Removal and Ecotoxicity of 2,4-D and MCPA in Microbial Cultures Enriched with Structurally-Similar Plant Secondary Metabolites


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microcosm Setup
2.2. 2,4-D and MCPA Concentration Measurement and Their Ppercentage Removal
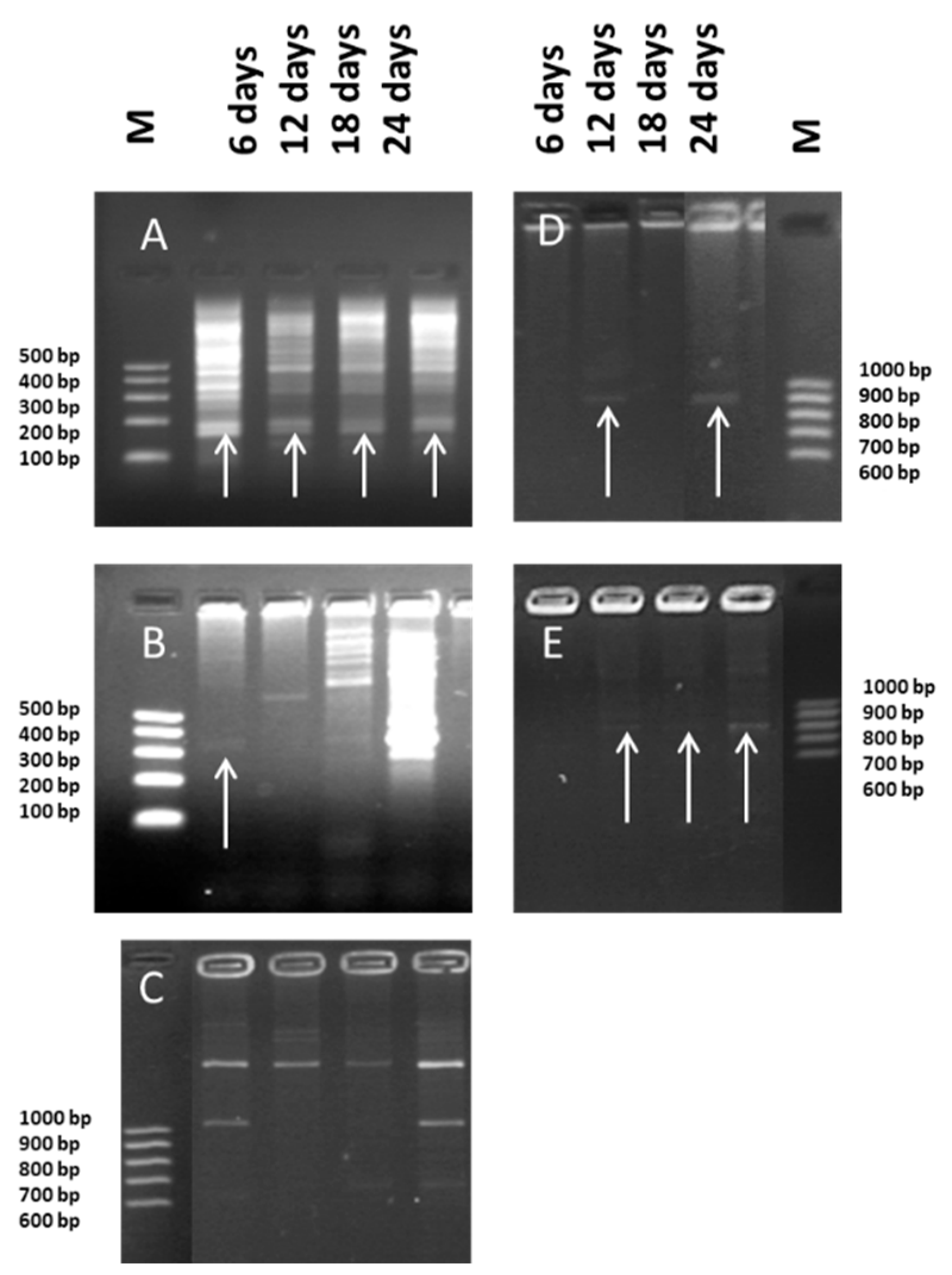
2.3. Molecular Analysis
2.4. Ecotoxicity Assessment
3. Results
3.1. Changes in 2,4-D and MCPA Concentration
3.2. Molecular Analysis
3.3. Changes in Ecotoxicity
4. Discussion
4.1. The Influence of Selected PSMs on 2,4-D/MCPA Removal
4.2. The Influence of Selected PSMs on the Degradation Potential of SE Bacteria toward 2,4-D/MCPA
4.3. The Influence of Selected PSMs on the Ecotoxicity of 2,4-D/MCPA-Enriched Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PSM | plant secondary metabolite |
| FA | ferulic acid |
| SA | syringic acid |
| PH | phenoxy herbicides |
| 2,4-D | 2,4-dichlorophenoxy acid |
| MCPA | 2-methyl-4-chlorophenoxyacetic acid |
| SE | soil extract |
| SSE | sterile soil extract |
| PR | percentage removal |
| PE | percentage effect |
References
- 2008–2012 Market Estimates. Pesticides Industry Sales and Usage; U.S. Environmental Protection Agency: Washington, DC, USA, 2017.
- Smith, A.E.; Mortensen, K.; Aubin, A.J.; Molloy, M.M. Degradation of MCPA, 2,4-D, and Other Phenoxyalkanoic Acid Herbicides Using an Isolated Soil Bacterium. J. Agric. Food Chem. 1994, 42, 401–405. [Google Scholar] [CrossRef]
- Michael, A. Kamrin Phenoxy and Benzoic Acid Herbicides. In Pesticide Profiles, 1st ed.; CRC Press: Boca Raton, FL, USA, 1997; ISBN 978-1-56670-190-7. [Google Scholar]
- Skiba, E.; Wolf, W.M. Commercial Phenoxyacetic Herbicides Control Heavy Metal Uptake by wheat in a divergent way than pure active substances alone. Environ. Sci. Eur. 2017, 29, 26. [Google Scholar] [CrossRef] [PubMed]
- Eurostat Sales of pesticides by type of pesticide. Available online: https://ec.europa.eu/eurostat/web/products-datasets/product?code=tai02 (accessed on 11 March 2019).
- Ignatowicz, K.; Struk-Sokołowska, J. Sezonowe wahania zanieczyszczeń agrotechnicznych w rzece Narwi ze szczególnym uwzględnieniem herbicydów fenoksyoctowych. Środkowo-Pomorskie Tow. Nauk. Ochr. Środowiska. 2004, 4, 189–205. [Google Scholar]
