1. Introduction
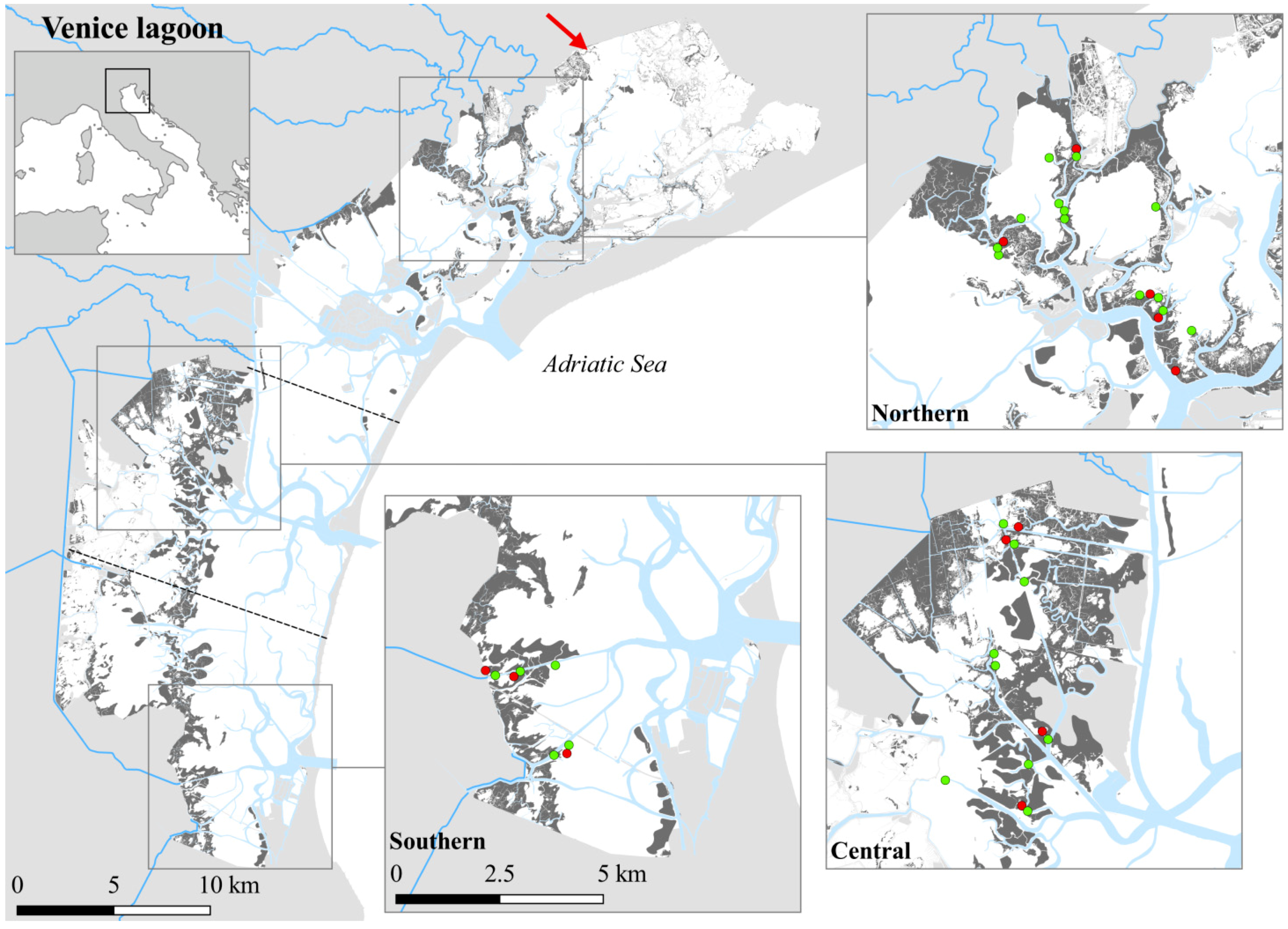
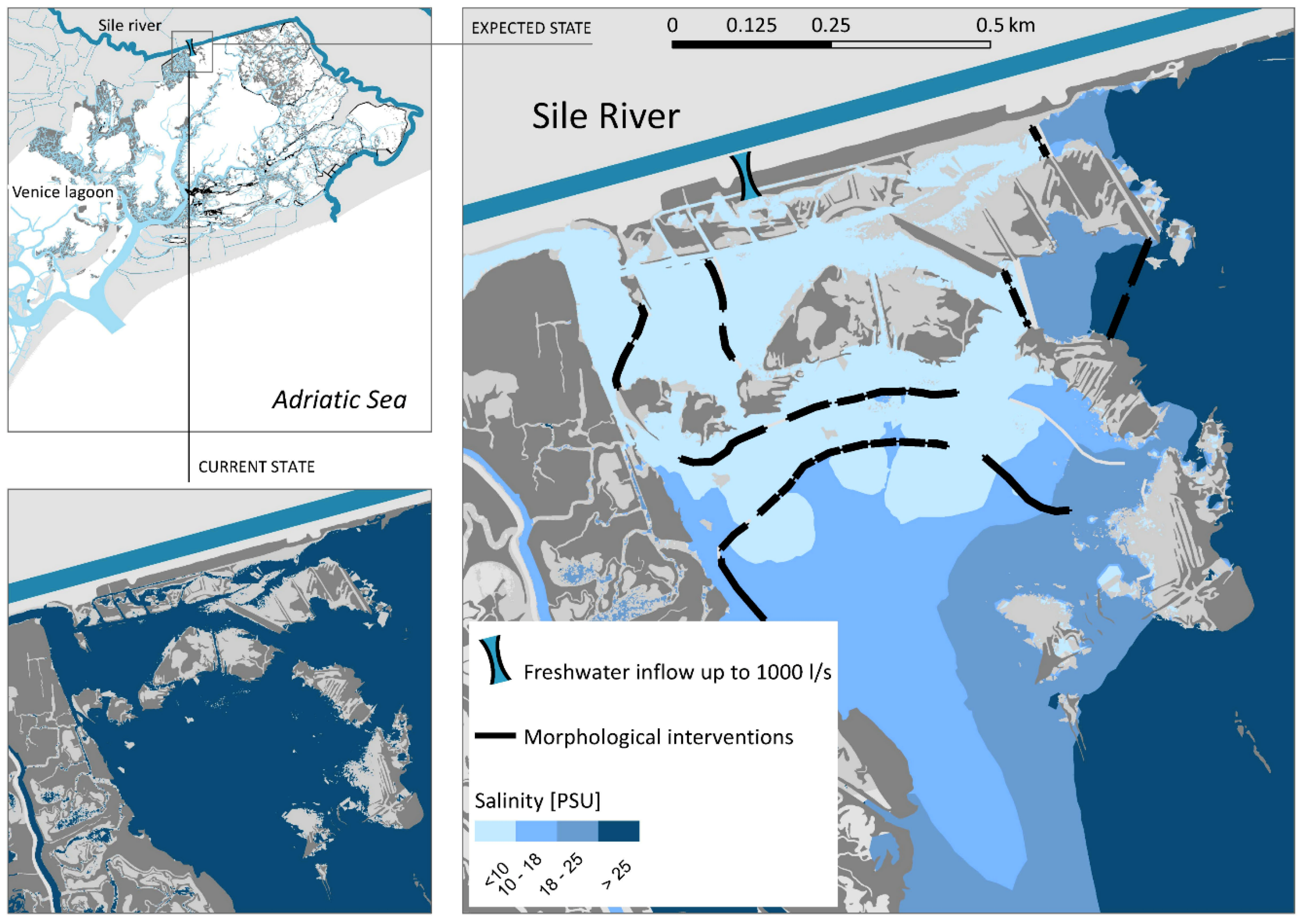
2. Materials and Methods
3. Results
3.1. Environmental Variability
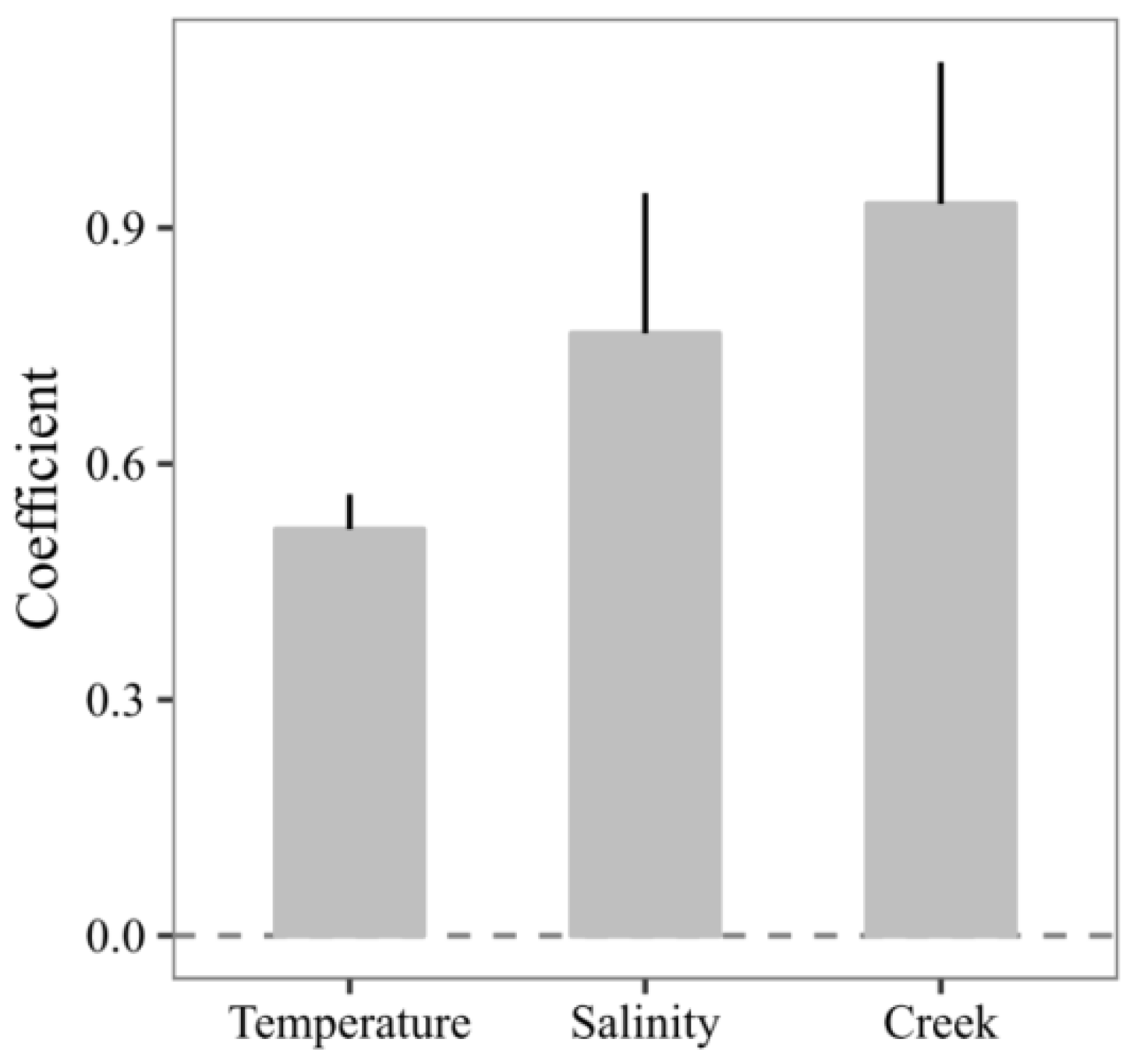
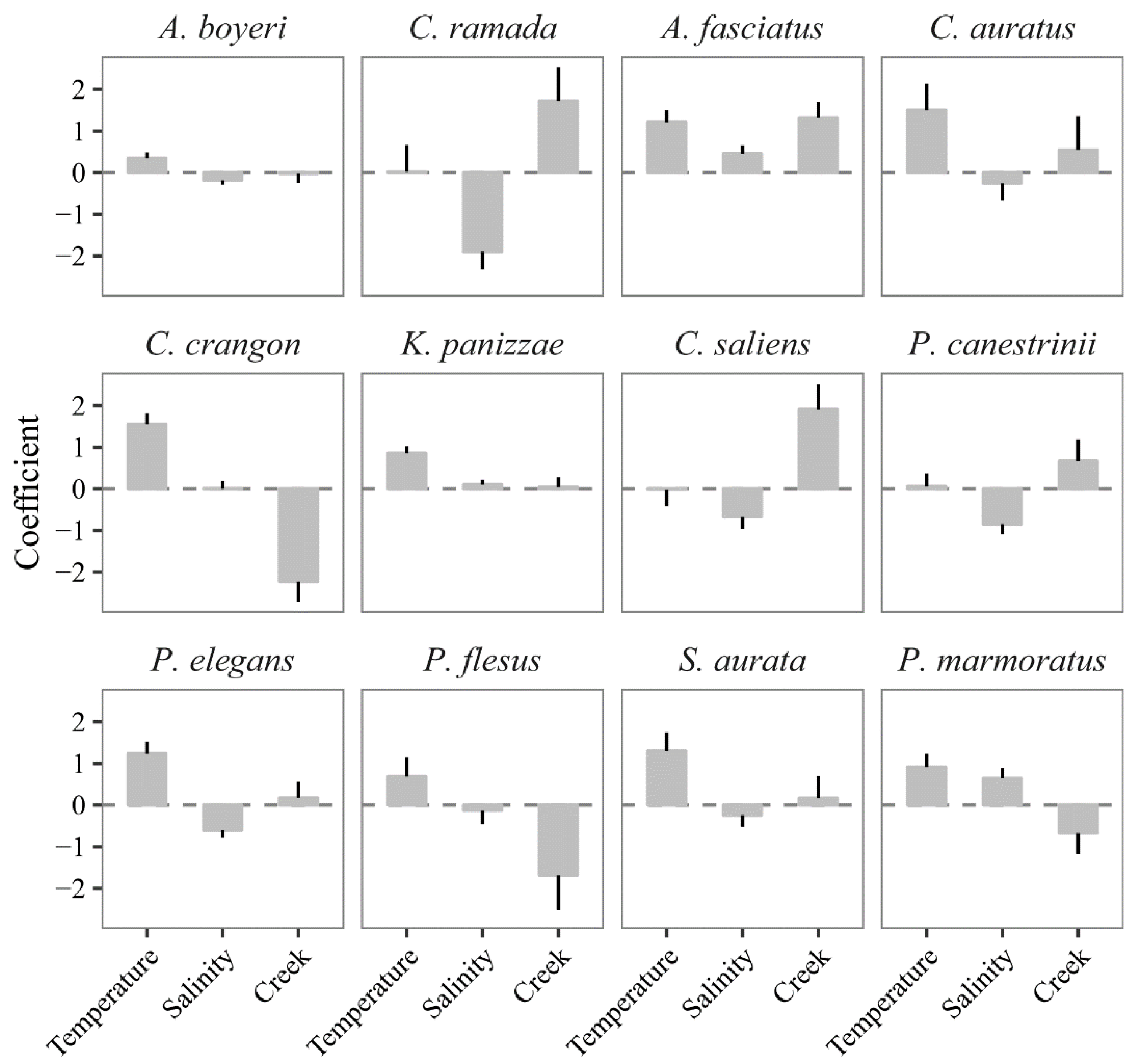
3.2. Model Calibration
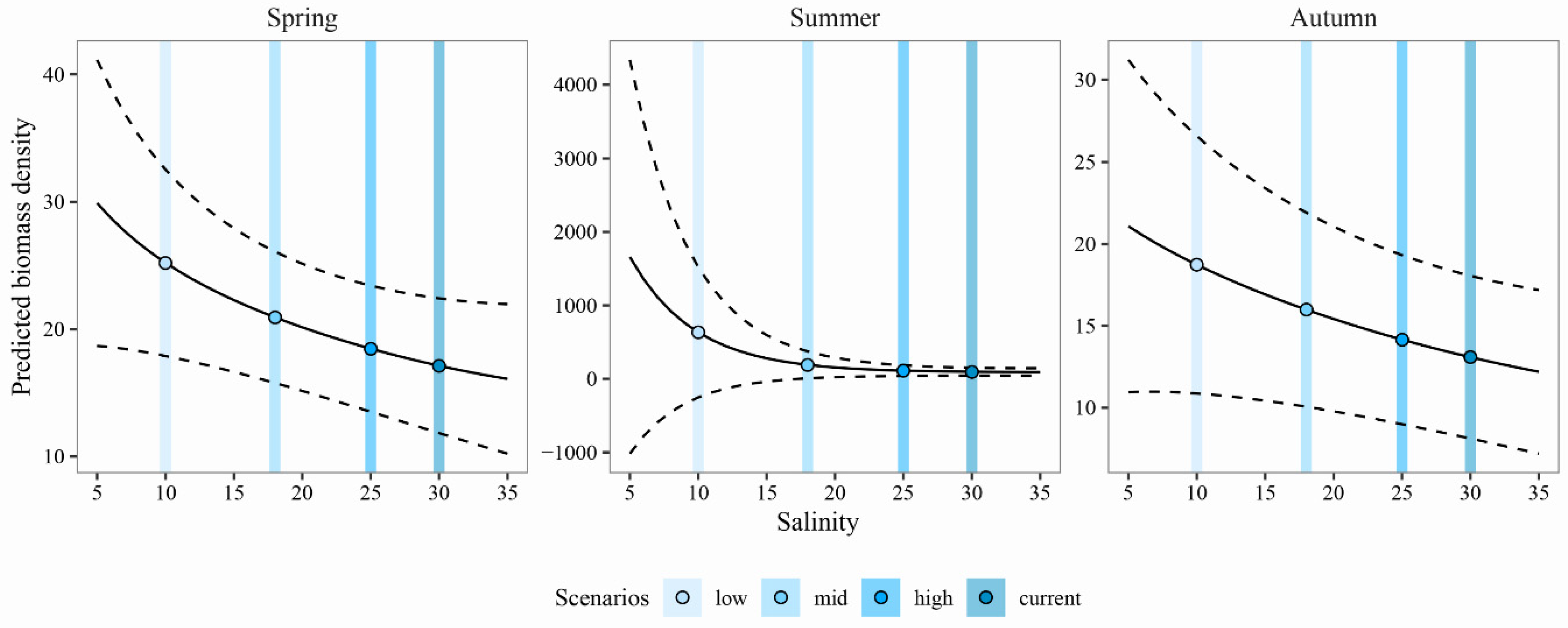
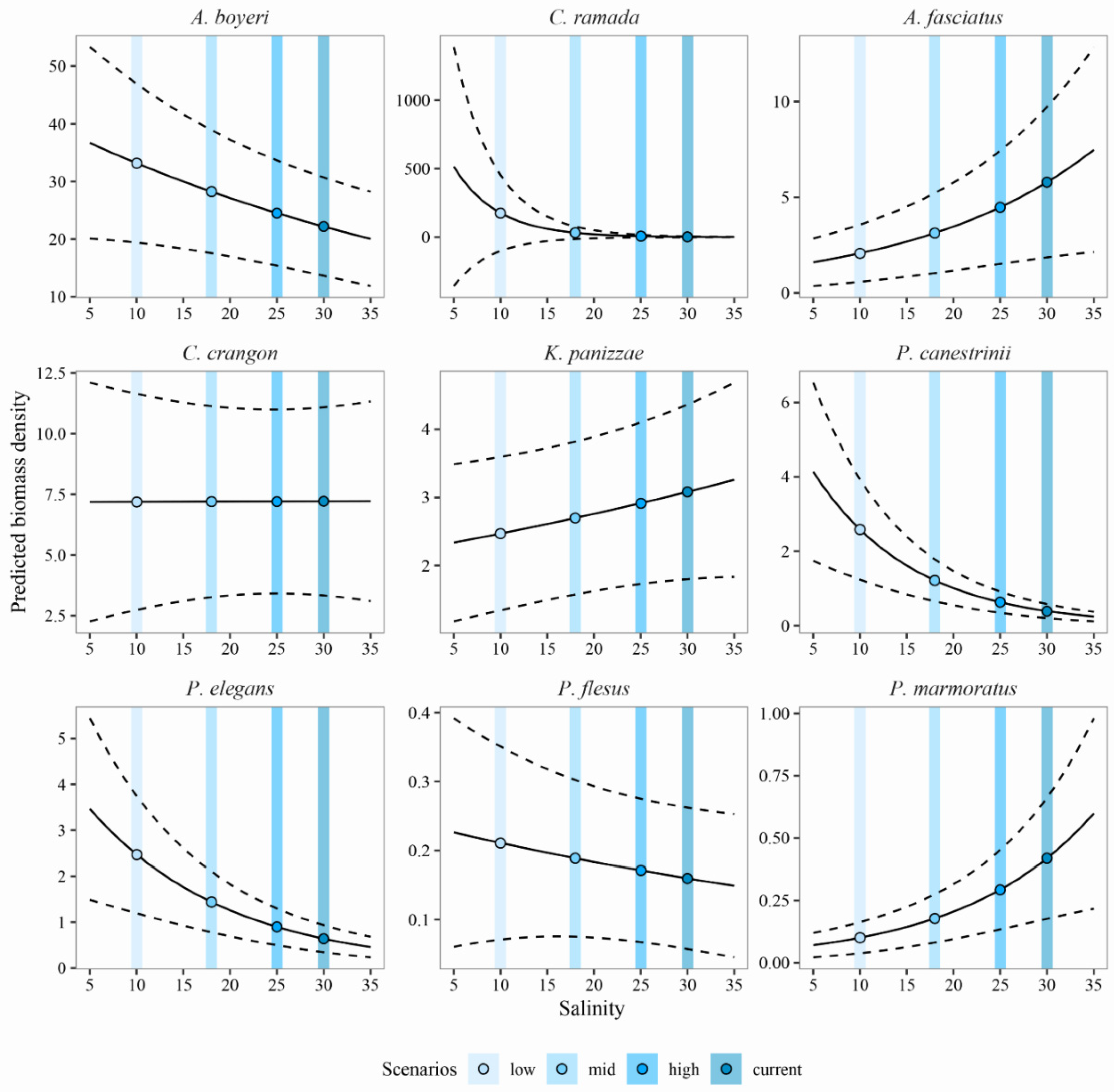
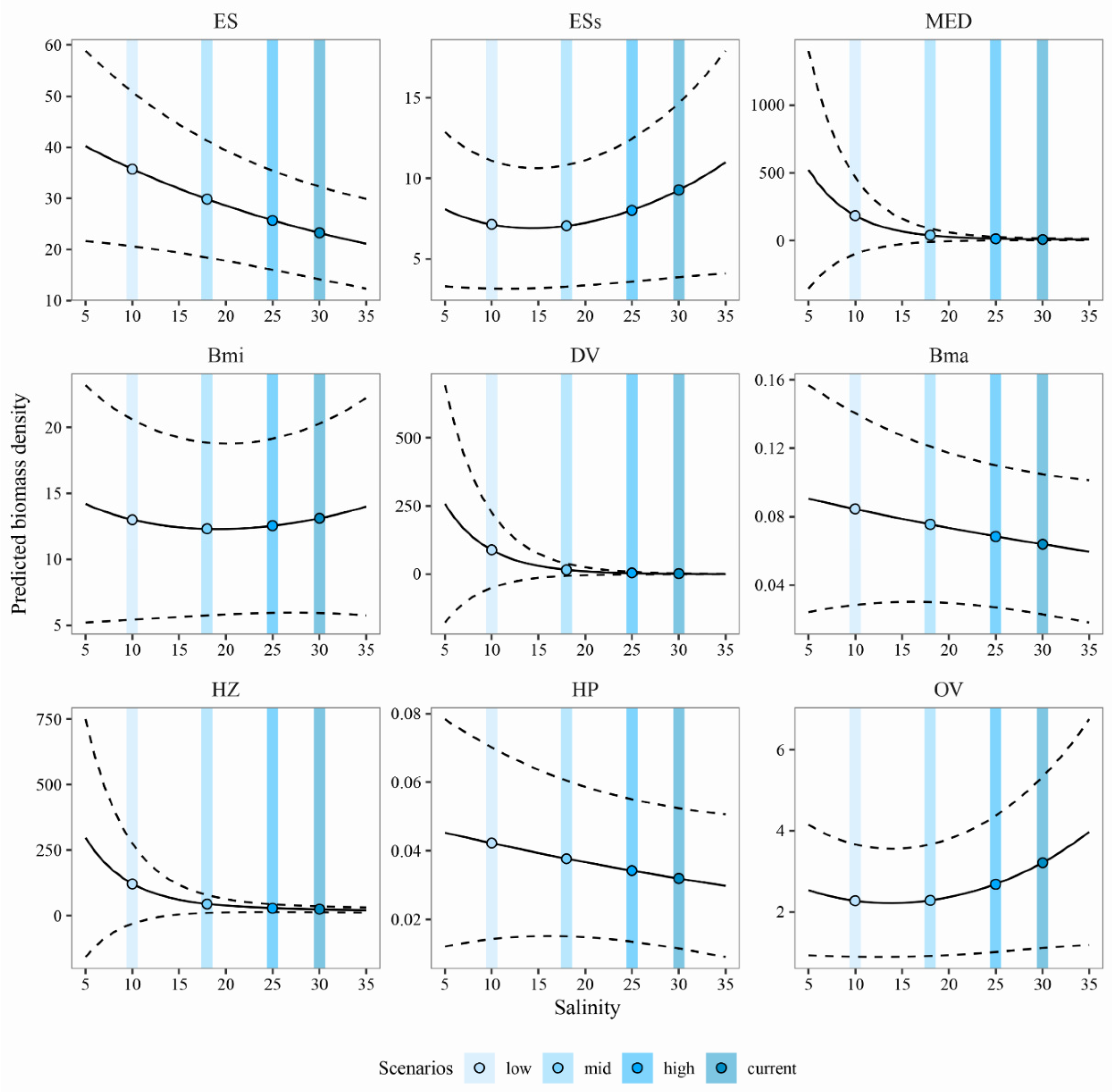
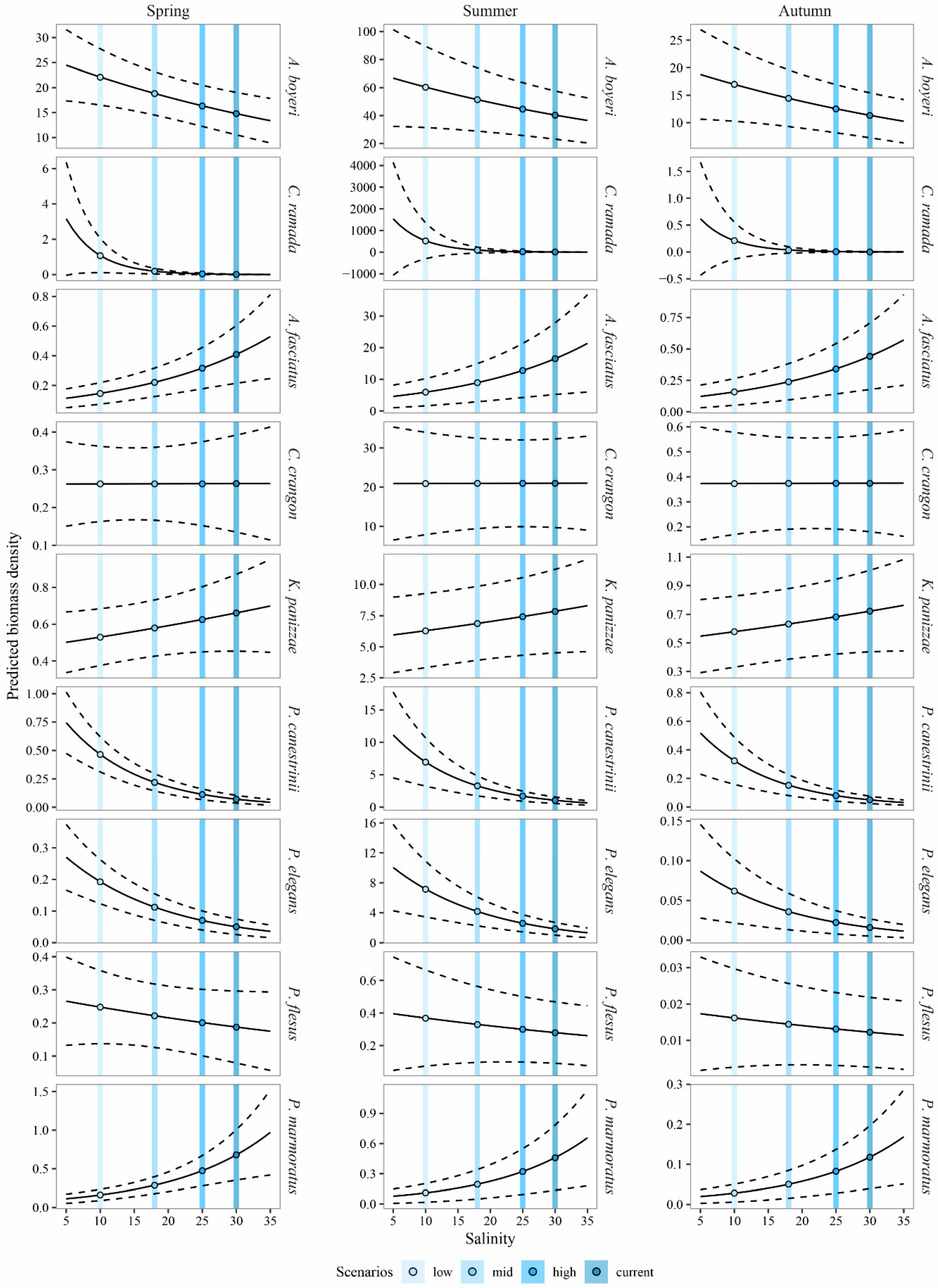
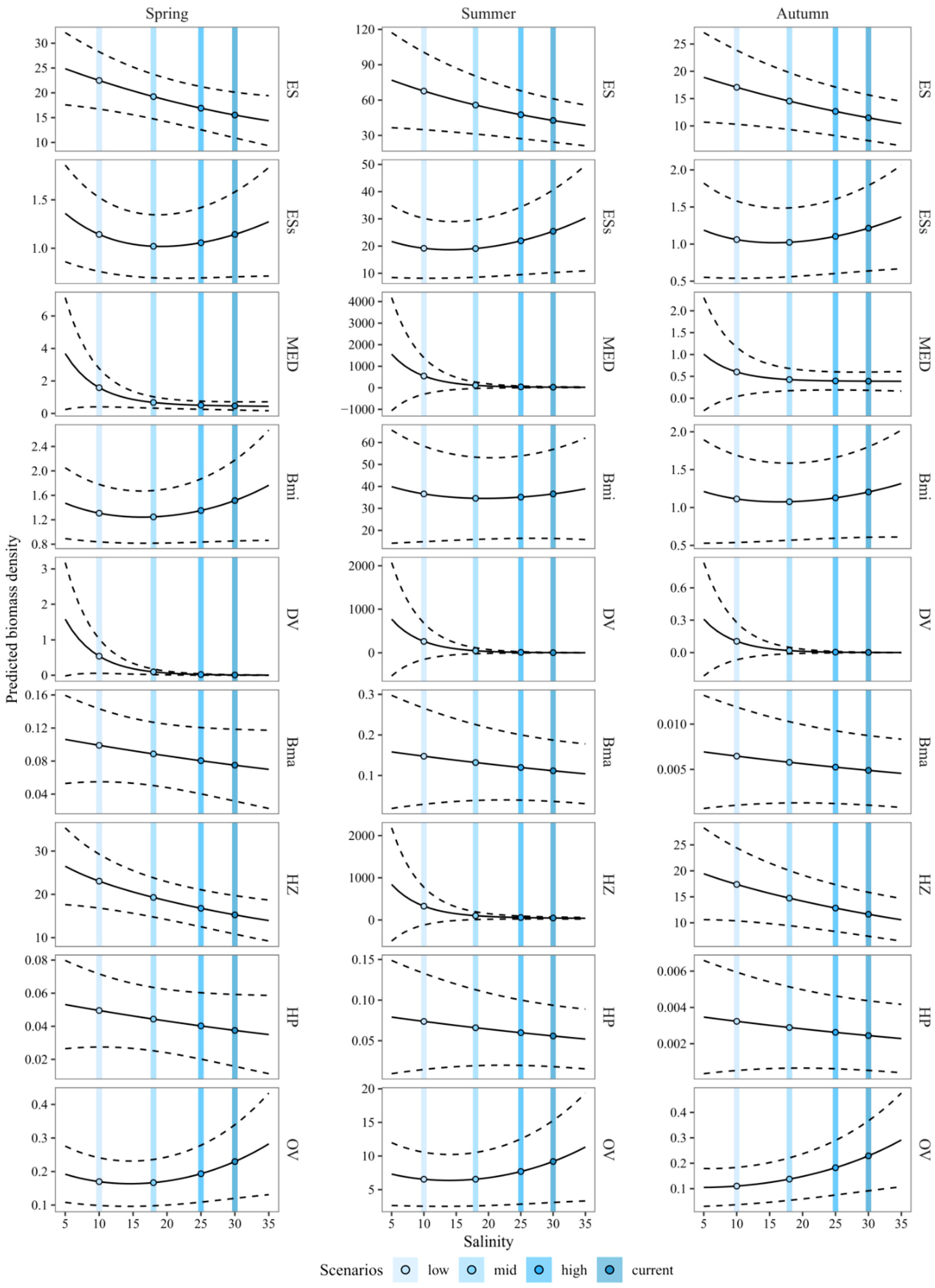
3.3. Predicting the Expected Assemblages
4. Discussion
4.1. Predicting the Expected Nekton Changes after Salinity Reduction
4.2. Implications for Management and Conservation
Author Contributions
Funding
Conflicts of Interest
Appendix A

Appendix B


References
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem: Ecology, Threats and Management, 3rd ed.; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Sheaves, M. Consequences of ecological connectivity: The coastal ecosystem mosaic. Mar. Ecol. Prog. Ser. 2009, 391, 107–115. [Google Scholar] [CrossRef]
- Elliott, M.; Hemingway, K.L. Fishes in Estuaries; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- Kneib, R.T. Salt marsh ecoscapes and production transfers by estuarine nekton in the Southeastern United States. In Concepts and Controversies in Tidal Marsh Ecology; Weinstein, M.P., Kreeger, D.A., Eds.; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M. Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth 2011, 36, 160–166. [Google Scholar] [CrossRef]
- Smyth, K.; Elliott, M. Effects of changing salinity on the ecology of the marine environment. In Stressors in the Marine Environment; Solan, M., Witheley, N., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 161–174. [Google Scholar]
- Rodríguez-Climent, S.; Caiola, N.; Ibáñez, C. Salinity as the main factor structuring small-bodied fish assemblages in hydrologically altered Mediterranean coastal lagoons. Sci. Mar. 2013, 77, 37–45. [Google Scholar] [Green Version]
- Prado, P.; Vergara, C.; Caiola, N.; Ibáñez, C. Influence of salinity regime on the food-web structure and feeding ecology of fish species from Mediterranean coastal lagoons. Estuar. Coast. Shelf Sci. 2014, 139, 1–10. [Google Scholar] [CrossRef]
- Martino, E.J.; Able, K.W. Fish assemblages across the marine to low salinity transition zone of a temperate estuary. Estuar. Coast. Shelf Sci. 2003, 56, 969–987. [Google Scholar] [CrossRef]
- Harrison, T.D.; Whitfield, A.K. Temperature and salinity as primary determinants influencing the biogeography of fishes in South African estuaries. Estuar. Coast. Shelf Sci. 2006, 66, 335–345. [Google Scholar] [CrossRef]
