1. Introduction
Ponds (standing waterbodies from 25 m
2 to 2 ha in size [
1]) in agricultural landscapes are very common and they cover more than 3% of the Earth’s terrestrial surface area [
2,
3]. Ponds are the most numerous representatives of inland standing waters [
4]. They host unique flora and contribute to regional biodiversity [
5,
6,
7]. The information about the value of ponds is insufficient [
8]. Little is known about their environmental sensitivity, structure, functioning, relations with the surrounding landscapes, and how they are affected by human activity [
9]. Currently, there is a growing interest in pond ecology [
4,
6], mostly due to the recent revelation and acknowledgement of their general ecological importance [
2]. Small farmland ponds are especially vulnerable to climate change and habitat degradation (including pollution) due to their limited volume [
2,
7,
10,
11,
12]. The estimated loss of ponds in the 20th century exceeded 50% in many European countries [
13], and was up to 90% in some regions due to agricultural land drainage and urbanisation [
7,
14]. Recognising the loss of ponds, restoration and reconstruction projects were recently implemented in several regions worldwide [
4]. Ponds may act as important surrogate habitats for species whose natural habitats have been lost [
15]. Ponds may increase the biodiversity of macrophytes and positively affect the occurrence of larger amounts of rare and endemic plant species more so than other freshwater habitats [
1,
2]. However, we generally lack information as to how freshwater habitats, including ponds, support biodiversity [
4]. Most farmland ponds have small catchment areas and specific physicochemical properties [
16]. All these variables cause considerable differences in habitat conditions between big and small ponds [
17,
18,
19]. In view of their potentially high ecological value, information about their biological quality is essential for long-term conservation and management of biodiversity [
20].
Macrophytes create an environment that is fundamentally different from that of the open water and that potentially may have great impact on the interactions between the different trophic levels [
21]. The diversity of macrophytes may increase with the abundance and proximity of other wetland habitats in the landscape [
22], but this is not necessarily always the case [
23], and the positive effect of connectivity may even be reversed by the presence of strong ecological interactions in connected waterbodies [
24]. The regional effects associated with intensive land use (farmlands or urban areas) are very typical for decreasing macrophyte diversity [
25]. A high degree of variability also exists in the relationships between the local environment and the diversity of pond-dwelling macrophytes, but surface area, water chemistry and the hydrological regime are usually ranked among the most influential local factors [
26]. Macrophytes from different groups are likely to respond to the variability of the pond environment in different ways, resulting in different diversity patterns [
25,
27]. Freshwater macrophyte diversity is highly variable because it may involve interactions of several environmental factors [
26]. Aquatic plants may also reduce the content of nutrients and water turbidity [
28,
29]. This group can be used as bioindicators because it reacts to changes in the environment and helps to predict changes in ecosystems [
30,
31]. Rooted submerged macrophytes play an important role in nutrient cycling in ponds by mediating fluxes of nutrients from sediments into the water [
29]. Macrophytes mobilize nutrients either directly (from sediments through root uptake and senescence) or indirectly (from sediments by causing marked fluctuations in pH and oxygen, which enhance the rate of P release from sediments) [
32]. In particular, high pH values (about 9–10) associated with macrophyte photosynthesis can result in ligand exchange with P adsorbed to iron oxide-hydroxides on sediment particles, thus enhancing the rate of P release from sediments [
32].
The hypotheses of our research were: (1) the morphometric variables and physio-chemical properties of water are correlated with the distribution of rooted and floating macrophytes in ponds; (2) there are macrophyte species characteristic of a specific location. The overall aim of the study was to test if there is a correlation between macrophytes’ (rooted and floating) temporal changes and the influence of environmental factors on the ecosystems of ponds in the reference period (2008–2018). The second aim was to assess whether the pond ecosystems (similar in terms of land use) can be characterized by a similar composition of macrophytes.