- Ismail, B.S.; Prayitno, S.; Tayeb, M.A. Contamination of rice field water with sulfonylurea and phenoxy herbicides in the Muda Irrigation Scheme, Kedah, Malaysia. Environ. Monit. Assess. 2015, 187, 406. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wu, J.Q.; Jiang, L.L.; Shen, L.Z.; Li, J.Y.; He, Z.H.; Wei, P.; Lv, Z.; He, M.F. Developmental toxicity of 2,4-dichlorophenoxyacetic acid in zebrafish embryos. Chemosphere. 2017, 171, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Caux, P.-Y.; Ménard, L.; Kent, R.A. Comparative study of the effects of MCPA, butylate, atrazine, and cyanazine on Selenastrum capricornutum. Environ. Pollut. 1996, 92, 219–225. [Google Scholar] [CrossRef]
- Batogliu-Pazarbasi, M.; Milosevic, N.; Malaguerra, F.; Binning, P.J.; Albrechtsen, H.J.; Bjerg, P.L.; Aamand, J. Discharge of landfill leachate to streambed sediments impacts the mineralization potential of phenoxy acid herbicides depending on the initial abundance of tfdA gene classes. Environ. Pollut. 2013, 176, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Mcgowan, C.; Fulthorpe, R.; Wright, A.; Tiedje, J.M.; Gowan, C.M.C. Evidence for Interspecies Gene Transfer in the Evolution of Evidence for Interspecies Gene Transfer in the Evolution of 2,4-Dichlorophenoxyacetic Acid Degraders. Appl. Environ. Microbiol. 1998, 64, 4089–4092. [Google Scholar]
- Kitagawa, W.; Kamagata, Y. Diversity of 2,4-Dichlorophenoxyacetic Acid (2,4-D)-Degradative Genes and Degrading Bacteria. In Biodegradative Bacteria: How Bacteria Degrade, Survive, Adapt, and Evolve; Nojiri, H., Fukuda, M., Tsuda, M., Kamagata, Y., Eds.; Springer: Japan, 2014; pp. 43–57. ISBN 978-4-431-54519-4. [Google Scholar]
- Musilova, L.; Ridl, J.; Polivkova, M.; Macek, T.; Uhlik, O. Effects of Secondary Plant Metabolites on Microbial Populations: Changes in Community Structure and Metabolic Activity in Contaminated Environments. Int. J. Mol. Sci. 2016, 17, 1205. [Google Scholar] [CrossRef]
- Hu, C.; Zhang, Y.; Tang, X.; Luo, W. PCB Biodegradation and bphA1 Gene Expression Induced by Salicylic Acid and Biphenyl with Pseudomonas fluorescence P2W and Ralstonia eutropha H850. Pol. J. Environ. Stud. 2014, 23, 1591–1598. [Google Scholar]