- Franco, A.; Franzoi, P.; Torricelli, P. Structure and functioning of Mediterranean lagoon fish assemblages: A key for the identification of water body types. Estuar. Coast. Shelf Sci. 2008, 79, 549–558. [Google Scholar] [CrossRef]
- Marchand, J.; Codling, I.; Drake, P.; Elliott, M.; Pihl, L.; Rebelo, J. Environmental quality of estuaries. In Fishes in Estuaries; Elliott, M., Hemingway, K.L., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2002; pp. 322–409. [Google Scholar]
- Vasconcelos, R.P.; Reis-Santos, P.; Fonseca, V.; Maia, A.; Ruano, M.; França, S.; Vinagre, C.; Costa, M.J.; Cabral, H. Assessing anthropogenic pressures on estuarine fish nurseries along the Portuguese coast: A multi-metric index and conceptual approach. Sci. Total Environ. 2007, 374, 199–215. [Google Scholar] [CrossRef]
- Perring, M.P.; Standish, R.J.; Price, J.N.; Craig, M.D.; Erickson, T.E.; Ruthrof, K.X.; Whiteley, A.S.; Valentine, L.E.; Hobbs, R.J. Advances in restoration ecology: Rising to the challenges of the coming decades. Ecosphere 2015, 6, 1–25. [Google Scholar] [CrossRef]
- Teichert, N.; Borja, A.; Chust, G.; Uriarte, A.; Lepage, M. Restoring fish ecological quality in estuaries: Implication of interactive and cumulative effects among anthropogenic stressors. Sci. Total Environ. 2016, 542, 383–393. [Google Scholar] [CrossRef]
- Borja, A.; Dauer, D.M.; Elliott, M.; Simenstad, C.A. Medium-and long-term recovery of estuarine and coastal ecosystems: Patterns, rates and restoration effectiveness. Estuar. Coasts 2010, 33, 1249–1260. [Google Scholar] [CrossRef]
- Van Katwijk, M.M.; Thorhaug, A.; Marbà, N.; Orth, R.J.; Duarte, C.M.; Kendrick, G.A.; Althuizen, I.H.J.; Balestri, E.; Bernard, G.; Cambridge, M.L.; et al. Global analysis of seagrass restoration: The importance of large-scale planting. J. Appl. Ecol. 2015, 53, 567–578. [Google Scholar] [CrossRef]
- Society for Ecological Restoration. Tidal Marsh Restoration. A Synthesis of Science and Management; Roman, C.T., Burdick, D.M., Eds.; Island Press: Washington, DC, USA, 2012; ISBN 9781597265768. [Google Scholar]
- Farrugia, T.J.; Espinoza, M.; Lowe, C.G. The fish community of a newly restored southern California estuary: Ecological perspective 3 years after restoration. Environ. Biol. Fishes 2014, 97, 1129–1147. [Google Scholar] [CrossRef]
- Castro, N.; Félix, P.M.; Neto, J.; Cabral, H.; Marques, J.C.; Costa, M.J.; Costa, J. Fish communities’ response to implementation of restoring measures in a highly artificialized estuary. Ecol. Indic. 2016, 67, 743–752. [Google Scholar] [CrossRef]