2. Materials and Methods
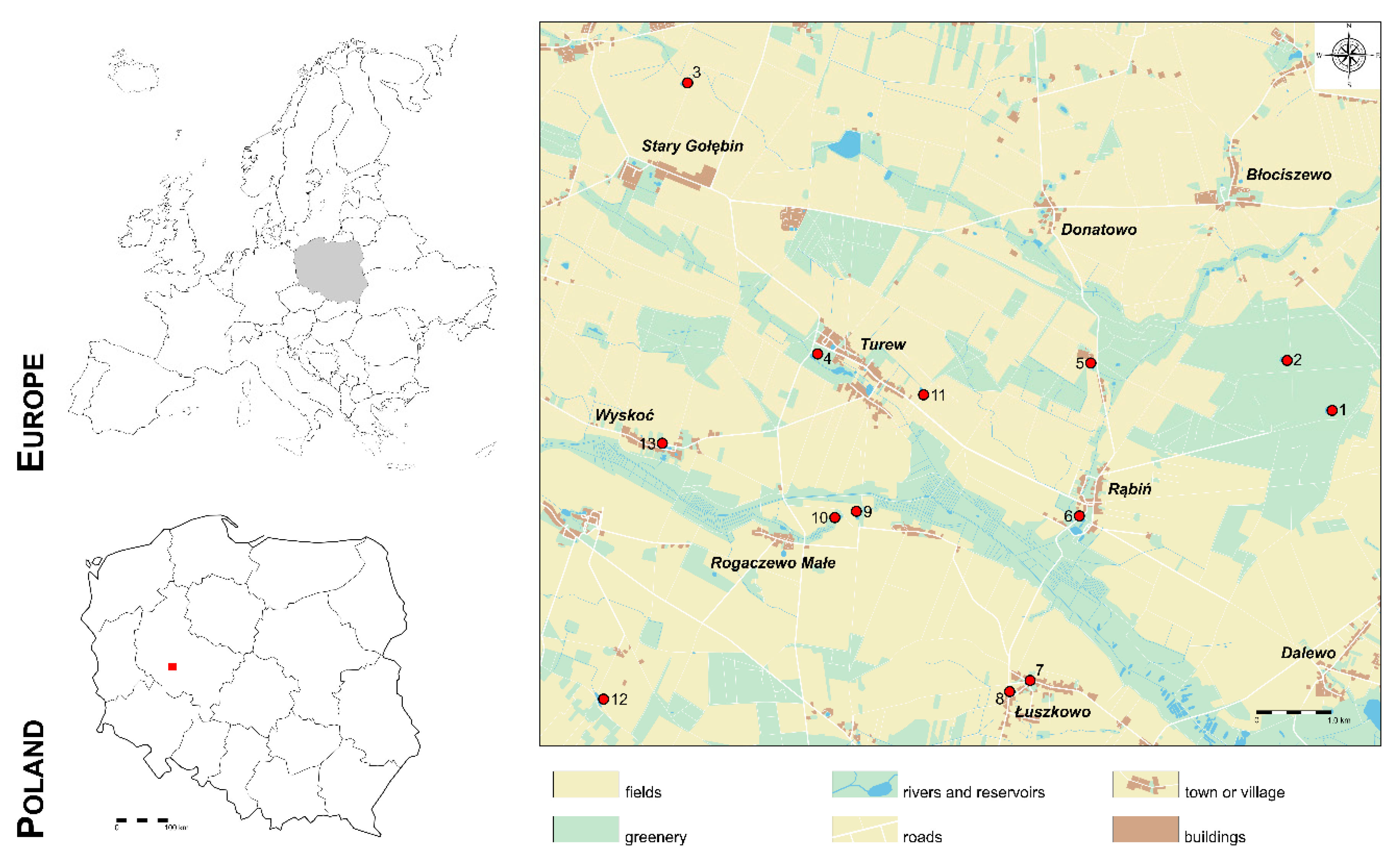
The research was conducted on 13 ponds located in the General Dezydery Chłapowski Scenic Park in Greater Poland Voivodeship, Poland. No specific permissions were required for the locations. It is clearly stated in the XLIV/858/14 Bill of Wielkopolska Voivodeship Parliament (28 April 2014) regarding establishment of the General Dezydery Chłapowski Scenic Park. Our field studies did not involve any endangered or protected species. The ponds differed in their location in the landscape, surface area, shape, and aquatic vegetation. The geographical coordinates and changes in the morphometric parameters of the ponds under analysis are presented in
Table 1. Some ponds were typical field hollows, others were in built-up areas, and two were in a forest complex (
Figure 1).