- Uhlik, O.; Musilova, L.; Ridl, J.; Hroudova, M.; Vlcek, C.; Koubek, J.; Holeckova, M.; Mackova, M.; Macek, T. Plant secondary metabolite-induced shifts in bacterial community structure and degradative ability in contaminated soil. Appl. Microbiol. Biotechnol. 2013, 97, 9245–9256. [Google Scholar] [CrossRef] [PubMed]
- Kruczek, M. Pumpkin (Cucurbita sp.) as a source of health-beneficial compounds with antioxidant properties Dynia (Cucurbita sp.) jako źródło prozdrowotnych związków o charakterze antyoksydacyjnym. Przem. Chem. 2015, 1, 86–90. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Chung, I.-M.; Thiruvengadam, M. Evaluation of phenolic compounds, antioxidant and antimicrobial activities from transgenic hairy root cultures of gherkin (Cucumis anguria L.). South. African, J. Bot. 2015, 100, 80–86. [Google Scholar] [CrossRef]
- Suttinun, O.; Lederman, P.B.; Luepromchai, E. Application of terpene-induced cell for enhancing biodegradation of TCE contaminated soil. Songklanakarin, J. Sci. Technol. 2004, 26, 131–142. [Google Scholar]
- White, J.C.; Mattina, M.I.; Lee, W.Y.; Eitzer, B.D.; Iannucci-Berger, W. Role of organic acids in enhancing the desorption and uptake of weathered p,p-DDE by Cucurbita pepo. Environ. Pollut. 2003, 124, 71–80. [Google Scholar] [CrossRef]
- Yi, H.; Crowley, D.E. Biostimulation of PAH degradation with plants containing high concentrations of linoleic acid. Environ. Sci. Technol. 2007, 41, 4382–4388. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, E.; Rhodes, A.H.; Owen, S.M.; Semple, K.T. Biogenic volatile organic compounds as a potential stimulator for organic contaminant degradation by soil microorganisms. Environ. Pollut. 2009, 157, 86–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Wu, F. Effects of amendments of ferulic acid on soil microbial communities in the rhizosphere of cucumber (Cucumis sativus L.). Eur. J. Soil Biol. 2012, 50, 191–197. [Google Scholar] [CrossRef]
- Modern Water RaPID Assay ® 2,4-D. Available online: https://www.modernwater.com/pdf/MW_Factsheet_Rapid-Assay_2-4-D.pdf (accessed on 11 September 2017).