- Reese, M.M.; Stunz, G.W.; Bushon, A.M. Recruitment of estuarine-dependent nekton through a new tidal inlet: The opening of Packery Channel in Corpus Christi, TX, USA. Estuar. Coasts 2008, 31, 1143–1157. [Google Scholar] [CrossRef]
- Facca, C.; Bonometto, A.; Boscolo, R.; Buosi, A.; Parravicini, M.; Siega, A.; Volpe, V.; Sfriso, A. Coastal lagoon recovery by seagrass restoration. A new strategic approach to meet HD & WFD objectives. In Proceedings of the 9th European Conference on Ecological Restoration, Oulu, Finland, 3–8 August 2014. [Google Scholar]
- Simenstad, C.; Reed, D.; Ford, M. When is restoration not? Ecol. Eng. 2006, 26, 27–39. [Google Scholar] [CrossRef]
- Elliott, M.; Burdon, D.; Hemingway, K.L.; Apitz, S.E. Estuarine, coastal and marine ecosystem restoration: Confusing management and science—A revision of concepts. Estuar. Coast. Shelf Sci. 2007, 74, 349–366. [Google Scholar] [CrossRef]
- Saunders, K.M.; Mcminn, A.; Roberts, D.; Hodgson, D.A.; Heijnis, H. Recent human-induced salinity changes in Ramsar-listed Orielton Lagoon, south-east Tasmania, Australia: A new approach for coastal lagoon conservation and management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2007, 17, 51–70. [Google Scholar] [CrossRef]
- Mohapatra, A.; Mohanty, R.K.; Mohanty, S.K.; Bhatta, K.S.; Das, N.R. Fisheries enhancement and biodiversity assessment of fish, prawn and mud crab in Chilika lagoon through hydrological intervention. Wetl. Ecol. Manag. 2007, 15, 229–251. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rivera-Monroy, V.H.; Chen, R.; Botero, L. Adapting an ecological mangrove model to simulate trajectories in restoration ecology. Mar. Pollut. Bull. 1998, 37, 404–419. [Google Scholar] [CrossRef]
- Das, A.; Justic, D.; Inoue, M.; Hoda, A.; Huang, H.; Park, D. Impacts of Mississippi River diversions on salinity gradients in a deltaic Louisiana estuary: Ecological and management implications. Estuar. Coast. Shelf Sci. 2012, 111, 17–26. [Google Scholar] [CrossRef]
- Koutrakis, E.T.; Sylaios, G.; Kamidis, N.; Markou, D.; Sapounidis, A. Fish fauna recovery in a newly re-flooded Mediterranean coastal lagoon. Estuar. Coast. Shelf Sci. 2009, 83, 505–515. [Google Scholar] [CrossRef]
- Zedler, J.B.; Callaway, J.C. Evaluating the progress of engineered tidal wetlands. Ecol. Eng. 2000, 15, 211–225. [Google Scholar] [CrossRef]
- McAlpine, C.; Catterall, C.P.; Mac Nally, R.; Lindenmayer, D.; Reid, J.L.; Holl, K.D.; Bennett, A.F.; Runting, R.K.; Wilson, K.; Hobbs, R.J.; et al. Integrating plant- and animal- based perspectives for more effective restoration of biodiversity. Front. Ecol. Environ. 2016, 14, 37–45. [Google Scholar] [CrossRef]