Forest ponds (Nos. 1 and 2) were in the planted forests: Pond No. 1—deciduous forest, and Pond No. 2—coniferous forest. Pond No. 1 was a natural reservoir, while Pond No. 2 was created in the 1990s. Field ponds (Nos. 3, 5, 9, 10, 11, 12) were all natural reservoirs with the exception of No. 9, which was created by the Polish Academy of Sciences for scientific purposes in 1995. Pond No. 3—surrounded by fields, cultivation of maize, fertilized organically and by minerals. Pond No. 5—surrounded by fields, rye, and wheat cultivation, fertilized organically and by minerals. Pond No. 9—surrounded by meadow, field, and a paved road; rye, wheat, and rape cultivation; fertilized organically and by minerals. Pond No. 10—surrounded by fields; rye cultivation; fertilized organically and by minerals. Pond No. 11—surrounded by fields and a meadow; cultivation of wheat and rape; fertilized organically and by minerals. Pond No. 12—surrounded by fields; cultivation of wheat and rape; fertilized organically and by minerals. Village ponds (Nos. 4, 6, 7, 8, and 13) were ponds created in the villages, without drainage, with the exception of No. 4, which was a natural pond. No wastewater was supplied to any pond and no wastewater was collected. Humans affected none of the ponds directly (e.g., fishing). Pond No. 4—surrounded by an old park where deciduous trees grow. Pond No. 6—surrounded by roads and meadows. Ponds Nos. 7, 8 and 13—located in the center of the village, surrounded by buildings and roads.
∆ (the delta symbol) means the increase or decrease in area and depth in 2018 in relation to the mean area and depth calculated from the ten-year period and is expressed as a percentage:
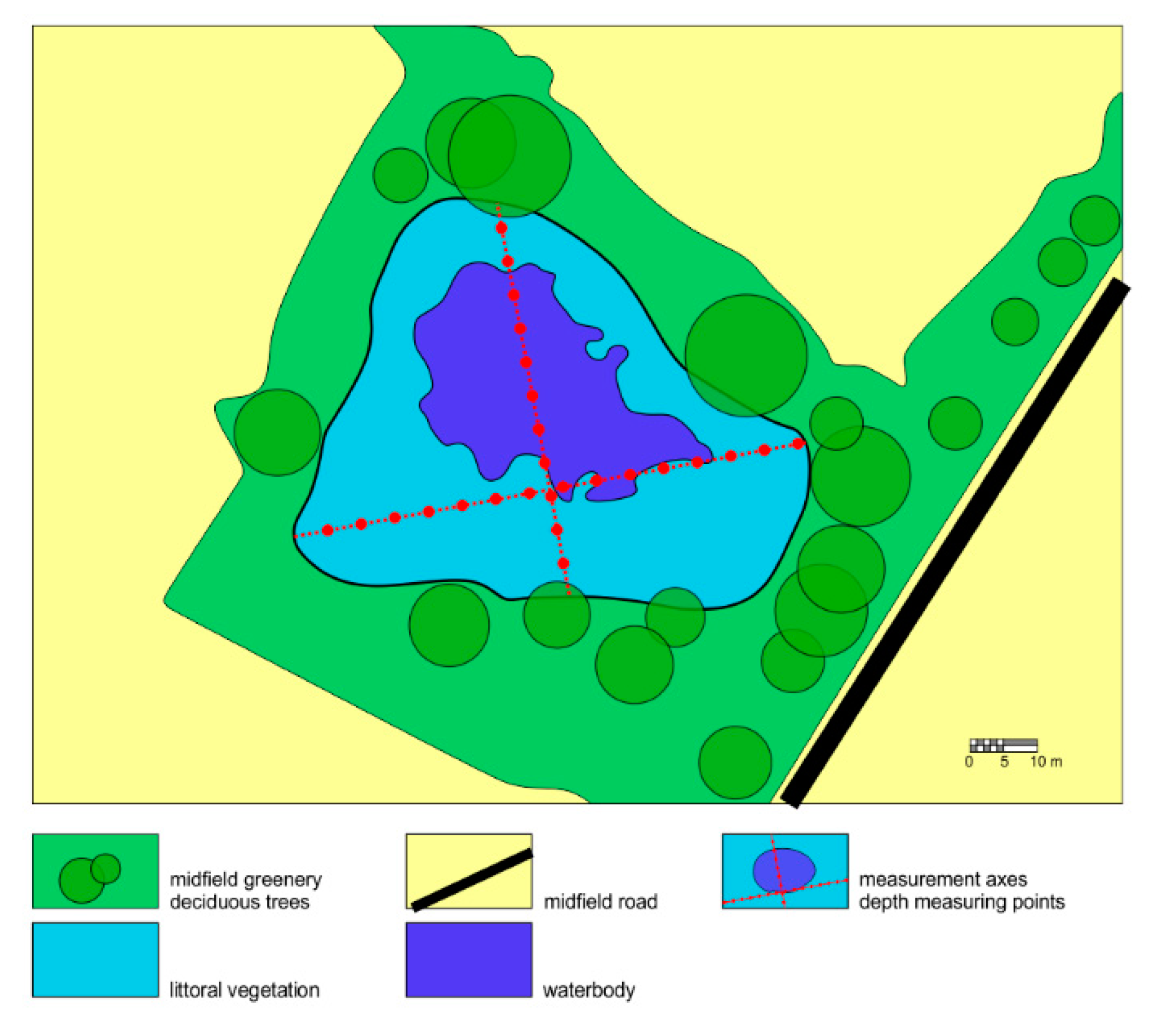
The research spanned a period of ten years (2008–2018), with measurements made every second year. The surface of the studied ponds was measured by a qualified surveyor using the Pentax R-1500N (TI Asahi Co., Ltd., Saitama, Japan) total station theodolite with TopoLite software (TI Asahi Co., Ltd., Saitama, Japan), and mean depth was measured by a depth pole (
Figure 2).