- Orphan, V.J.; Sylva, S.P.; Hayes, J.M.; Delong, E.F. Comparative Analysis of Methane-Oxidizing Archaea and Sulfate-Reducing Bacteria in Anoxic Marine Sediments Comparative Analysis of Methane-Oxidizing Archaea and Sulfate-Reducing Bacteria in Anoxic Marine Sediments. Appl. Environ. Microbiol. 2001, 67, 1922–1934. [Google Scholar] [CrossRef] [PubMed]
- MicroBioTests Inc. Standard Operational Procedure, Phytotoxkit. Seed Germination and Early Growth Microbiotest with Higher Plants. Available online: https://www.microbiotests.com/SOPs/Phytotestkit%20(complete%20test)%20SOP%20-%20A5.pdf (accessed on 11 September 2017).
- Persoone, G.; Marsalek, B.; Blinova, I.; Törökne, A.; Zarina, D.; Manusadzianas, L.; Nalecz-Jawecki, G.; Tofan, L.; Stepanova, N.; Tothova, L.; et al. A practical and user-friendly toxicity classification system with microbiotests for natural waters and wastewaters. Environ. Toxicol. 2003, 18, 395–402. [Google Scholar] [CrossRef]
- Paszko, T.; Muszyński, P.; Materska, M.; Bojanowska, M.; Kostecka, M.; Jackowska, I. Adsorption and degradation of phenoxyalkanoic acid herbicides in soils: A review. Environ. Toxicol. Chem. 2016, 35, 271–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrilescu, M. Fate of Pesticides in the Environment and its Bioremediation. Eng. Life Sci. 2005, 5, 497–526. [Google Scholar] [CrossRef]
- McMartin, D.W.; Gillies, J.A.; Headley, J.V.; Peterson, H.G. Biodegradation Kinetics of 2,4-Dichlorophenoxyacetic Acid (2,4-D) in South Saskatchewan River Water. Can. Water Resour. J. 2000, 25, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Urbaniak, M.; Mierzejewska, E.; Tankiewicz, M. The stimulating role of syringic acid, a plant secondary metabolite, in the microbial degradation of structurally-related herbicide, MCPA. PeerJ 2019, 7, e6745. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Tashiro, Y.; Uobe, K.; Kamagata, Y.; Suyama, K.; Yamamoto, H. Root Nodule Bradyrhizobium spp. Harbor tfdAα and cadA, Homologous with Genes Encoding 2,4-Dichlorophenoxyacetic Acid-Degrading Proteins. Appl. Environ. Microbiol. 2004, 70, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- Poll, C.; Pagel, H.; Devers-Lamrani, M.; Martin-Laurent, F.; Ingwersen, J.; Streck, T.; Kandeler, E. Regulation of bacterial and fungal MCPA degradation at the soil-litter interface. Soil Biol. Biochem. 2010, 42, 1879–1887. [Google Scholar] [CrossRef]
- Kitagawa, W.; Takami, S.; Miyauchi, K.; Masai, E.; Kamagata, Y.; Tiedje, J.M.; Fukuda, M. Novel 2,4-Dichlorophenoxyacetic Acid Degradation Genes from Oligotrophic. Society 2002, 184, 509–518. [Google Scholar]
- Bælum, J.; Jacobsen, C.S.; Holben, W.E. Comparison of 16S rRNA gene phylogeny and functional tfdA gene distribution in thirty-one different 2,4-dichlorophenoxyacetic acid and 4-chloro-2-methylphenoxyacetic acid degraders. Syst. Appl. Microbiol. 2010, 33, 67–70. [Google Scholar] [CrossRef]
- Mierzejewska, E.; Baran, A.; Urbaniak, M. The influence of MCPA on soil phytotoxicity and the presence of genes involved in its biodegradation. Arch. Environ. Prot. 2017, 44, 58–64. [Google Scholar]