- Bourque, A.S.; Fourqurean, J.W. Effects of common seagrass restoration methods on ecosystem structure in subtropical seagrass meadows. Mar. Environ. Res. 2014, 97, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Dolbeth, M.; Cardoso, P.; Grilo, T.; Raffaelli, D.; Pardal, M.A. Drivers of estuarine benthic species distribution patterns following a restoration of a seagrass bed: A functional trait analyses. Mar. Pollut. Bull. 2013, 72, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.H.; Harrower, W.L.; Garris, H.W.; Davidson, S.; Hebert, P.D.N.; Howie, R.; Moody, A.; Polster, D.; Schmitz, O.J.; Sinclair, A.R.E.; et al. A call for applying trophic structure in ecological restoration. Restor. Ecol. 2015, 23, 503–507. [Google Scholar] [CrossRef]
- Brudvig, L.A. Toward prediction in the restoration of biodiversity. J. Appl. Ecol. 2017, 54, 1013–1017. [Google Scholar] [CrossRef] [Green Version]
- Scapin, L.; Zucchetta, M.; Sfriso, A.; Franzoi, P. Predicting the response of nekton assemblages to seagrass transplantations in the Venice lagoon: An approach to assess ecological restoration. Aquat. Conserv. Mar. Freshw. Ecosyst. Mar. Freshw. Ecosyst. 2019, 26, 849–864. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Barak, R.S.; Bauer, J.T.; Caughlin, T.T.; Laughlin, D.C.; Larios, L.; Matthews, J.W.; Stuble, K.L.; Turley, N.E.; Zirbel, C.R. Interpreting variation to advance predictive restoration science. J. Appl. Ecol. 2017, 54, 1018–1027. [Google Scholar] [CrossRef] [Green Version]
- Ebberts, B.D.; Zelinsky, B.D.; Karnezis, J.P.; Studebaker, C.A.; Lopez-Johnston, S.; Creason, A.M.; Krasnow, L.; Johnson, G.E.; Thom, R.M. Estuary ecosystem restoration: Implementing and institutionalizing adaptive management. Restor. Ecol. 2017, 26, 360–369. [Google Scholar] [CrossRef]
- Duarte, C.M.; Borja, A.; Carstensen, J.; Elliott, M.; Krause-Jensen, D.; Marbà, N. Paradigms in the recovery of estuarine and coastal ecosystems. Estuar. Coasts 2015, 38, 1202–1212. [Google Scholar] [CrossRef]
- Elliott, M.; Quintino, V. The Estuarine Quality Paradox, Environmental Homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Zucchetta, M.; Scapin, L.; Cavraro, F.; Pranovi, F.; Franco, A.; Franzoi, P. Can the effects of anthropogenic pressures and environmental variability on nekton fauna be detected in fishery data? Insights from the monitoring of the artisanal fishery within the Venice lagoon. Estuar. Coasts 2016, 39, 1164–1182. [Google Scholar] [CrossRef]