The morphometric parameters, such as the depth and area of ponds, were measured at the beginning of July in 2008, 2010, 2012, 2014, 2016, and 2018—six times in total during the research period. Macrophytes were identified at the beginning of July and then verified twice (on 15 July and 31 July in terms of area they occupied) in the research year, which gives three observations per year. This was the optimal period for the identification of macrophytes during the growing season. Similarly, physicochemical parameters of water in ponds were collected and tested (three observations per year).
Samples of water were analyzed for the content of NH
4+, NO
3−, NO
2−, TP, PO
43−, Na, K, Mg, Ca, and Fe—according to standard methods [
33]. The pH value and temperature were also measured using a waterproof CP-411 device with a glass electrode EPS-1 type. We collected water samples at depths of 20–50 cm, about 1 m away from the shore of ponds.
The following methods were used to analyze the content of elements in AAS AAnalyst 200 spectrometer with specific optimisation parameters (
Table A1):
Flame Atomic Absorption Spectrometry (F-AAS)—Ca, Mg, and Fe;
Atomic Emission Spectrometry (AES)—Na, K.
In order to determine the differences between ponds in terms of anthropogenic pressure, the ECELS methodology proposed by Sala et al. [
34] was used. The index is composed of five components (morphology, human activity, water aspects, emergent vegetation, hydrophytic vegetation), each assessing an independent aspect of the conservation status of a wetland. Each component has a modifying section that assesses additional particularities of the component where one or several options can be chosen resulting in addition or subtraction of points. The score obtained for each component cannot exceed a maximum value nor have negatives values. The maximum values of the five components are 20, 20, 10, 30 and 20, respectively. The sum of all the values obtained for each component gives the ECELS index final score, which can range between 0 and 100. A categorization of the values is proposed following the guidelines of the Water Framework Directive (2000/60/EC).
The species composition of macrophytes in the 13 ponds was investigated every second year during the ten-year research period (2008–2018). The species of macrophytes in ponds were listed and the percentage of the area occupied by dominant species (a share of over 1% in pond) was determined [
35].