- Silva, T.M.; Stets, M.I.; Mazzetto, A.M.; Andrade, F.D.; Pileggi, S.A.V.; Fávero, P.R.; Cantú, M.D.; Carrilho, E.; Carneiro, P.I.B.; Pileggi, M. Degradation of 2,4-D Herbicide by Microorganisms Isolated from Brazilian Contaminated Soil. Braz. J. Microbiol. 2007, 38, 522–525. [Google Scholar] [CrossRef]
- Del Pilar Castillo, M.; Andersson, A.; Ander, P.; Stenström, J.; Torstensson, L. Establishment of the white rot fungus Phanerochaete chrysosporium on unsterile straw in solid substrate fermentation systems intended for degradation of pesticides. World J. Microbiol. Biotechnol. 2001, 17, 627–633. [Google Scholar] [CrossRef]
- Bælum, J.; Henriksen, T.; Christian, H.; Hansen, B.; Jacobsen, C.S. Degradation of 4-Chloro-2-Methylphenoxyacetic Acid in Top- and Subsoil Is Quantitatively Linked to the Class III tfdA Gene. Appl. Environ. Microbiol. 2006, 72, 1476–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrig, A.; Müller, R.H.; Babel, W. Isolation of phenoxy herbicide-degrading Rhodoferax species from contaminated building material. Acta Biotechnol. 1997, 17, 351–356. [Google Scholar] [CrossRef]
- Lee, T.H.; Kurata, S.; Nakatsu, C.H.; Kamagata, Y. Molecular analysis of bacterial community based on 16S rDNA and functional genes in activated sludge enriched with 2,4-dichlorophenoxyacetic acid (2,4-D) under different cultural conditions. Microb. Ecol. 2005, 49, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Streber, W.R.; Timmis, K.N.; Zenk, M.H. Analysis, cloning, and high-level expression of 2,4-dichlorophenoxyacetate monooxygenase gene tfdA of Alcaligenes eutrophus JMP134. J. Bacteriol. 1987, 169, 2950–2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazmiño, D.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Archilla-Ruiz, A.; del Río, L.A.; Sandalio, L.M. Differential response of young and adult leaves to herbicide 2,4-dichlorophenoxyacetic acid in pea plants: Role of reactive oxygen species. Plant. Cell Environ. 2011, 34, 1874–1889. [Google Scholar] [CrossRef] [PubMed]
- Suwa, Y.; Wright, A.D.; Fukumori, F.; Nummy, K.A.; Hausinger, R.P.; Holben, W.E.; Forney, L.J. Characterization of a chromosomally encoded 2,4-dichlorophenoxyacetic acid (2,4-D)/alpha-ketoglutarate dioxygenase from Burkholderia sp. RASC. Appl. Environ. Microbiol. 1996, 62, 2464–2469. [Google Scholar] [PubMed]
- Fulthorpe, R.R.; McGowan, C.; Maltseva, O.V.; Holben, W.E.; Tiedje, J.M. 2,4-Dichlorophenoxyacetic acid-degrading bacteria contain mosaics of catabolic genes. Appl. Environ. Microbiol. 1995, 61, 3274–3281. [Google Scholar] [Green Version]
- Xia, Z.Y.; Zhang, L.; Zhao, Y.; Yan, X.; Li, S.P.; Gu, T.; Jiang, J.D. Biodegradation of the Herbicide 2,4-Dichlorophenoxyacetic Acid by a New Isolated Strain of Achromobacter sp. LZ35. Curr. Microbiol. 2017, 74, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Lappin, H.M.; Greaves, M.P.; Slatert, J.H. Degradation of the Herbicide Mecoprop [2-(2-Methyl-4-Chlorophenoxy) Propionic Acid ] by a Synergistic Microbial Community. Appl. Environ. Microbiol. 1985, 49, 429–433. [Google Scholar] [PubMed]