- Pasquaud, S.; Courrat, A.; Fonseca, V.F.; Gamito, R.; Gonçalves, C.I.; Lobry, J.; Lepage, M.; Costa, M.J.; Cabral, H. Strength and time lag of relationships between human pressures and fish-based metrics used to assess ecological quality of estuarine systems. Estuar. Coast. Shelf Sci. 2013, 134, 119–127. [Google Scholar] [CrossRef]
- Fonseca, V.F.; Vasconcelos, R.P.; Gamito, R.; Pasquaud, S.; Gonçalves, C.I.; Costa, J.L.; Costa, M.J.; Cabral, H. Fish community-based measures of estuarine ecological quality and pressure–impact relationships. Estuar. Coast. Shelf Sci. 2013, 134, 128–137. [Google Scholar] [CrossRef]
- Franco, A.; Torricelli, P.; Franzoi, P. A habitat-specific fish-based approach to assess the ecological status of Mediterranean coastal lagoons. Mar. Pollut. Bull. 2009, 58, 1704–1717. [Google Scholar] [CrossRef] [PubMed]
- Milbrandt, E.C.; Bartleson, R.D.; Coen, L.D.; Rybak, O.; Thompson, M.A.; DeAngelo, J.A.; Stevens, P.W. Local and regional effects of reopening a tidal inlet on estuarine water quality, seagrass habitat, and fish assemblages. Cont. Shelf Res. 2012, 41, 1–16. [Google Scholar] [CrossRef]
- Able, K.W.; Grothues, T.M.; Hagan, S.M.; Kimball, M.E.; Nemerson, D.M.; Taghon, G.L. Long-term response of fishes and other fauna to restoration of former salt hay farms: Multiple measures of restoration success. Rev. Fish Biol. Fish. 2008, 18, 65–97. [Google Scholar] [CrossRef]
- Scapin, L.; Zucchetta, M.; Facca, C.; Sfriso, A.; Franzoi, P. Using fish assemblage to identify success criteria for seagrass habitat restoration. Web Ecol. 2016, 16, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Boys, C.A.; Williams, R.J. Succession of fish and crustacean assemblages following reinstatement of tidal flow in a temperate coastal wetland. Ecol. Eng. 2012, 49, 221–232. [Google Scholar] [CrossRef]
- Carniello, L.; Defina, A.; D’Alpaos, L. Morphological evolution of the Venice lagoon: Evidence from the past and trend for the future. J. Geophys. Res. Earth Surf. 2009, 114, 1–10. [Google Scholar] [CrossRef]
- Sarretta, A.; Pillon, S.; Molinaroli, E.; Guerzoni, S.; Fontolan, G. Sediment budget in the Lagoon of Venice, Italy. Cont. Shelf Res. 2010, 30, 934–949. [Google Scholar] [CrossRef] [Green Version]
- Ibanez, C.; Curco, A.; Day, J.W.; Prat, N. Structure and productivity of microtidal Mediterranean coastal marshes. In Concepts and Controversies in Tidal Marsh Ecology; Weinstein, M.P., Kreeger, D.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 107–136. [Google Scholar]