In order to determine the indicator species for ponds in different land use types, the IndVal method described by Dufrêne and Legendre [
36] was used. For each species
i in each site group
j, we computed the product of
Aij (which is the mean abundance of species
i in the sites of group
j compared to all groups in the study) by
Bij, which is the relative frequency of occurrence of species
i in the sites of group
j, as follows:
Nindividualsij—the mean number of individuals of species i across sites of group j.
Nindividualsi—the sum of the mean numbers of individuals of species i over all groups.
Nsitesij—the number of sites in cluster j where species i is present.
Nsitesj—the total number of sites in that cluster. Bij reaches a maximum when species i is present in all objects of cluster j.
Latin names of the macrophytes were used according to the Plant List [
37].
Statistical analyses and models were based on discriminant analysis. Canonical variate analysis (CVA), which is the canonical variant of Fisher’s linear discriminant analysis (LDA), was used to construct a CCA analysis [
38]. Progressive stepwise analysis was applied to find which variables determined the distribution of macrophytes in ponds to the greatest extent. CCA analysis was preceded by stepwise regression analysis, which eliminated statistically insignificant variables (
p > 0.05 and F < 2). A Monte Carlo permutation test was applied to determine the significance limit (the number of permutations: 9999). We have also applied linear regression, multiple regression and correlations in presented paper. All multiple regression and ANOVA analyses were performed with the PQstat software.
3. Results
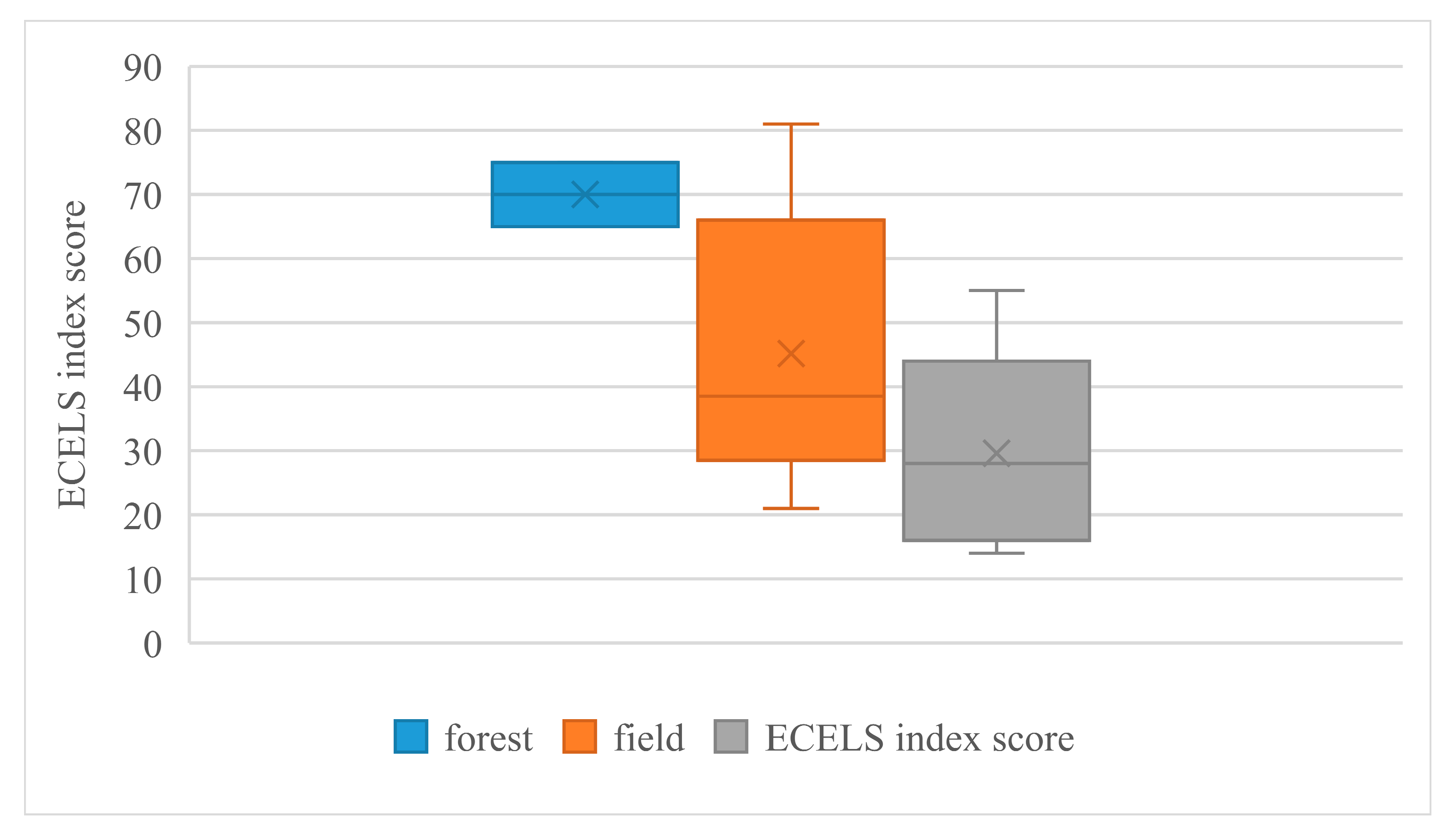
Figure 3 presents the ranges of ECELS values for ponds located in areas near forests, fields and villages. According to the analysis, the highest values (good and moderate) were characteristic of ponds located in the forest. Village ponds, on the other hand, corresponded to poor and bad levels of the ECELS indicator. The low scores were associated with components related to human activity around the ponds and the lack of submerged and floating macrophytes (