- Podolska, G. The effectiveness and phytotoxicity of herbicide in buckwheat cv. Kora. Polish, J. Agron. 2014, 19, 17–24. [Google Scholar]
- Polit, J.T.; Praczyk, T.; Pernak, J.; Sobiech, Ł.; Jakubiak, E.; Skrzypczak, G. Inhibition of germination and early growth of rape seed (Brassica napus L.) by MCPA in anionic and ester form. Acta Physiol. Plant. 2014, 36, 699–711. [Google Scholar] [CrossRef]
- Wei, Y.D.; Zheng, H.G.; Hall, J.C. Role of auxinic herbicide-induced ethylene on hypocotyl elongation and root/hypocotyl radial expansion. Pest. Manag. Sci. 2000, 56, 377–387. [Google Scholar] [CrossRef]
- Sarikaya, R.; Yilmaz, M. Investigation of acute toxicity and the effect of 2,4-D (2,4-dichlorophenoxyacetic dichlorophenoxyacetic acid) herbicide on the behavior of the common carp (Cyprinus carpio L., 1758; Pisces, Cyprinidae). Chemosphere 2003, 52, 195–201. [Google Scholar] [PubMed]
- Bukowska, B. Effects of 2,4-D and its metabolite 2,4-dichlorophenol on antioxidant enzymes and level of glutathione in human erythrocytes. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 135, 435–441. [Google Scholar] [CrossRef]
- Martínez-Ruiz, E.B.; Martínez-Jerónimo, F. Exposure to the herbicide 2,4-D produces different toxic effects in two different phytoplankters: A green microalga (Ankistrodesmus falcatus) and a toxigenic cyanobacterium (Microcystis aeruginosa). Sci. Total Environ. 2018, 619–620, 1566–1578. [Google Scholar] [CrossRef] [PubMed]
- Aronzon, C.M.; Sandoval, M.T.; Herkovits, J.; Pérez-Coll, C.S. Stage-dependent toxicity of 2,4-dichlorophenoxyacetic on the embryonic development of a South American toad, Rhinella arenarum. Environ. Toxicol. 2011, 26, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Estevam, E.C.; Nakano, E.; Kawano, T.; de Bragança Pereira, C.A.; Amancio, F.F.; de Albuquerque Melo, A.M.M. Dominant lethal effects of 2,4-D in Biomphalaria glabrata. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2006, 611, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, N.; Hunziker, R.; Escher, B.; Eggen, R. Acute toxicity of (chloro-)catechol-copper combinations in Escherichia coil corresponds to their membrane toxicity in vitro. Environ. Toxicol. Chem. 2001, 20, 239–247. [Google Scholar] [PubMed]
- Lurquin, P.F. Production of a toxic metabolite in 2,4-D-resistant GM crop plants. 3 Biotech. 2016, 6, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Piaia, B.; Alves, C.; Gularte, O.; Teixeira, D.; Cristofari, M.; Ricardo, M.; Carriço, S.; Chimelo, M.; Luiz, R.; Luis, E.; et al. Chemosphere The phytoremediation potential of Plectranthus neochilus on 2,4-dichlorophenoxyacetic acid and the role of antioxidant capacity in herbicide tolerance. Chemosphere 2017, 188, 231–240. [Google Scholar]