- Bondesan, M.; Castiglioni, G.B.; Elmi, C.; Gabbbianelli, G.; Marocco, R.; Pirazzoli, P.A.; Tomasin, A. Coastal areas at risk from storm surges and sea-level rise in northeastern Italy. J. Coast. Res. 1995, 11, 1354–1379. [Google Scholar]
- Cencini, C. Physical processes and human activities in the evolution of the Po delta, Italy. J. Coast. Res. 1998, 14, 774–793. [Google Scholar]
- Fontolan, G.; Pillon, S.; Bezzi, A.; Villalta, R.; Lipizer, M.; Triches, A.; D’Aietti, A. Human impact and the historical transformation of saltmarshes in the Marano and Grado Lagoon, northern Adriatic Sea. Estuar. Coast. Shelf Sci. 2012, 113, 41–56. [Google Scholar] [CrossRef]
- Marani, M.; Lanzoni, S.; Silvestri, S.; Rinaldo, A. Tidal landforms, patterns of halophytic vegetation and the fate of the lagoon of Venice. J. Mar. Syst. 2004, 51, 191–210. [Google Scholar] [CrossRef]
- Tambroni, N.; Seminara, G. Are inlets responsible for the morphological degradation of Venice Lagoon? J. Geophys. Res. 2006, 111, F03013. [Google Scholar] [CrossRef]
- Ferrarin, C.; Ghezzo, M.; Umgiesser, G.; Tagliapietra, D.; Camatti, E.; Zaggia, L.; Sarretta, A. Assessing hydrological effects of human interventions on coastal systems: Numerical applications to the Venice Lagoon. Hydrol. Earth Syst. Sci. 2013, 17, 1733–1748. [Google Scholar] [CrossRef]
- Defina, A.; Carniello, L.; Fagherazzi, S.; D’Alpaos, L. Self-organization of shallow basins in tidal flats and salt marshes. J. Geophys. Res. Earth Surf. 2007, 112, 1–11. [Google Scholar] [CrossRef]
- Molinaroli, E.; Guerzoni, S.; Sarretta, A.; Masiol, M.; Pistolato, M. Thirty-year changes (1970 to 2000) in bathymetry and sediment texture recorded in the Lagoon of Venice sub-basins, Italy. Mar. Geol. 2009, 258, 115–125. [Google Scholar] [CrossRef] [Green Version]
- D’Alpaos, L. Fatti e Misfatti di Idraulica Lagunare. La Laguna di Venezia dalla Diversione dei Fiumi alle Nuove Opere delle Bocche di Porto; Istituto Veneto di Scienze Lettere ed Arti.: Venice, Italy, 2010. [Google Scholar]
- Feola, A.; Bonometto, A.; Ponis, E.; Cacciatore, F.; Oselladore, F.; Matticchio, B.; Canesso, D.; Sponga, S.; Volpe, V.; Lizier, M.; et al. LIFE LAGOON REFRESH. Ecological restoration in Venice Lagoon (Italy): Concrete actions supported by numerical modeling and stakeholder involvement. In Proceedings of the Citizen Observatories for natural hazards and Water Management—2nd International Conference, Venice, Italy, 27–30 November 2018; Available online: http://www.lifelagoonrefresh.eu/file/pubblicazioni/COWM_2018_Extendedabstract.pdf (accessed on 26 April 2019).