Table 2).
The ponds analyzed in the study were found in land transformed by agriculture. They were in forests, fields, and in villages. These ponds were small elements of the environment. By analyzing using repeated measures ANOVA, we can conclude that most ponds differed in terms of change of their area over time. This may indicate that pond area was not influenced by one factor only, but by a group of environmental variables (
Table 3).
All ponds were very shallow—their average depth ranged from 15 cm to 1.5 m. A similar relationship was observed in changes of depth over time in the analyzed ponds. The analysis showed that the changes in depth of tested ponds over time differed significantly in most ponds. Therefore, we cannot say that one factor could have a clear influence on changes of this parameter over time (
Table 4).
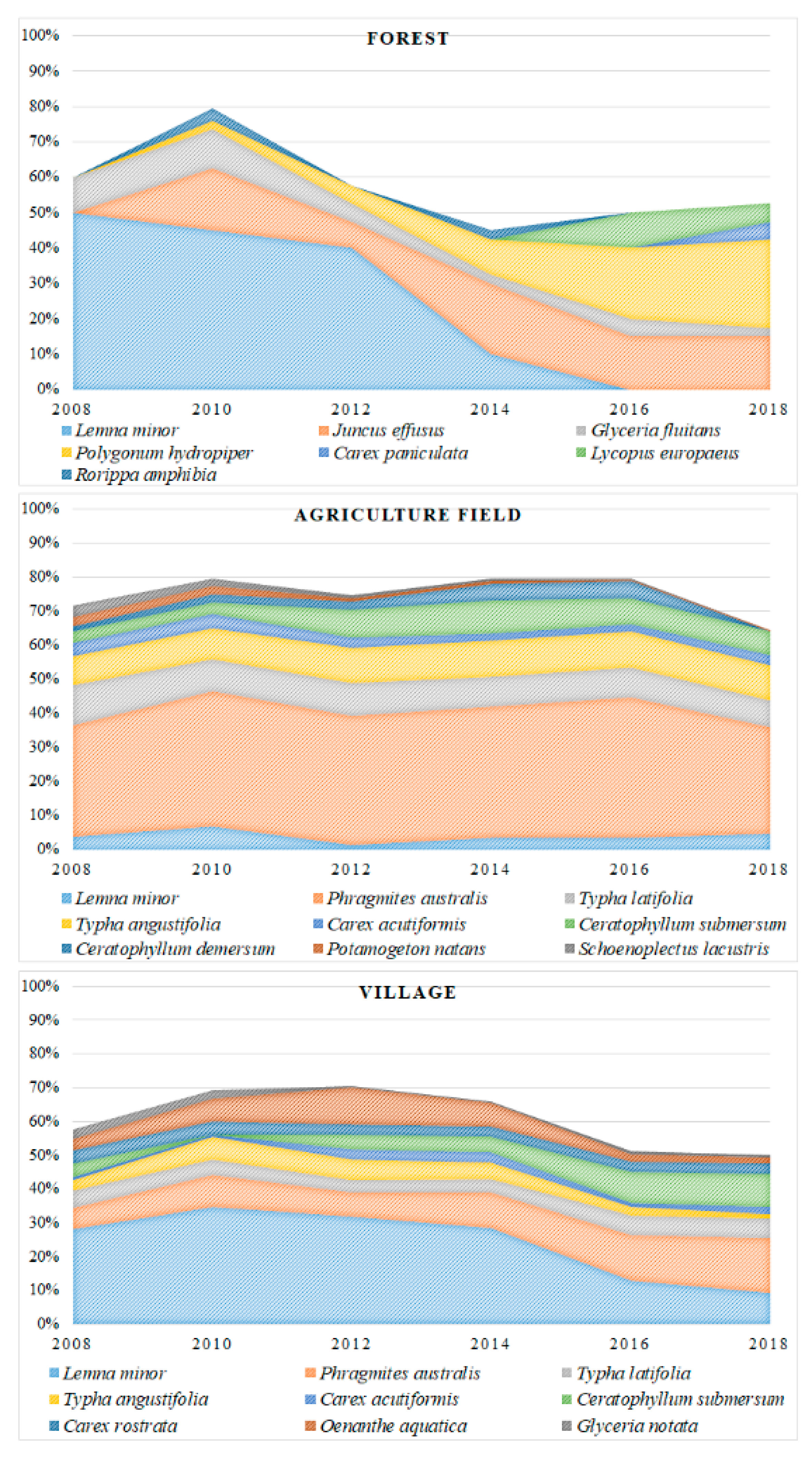
Macrophytes occupied various surface areas in the researched ponds (
Figure 4). Forest ponds were occupied by macrophytes to the smallest extent and had the smallest share of flora species associated with the aquatic environment.
Lemna minor L. was a dominating species at the beginning of the research period. However, in the last two years of the study, the largest increase of surface area was found for
Juncus effusus L. and
Polygonum hydropiper L. A significantly higher percentage of macrophytes’ surface area coverage was found in the case of agriculture ponds, where
Phragmites australis (Cav.) Trin. ex Steud predominated. Village ponds were characterized by a decrease of
Lemna minor L. surface area coverage and an increase in the area occupied mainly by two species:
Phragmites australis (Cav.) Trin. ex Steud and
Ceratophyllum submersum L.
In order to point out indicator species for different land use areas (forests, fields, villages), the IndVal method was applied [