- Shi, C.; Sun, Y.; Zheng, Z.; Zhang, X.; Song, K.; Jia, Z.; Chen, Y.; Yang, M.; Liu, X.; Dong, R.; et al. Antimicrobial activity of syringic acid against Cronobacter sakazakii and its effect on cell membrane. Food Chem. 2016, 197, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Ferulic acid impairs rhizogenesis and root growth, and alters associated biochemical changes in mung bean (Vigna radiata) hypocotyls. J. Plant. Interact. 2014, 9, 267–274. [Google Scholar] [CrossRef]


{kind=link}
| Contaminant. | Studied Plant | Plant Secondary Metabolite | Observed Effect | Literature |
|---|---|---|---|---|
| Polychlorinated biphenyls (PCBs) | n.a. | flavonoid, naringin | enhancement of PCBs reduction | [15] |
| Trichloroethene (TCE) | n.a. | cumen | enhancement of the biodegradation of TCE by R. gordoniae up to 75% within 24 h | [18] |
| Dichlorodiphenyldichloroethylene (DDE) | Cucurbita pepo | low molecular weight acids (e.g., citric acid) | enhancement of p,p’-DDE bioavailability in soil | [19] |
| Polycyclic aromatic hydrocarbons (PAH) | Apium graveolens | linoleic acid | enhancement of benzo[α]pyrene removal | [20] |
| 2,4-dichlorophenol (2,4-DCP) | soil samples taken from sites located under Pinus sylvestris, Quercus robur | limonene and α-pinene | induction of 2,4-DCP degradation by indigenous soil microbiota | [21] |
| PH | Concentration (mM) | Sample * | PR |
|---|---|---|---|
| 2,4-D | 0.1 | MSM | 86 |
| MSM + FA | 93 | ||
| MSM + FA + SSE | 83 | ||
| MSM + SE | 98 | ||
| MSM + SE + FA | 100 | ||
| 0.5 | MSM | 100 | |
| MSM + FA | 100 | ||
| MSM + FA + SSE | 99 | ||
| MSM + SE | 100 | ||
| MSM + SE + FA | 97 | ||
| MCPA | 0.1 | MSM | 40 |
| MSM + SA | 11 | ||
| MSM + SA + SSE | 19 | ||
| MSM + SE | 53 | ||
| MSM + SE + SA | 99 | ||
| 0.5 | MSM | 27 | |
| MSM + SA | 5 | ||
| MSM + SA + SSE | 12 | ||
| MSM + SE | 99 | ||
| MSM + SE + SA | 100 |
| PH | Target Gene | 16S rRNA | tfdA alfa | tfdA CI | tfdA CII | tfdA CIII | tfdA | 16S rRNA | tfdA alfa | tfdA CI | tfdA CII | tfdA CIII | tfdA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days of Incubation | 2,4-D/MCPA Concentration (mM) | MSM + SE | MSM + SE + FA | |||||||||||
| 2,4-D | 6 | 0 | + | + | + | + | + | |||||||
| 12 | + | + | + | |||||||||||
| 18 | + | + | + | + | + | |||||||||
| 24 | + | + | + | + | ||||||||||
| 6 | 0.1 | + | + | + | + | |||||||||
| 12 | + | + | + | + | ||||||||||
| 18 | + | + | + | + | + | |||||||||
| 24 | + | + | + | + | ||||||||||
| 6 | 0.5 | + | + | |||||||||||
| 12 | + | + | + | + | ||||||||||
| 18 | + | + | + | |||||||||||
| 24 | + | + | + | |||||||||||
| MSM + SE | MSM + SE + SA | |||||||||||||
| MCPA | 6 | 0 | + | + | + | |||||||||
| 12 | + | + | + | + | ||||||||||
| 18 | + | + | + | + | + | + | ||||||||
| 24 | + | + | + | + | ||||||||||
| 6 | 0.1 | + | + | + | + | + | ||||||||
| 12 | + | + | + | |||||||||||
| 18 | + | + | + | |||||||||||
| 24 | + | + | + | |||||||||||
| 6 | 0.5 | + | + | + | + | + | ||||||||
| 12 | + | + | + | + | + | + | ||||||||
| 18 | + | + | + | + | + | + | + | |||||||
| 24 | + | + | + | + | + | + | + | + | ||||||
| PH | Concentration | Sample Variant * | Nr of Detected Functional Genes | ||||