- Umgiesser, G.; Melaku canu, D.; Cucco, A.; Solidoro, C. A finite element model for the Venice Lagoon. Development, set up, calibration and validation. J. Mar. Syst. 2004, 51, 123–145. [Google Scholar] [CrossRef]
- Solidoro, C.; Melaku Canu, D.; Cucco, A.; Umgiesser, G. A partition of the Venice Lagoon based on physical properties and analysis of general circulation. J. Mar. Syst. 2004, 51, 147–160. [Google Scholar] [CrossRef]
- Molinaroli, E.; Guerzoni, S.; Sarretta, A.; Cucco, A.; Umgiesser, G. Links between hydrology and sedimentology in the Lagoon of Venice, Italy. J. Mar. Syst. 2007, 68, 303–317. [Google Scholar] [CrossRef] [Green Version]
- Ghezzo, M.; Guerzoni, S.; Cucco, A.; Umgiesser, G. Changes in Venice Lagoon dynamics due to construction of mobile barriers. Coast. Eng. 2010, 57, 694–708. [Google Scholar] [CrossRef] [Green Version]
- Ghezzo, M.; Sarretta, A.; Sigovini, M.; Guerzoni, S.; Tagliapietra, D.; Umgiesser, G. Modeling the inter-annual variability of salinity in the lagoon of Venice in relation to the water framework directive typologies. Ocean Coast. Manag. 2011, 54, 706–719. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Franzoi, P.; Malavasi, S.; Riccato, F.; Torricelli, P.; Mainardi, D. Use of shallow water habitats by fish assemblages in a Mediterranean coastal lagoon. Estuar. Coast. Shelf Sci. 2006, 66, 67–83. [Google Scholar] [CrossRef]
- Malavasi, S.; Franco, A.; Fiorin, R.; Franzoi, P.; Torricelli, P.; Mainardi, D. The shallow water gobiid assemblage of the Venice Lagoon: Abundance, seasonal variation and habitat partitioning. J. Fish Biol. 2005, 67, 146–165. [Google Scholar] [CrossRef]
- Franzoi, P.; Franco, A.; Torricelli, P. Fish assemblage diversity and dynamics in the Venice lagoon. Rendiconti Lincei 2010, 21, 269–281. [Google Scholar] [CrossRef]
- Scapin, L.; Zucchetta, M.; Sfriso, A.; Franzoi, P. Local habitat and seascape structure influence seagrass fish assemblages in the Venice lagoon: The value of conservation at multiple spatial scales. Estuar. Coasts 2018, 41, 2410–2425. [Google Scholar] [CrossRef]
- ARPAV. Piano di Monitoraggio dei Corpi Idrici della Laguna di Venezia Finalizzato alla Definizione dello Stato Ecologico, ai Sensi della Direttiva 2000/60/CE; Relazione Finale; Agenzia Regionale per la Prevenzione e Protezione Ambientale del Veneto: Venezia, Italy, 2012. [Google Scholar]
- Magistrato alle Acque di Venezia (ora Provveditorato Interregionale alle OO. PP. del Veneto—Trentino Alto Adige—Friuli Venezia Giulia)—Selc Studio B.12.3/III. La funzionalità dell’ambiente lagunare attraverso rilievi delle risorse alieutiche, dell’avifauna e dell’ittiofauna. Erodibilità del fondale e fattori di disturbo: Rilievi dell’erodibilità del fondale; Rapporto Intermedio; 2005. [Google Scholar]
- Magistrato alle Acque di Venezia (ora Provveditorato Interregionale alle OO. PP. del Veneto—Trentino Alto Adige—Friuli Venezia Giulia)—Thetis. Programma Generale delle Attività di Approfondimento del Quadro Conoscitivo di Riferimento per Gli Interventi Ambientali. 2° Stralcio Triennale (2003–2006) “Progetto ICSEL”; Attività A; Prodotto dal Concessionario, Consorzio Venezia Nuova: Venice, Italy, 2005. [Google Scholar]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. Mvabund—An R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning, 2nd ed.; Springer: New York, NY, USA, 2009; ISBN 978-0-387-84857-0. [Google Scholar]
- Potter, I.C.; Tweedley, J.R.; Elliott, M.; Whitfield, A.K. The ways in which fish use estuaries: A refinement and expansion of the guild approach. Fish Fish. 2013, 16, 230–239. [Google Scholar] [CrossRef]
- Franco, A.; Elliott, M.; Franzoi, P.; Torricelli, P. Life strategies of fishes in European estuaries: The functional guild approach. Mar. Ecol. Progr. Ser. 2008, 354, 219–228. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. Available online: http://www.fishbase.org (accessed on 29 June 2019).
- Catalano, B.; Penna, M.; Riccato, F.; Fiorin, R.; Franceschini, G.; Antonini, C.; Zucchetta, M.; Cicero, A.M.; Franzoi, P. Manuale per la classificazione dell’Elemento di Qualità Biologica “Fauna Ittica” nelle lagune costiere italiane. Applicazione dell’indice nazionale HFBI (Habitat Fish Bio-Indicator) ai sensi del D.Lgs 152/2006; ISPRA: Roma, Italy, 2017; ISBN 9788844808716. [Google Scholar]
- Franco, A.; Franzoi, P.; Malavasi, S.; Zucchetta, M.; Torricelli, P. Population and habitat status of two endemic sand gobies in lagoon marshes—Implications for conservation. Estuar. Coast. Shelf Sci. 2012, 114, 31–40. [Google Scholar] [CrossRef]
- Rountree, R.A.; Able, K.W. Spatial and temporal habitat use patterns for salt marsh nekton: Implications for ecological functions. Aquat. Ecol. 2007, 41, 25–45. [Google Scholar] [CrossRef]
- Poizat, G.; Rosecchi, E.; Chauvelon, P.; Contournet, P.; Crivelli, A.J. Long-term fish and macro-crustacean community variation in a Mediterranean lagoon. Estuar. Coast. Shelf Sci. 2004, 59, 615–624. [Google Scholar] [CrossRef]
- Milardi, M.; Gavioli, A.; Lanzoni, M.; Fano, E.A.; Castaldelli, G. Meteorological factors influence marine and resident fish movements in a brackish lagoon. Aquat. Ecol. 2019, 53, 251–263. [Google Scholar] [CrossRef]
- Franco, A.; Riccato, F.; Torricelli, P.; Franzoi, P. Fish assemblage response to environmental pressures in the Venice lagoon. Trans. Waters Bull. 2009, 3, 29–44. [Google Scholar]
- Franco, A.; Malavasi, S.; Zucchetta, M.; Torricelli, P.; Franzoi, P. Environmental influences on fish assemblage in the Venice Lagoon, Italy. Chem. Ecol. 2006, 22, S105–S118. [Google Scholar] [CrossRef]
- Cavraro, F.; Varin, C.; Malavasi, S. Lunar-induced reproductive patterns in transitional habitats: Insights from a Mediterranean killifish inhabiting northern Adriatic Saltmarshes. Estuar. Coast. Shelf Sci. 2014, 139, 60–66. [Google Scholar] [CrossRef]
- Zucchetta, M.; Franco, A.; Torricelli, P.; Franzoi, P. Habitat distribution model for European flounder juveniles in the Venice lagoon. J. Sea Res. 2010, 64, 133–144. [Google Scholar] [CrossRef]
- Verdiell-Cubedo, D.; Oliva-Paterna, F.J.; Ruiz-Navarro, A.; Torralva, M. Assessing the nursery role for marine fish species in a hypersaline coastal lagoon (Mar Menor, Mediterranean Sea). Mar. Biol. Res. 2013, 9, 739–748. [Google Scholar] [CrossRef]
- Tournois, J.; Darnaude, A.M.; Ferraton, F.; Aliaume, C.; Mercier, L.; McKenzie, D.J. Lagoon nurseries make a major contribution to adult populations of a highly prized coastal fish. Limnol. Oceanogr. 2017, 62, 1219–1233. [Google Scholar] [CrossRef]
- Cavraro, F.; Zucchetta, M.; Malavasi, S.; Franzoi, P. Small creeks in a big lagoon: The importance of marginal habitats for fish populations. Ecol. Eng. 2017, 99, 228–237. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ceoldo, S.; Marcomini, A. Recording the occurrence of trophic level changes in the lagoon of Venice over the ’90s. Environ. Int. 2005, 31, 993–1001. [Google Scholar] [PubMed]
- Sfriso, A.; Facca, C. Distribution and production of macrophytes and phytoplankton in the lagoon of Venice: Comparison of actual and past situation. Hydrobiologia 2007, 577, 71–85. [Google Scholar] [CrossRef]
- Whitfield, A.K.; Elliott, M. Fishes as indicators of environmental and ecological changes within estuaries: A review of progress and some suggestions. J. Fish Biol. 2002, 61, 229–250. [Google Scholar] [CrossRef]
- Cabral, H.; Fonseca, V.F.; Gamito, R.; Gonçalves, C.I.; Costa, J.L.; Erzini, K.; Gonçalves, J.; Martins, J.; Leite, L.; Andrade, J.P.; et al. Ecological quality assessment of transitional waters based on fish assemblages in Portuguese estuaries: The Estuarine Fish Assessment Index (EFAI). Ecol. Indic. 2012, 19, 144–153. [Google Scholar] [CrossRef]
- Cavraro, F.; Bettoso, N.; Zucchetta, M.; D’Aietti, A.; Faresi, L.; Franzoi, P. Body condition in fish as a tool to detect the effects of anthropogenic pressures in transitional waters. Aquat. Ecol. 2019, 53, 21–35. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Franco, T.P.; Neves, L.M.; Araújo, F.G. Better with more or less salt? The association of fish assemblages in coastal lagoons with different salinity ranges. Hydrobiologia 2019, 828, 83–100. [Google Scholar] [CrossRef]
- Manzo, C.; Fabbrocini, A.; Roselli, L.; D’Adamo, R. Characterization of the fish assemblage in a Mediterranean coastal lagoon: Lesina Lagoon (central Adriatic Sea). Reg. Stud. Mar. Sci. 2016, 8, 192–200. [Google Scholar] [CrossRef]
- Pérez-Domínguez, R.; Maci, S.; Courrat, A.; Lepage, M.; Borja, A.; Uriarte, A.; Neto, J.M.; Cabral, H.; St. Raykov, V.; Franco, A.; et al. Current developments on fish-based indices to assess ecological-quality status of estuaries and lagoons. Ecol. Indic. 2012, 23, 34–45. [Google Scholar]
- Alvarez, M.C.; Franco, A.; Pérez-Domínguez, R.; Elliott, M. Sensitivity analysis to explore responsiveness and dynamic range of multi-metric fish-based indices for assessing the ecological status of estuaries and lagoons. Hydrobiologia 2013, 704, 347–362. [Google Scholar] [CrossRef]
- Reyjol, Y.; Argillier, C.; Bonne, W.; Borja, A.; Buijse, A.D.; Cardoso, A.C.; Daufresne, M.; Kernan, M.; Ferreira, M.T.; Poikane, S.; et al. Assessing the ecological status in the context of the European Water Framework Directive: Where do we go now? Sci. Total Environ. 2014, 497–498, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Lepage, M.; Harrison, T.; Breine, J.; Cabral, H.; Coates, S.; Galván, C.; García, P.; Jager, Z.; Kelly, F.; Mosch, E.C.; et al. An approach to intercalibrate ecological classification tools using fish in transitional water of the North East Atlantic. Ecol. Indic. 2016, 67, 318–327. [Google Scholar] [CrossRef]
- Franco, A.; Fiorin, R.; Franzoi, P.; Torricelli, P. Threatened fishes of the world: Pomatoschistus canestrinii Ninni, 1883 (Gobiidae). Environ. Biol. Fishes 2005, 72, 32. [Google Scholar] [CrossRef]
- Franco, A.; Fiorin, R.; Zucchetta, M.; Torricelli, P.; Franzoi, P. Flounder growth and production as indicators of the nursery value of marsh habitats in a Mediterranean lagoon. J. Sea Res. 2010, 64, 457–464. [Google Scholar] [CrossRef]
- Pranovi, F.; Caccin, A.; Franzoi, P.; Malavasi, S.; Zucchetta, M.; Torricelli, P. Vulnerability of artisanal fisheries to climate change in the Venice Lagoon. J. Fish Biol. 2013, 83, 847–864. [Google Scholar]
- Pérez-Ruzafa, A.; Mompeán, M.C.; Marcos, C. Hydrographic, geomorphologic and fish assemblage relationships in coastal lagoons. Hydrobiologia 2007, 577, 107–125. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C. Fisheries in coastal lagoons: An assumed but poorly researched aspect of the ecology and functioning of coastal lagoons. Estuar. Coast. Shelf Sci. 2012, 110, 15–31. [Google Scholar] [CrossRef]
- Management of Coastal Lagoon Fisheries; Kapetsky, J.M.; Lasserre, G. (Eds.) FAO Studies and Reviews, GFCM 61; FAO: Rome, Italy, 1984.
- Ardizzone, G.D.; Cataudella, S.; Rossi, R. Management of coastal lagoon fisheries and aquaculture in Italy. AO Fish. Tech. Paper 1988, 293, 1–103. [Google Scholar]
- Nagelkerken, I.; Sheaves, M.; Baker, R.; Connolly, R.M. The seascape nursery: A novel spatial approach to identify and manage nurseries for coastal marine fauna. Fish Fish. 2015, 16, 362–371. [Google Scholar] [CrossRef]
- Brown, C.J.; Harborne, A.R.; Paris, C.B.; Mumby, P.J. Uniting paradigms of connectivity in marine ecology. Ecology 2016, 97, 2447–2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, A.K.; Pattrick, P. Habitat type and nursery function for coastal marine fish species, withemphasis on the Eastern Cape region, South Africa. Estuar. Coast. Shelf Sci. 2015, 160, 49–59. [Google Scholar] [CrossRef]
- Day, J.W.; Rybczyk, J.; Scarton, F.; Rismondo, A.; Are, D.; Cecconi, G. Soil accretionary dynamics, sea-level rise and the survival of wetlands in Venice lagoon: A field and modelling approach. Estuar. Coast. Shelf Sci. 1999, 49, 607–628. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Bellafiore, D.; Ghezzo, M.; Tagliapietra, D.; Umgiesser, G. Climate change and artificial barrier effects on the Venice Lagoon: Inundation dynamics of salt marshes and implications for halophytes distribution. Ocean Coast. Manag. 2014, 100, 101–115. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Category | Formula | Predictors Included |
| m0 | Yi ~ intercept + εi | None |
| m1 | m0 + season + year | Temporal |
| m2 | m1 + sub-basin | Temporal + geographical |
| m3.1 | m2 + temperature | Temporal + geographical + environmental |
| m3.2 | m2 + temperature + salinity | |
| m3.3 | m2 + temperature + salinity + DO | |
| m3.4 | m2 + temperature + salinity + DO + turbidity | |
| m3.5 | m2 + temperature + salinity + DO + turbidity + grain | |
| m3.6 | m2 + temperature + salinity + DO + turbidity + grain + algae | |
| m4 | m3.x + location | Temporal + geographical + environmental + location (marsh creek or edge) |
| Test | Testing the Effect of: | |
| t1 | m0 vs. m1 | Temporal factor |
| t2 | m1 vs. m2 | Geographical factor, when only temporal factor was considered before |
| t3.1 | m2 vs. m3.1 | Each environmental factor, when temporal, geographical and all the significant previous environmental factors were considered before |
| t3.2 | m3.1 vs. m3.2 | |
| t3.3 | m3.2 vs. m3.3 | |
| t3.4 | m3.3 vs. m3.4 | |
| t3.5 | m3.4 vs. m3.5 | |
| t3.6 | m3.5 vs. m3.6 | |
| t4 | m3.x vs. m4 | Location factor, when temporal, geographical and all the significant environmental factors were considered before |
| Scenario | Salinity (PSU) | Season | Year | Sub-Basin | Other Environmental Variables | Location |
|---|---|---|---|---|---|---|
| Low | 10 | Either spring, summer or autumn | Average sampling year considered in calibration | Northern | Average values measured in the northern sub-basin during each season | Saltmarsh edge |
| Mid | 18 | |||||
| High | 25 | |||||
| Current | 30 |
| Average Biomass (g 100 m−2) | Spearman’s Coefficient | |||||
|---|---|---|---|---|---|---|
| Group | Species | EUFG | FMFG | Average | Standard Error | |
| Decapods | Palaemon elegans | ES | Bmi, OV | 0.93 | 0.60 | 0.004 |
| Crangon crangon | MED | Bmi | 3.63 | 0.60 | 0.22 | |
| Fish | Atherina boyeri | ES | HZ | 25.59 | 0.49 | 0.11 |
| Pomatoschistus canestrinii | ESs | Bmi, HZ | 1.38 | 0.48 | 0.09 | |
| Chelon ramada | MED | HZ, DV | 22.43 | 0.42 | 0.15 | |
| Platichthys flesus | MED | Bmi, Bma, HP | 0.65 | 0.39 | 0.04 | |
| Pomatoschistus marmoratus | ES | Bmi, HZ | 0.46 | 0.35 | 0.08 | |
| Knipowitschia panizzae | ESs | Bmi, HZ | 2.13 | 0.31 | 0.03 | |
| Aphanius fasciatus | ESs | Bmi, OV | 9.55 | 0.28 | 0.10 | |
| Chelon saliens | MED | HZ, DV | 2.04 | 0.18 | 0.08 | |
| Sparus aurata | MED | Bmi, Bma, HZ | 0.53 | 0.16 | 0.003 | |
| Chelon auratus | MED | HZ, DV | 4.68 | 0.14 | 0.22 | |
| Whole assemblage | 0.46 | 0.05 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scapin, L.; Zucchetta, M.; Bonometto, A.; Feola, A.; Boscolo Brusà, R.; Sfriso, A.; Franzoi, P. Expected Shifts in Nekton Community Following Salinity Reduction: Insights into Restoration and Management of Transitional Water Habitats. Water 2019, 11, 1354. https://doi.org/10.3390/w11071354
Scapin L, Zucchetta M, Bonometto A, Feola A, Boscolo Brusà R, Sfriso A, Franzoi P. Expected Shifts in Nekton Community Following Salinity Reduction: Insights into Restoration and Management of Transitional Water Habitats. Water. 2019; 11(7):1354. https://doi.org/10.3390/w11071354
Chicago/Turabian StyleScapin, Luca, Matteo Zucchetta, Andrea Bonometto, Alessandra Feola, Rossella Boscolo Brusà, Adriano Sfriso, and Piero Franzoi. 2019. "Expected Shifts in Nekton Community Following Salinity Reduction: Insights into Restoration and Management of Transitional Water Habitats" Water 11, no. 7: 1354. https://doi.org/10.3390/w11071354
APA StyleScapin, L., Zucchetta, M., Bonometto, A., Feola, A., Boscolo Brusà, R., Sfriso, A., & Franzoi, P. (2019). Expected Shifts in Nekton Community Following Salinity Reduction: Insights into Restoration and Management of Transitional Water Habitats. Water, 11(7), 1354. https://doi.org/10.3390/w11071354