|---|---|---|---|---|---|---|---|
| Days of Incubation | Total | ||||||
| 6 | 12 | 18 | 24 | ||||
| - | 0 | MSM + SE | 1 | 1 | 2 | 1 | 5 |
| MSM + SE + FA | 2 | 0 | 1 | 1 | 3 | ||
| MSM + SE + SA | 0 | 1 | 2 | 1 | 4 | ||
| 2,4-D | 0.1 | MSM + SE | 1 | 1 | 1 | 1 | 4 |
| MSM + SE + FA | 1 | 1 | 2 | 1 | 5 | ||
| 0.5 | MSM + SE | 0 | 1 | 1 | 1 | 3 | |
| MSM + SE + FA | 0 | 1 | 0 | 0 | 1 | ||
| MCPA | 0.1 | MSM + SE | 0 | 0 | 0 | 0 | 0 |
| MSM + SE + SA | 3 | 1 | 1 | 1 | 6 | ||
| 0.5 | MSM + SE | 3 | 1 | 1 | 3 | 8 | |
| MSM + SE + SA | 0 | 3 | 4 | 3 | 10 | ||
| Class of Microorganisms | Identified Strain | Homology | E-value |
|---|---|---|---|
| β-proteobacteria | Rhodoferax saidenbachensis strain OX0321 | 100% | 0 |
| Achromobacter dolens strain BFHC1 5 | 99% | 1.0 × 10−44 | |
| Burkholderia sp. strain A5 | 99% | 1.0 × 10−44 | |
| Cupriavidus sp. strain CI099 | 99% | 4.0 × 10−44 |
| PH | 2,4-D/MCPA Concentration (mM) | L. sativum | S. alba | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MSM | MSM + PSM * | MSM + SSE | MSM + SE | MSM + SE+ PSM * | MSM | MSM + PSM * | MSM + SSE | MSM + SE | MSM + SE + PSM * | ||
| PE at the Beginning of the Experiment | |||||||||||
| - | - | n.a. | n.a. | n.a. | 51 | n.a. | n.a. | n.a. | n.a. | 57 | n.a. |
| 2,4-D | 0.1 | 100 | n.a. | n.a. | n.a. | n.a. | 98 | n.a. | n.a. | n.a. | n.a. |
| 0.5 | 100 | n.a. | n.a. | n.a. | n.a. | 94 | n.a. | n.a. | n.a. | n.a. | |
| MCPA | 0.1 | 100 | n.a. | n.a. | n.a. | n.a. | 100 | n.a. | n.a. | n.a. | n.a. |
| 0.5 | 100 | n.a. | n.a. | n.a. | n.a. | 100 | n.a. | n.a. | n.a. | n.a. | |
| PE After 24 Days of Incubation | |||||||||||
| 2,4-D | 0.1 | 100 | 98 | 100 | 100 | 100 | 96 | 95 | 97 | 94 | 98 |
| 0.5 | 10 | 100 | 100 | 100 | 100 | 96 | 96 | 96 | 97 | 97 | |
| MCPA | 0.1 | 100 | 100 | 100 | −102 * | −69 * | 100 | 100 | 100 | −34 * | −21 * |
| 0.5 | 100 | 100 | 100 | −47 * | −55 * | 100 | 100 | 100 | −38 * | −35 * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mierzejewska, E.; Baran, A.; Tankiewicz, M.; Urbaniak, M. Removal and Ecotoxicity of 2,4-D and MCPA in Microbial Cultures Enriched with Structurally-Similar Plant Secondary Metabolites. Water 2019, 11, 1451. https://doi.org/10.3390/w11071451
Mierzejewska E, Baran A, Tankiewicz M, Urbaniak M. Removal and Ecotoxicity of 2,4-D and MCPA in Microbial Cultures Enriched with Structurally-Similar Plant Secondary Metabolites. Water. 2019; 11(7):1451. https://doi.org/10.3390/w11071451
Chicago/Turabian StyleMierzejewska, Elżbieta, Agnieszka Baran, Maciej Tankiewicz, and Magdalena Urbaniak. 2019. "Removal and Ecotoxicity of 2,4-D and MCPA in Microbial Cultures Enriched with Structurally-Similar Plant Secondary Metabolites" Water 11, no. 7: 1451. https://doi.org/10.3390/w11071451
APA StyleMierzejewska, E., Baran, A., Tankiewicz, M., & Urbaniak, M. (2019). Removal and Ecotoxicity of 2,4-D and MCPA in Microbial Cultures Enriched with Structurally-Similar Plant Secondary Metabolites. Water, 11(7), 1451. https://doi.org/10.3390/w11071451