36]. Then, a Monte Carlo test was carried out to determine the probability of a given IndVal value.
Table 5 includes those species for which
p < 0.05. Data analysis enabled the identification of three indicator species for ponds located in forest areas, six indicator species for ponds located on agricultural areas (fields), and five for ponds located in villages.
Table 6 presents linear regressions prepared for species during the research period (
p < 0.05). By far the most expansive species (the ability to exploit all available resources might correspond to the expansion of individual species in natural habitats and represent a criterion for their potential expansiveness) in the studied ponds was
Phragmites australis (Cav.) Trin. ex Steud, which increased coverage area significantly in three ponds (out of eight) located in villages and fields. On the other hand, during the research period, the area occupied by
Lemna minor L. was reduced in three ponds (out of seven), although it is one of the most common and characteristic species for ponds, indicating their low ecological status and eutrophication [
39]. In other cases, single increases or decreases were observed for the area occupied by the species listed in
Table 6. Presented trend lines are linear functions of the surface change of the species examined over time.
x—time measured in years.
y—area occupied by a given species in the total area of pond (expressed in %).
The conducted stepwise regression analysis answered the question as to whether there is a pattern for environmental variables and distribution of macrophytes in time and space. The analysis allowed the preparation of regression models for 15 species tested—53.57% of all analyzed species (
Table 7).
The variables that most often and most strongly correlated with the species studied were air temperature, Na, NH
4+, and NO
3− concentration levels (significant for seven species); K concentration levels were significant for six species; water temperature and Fe concentration levels were significant for for five species (
Table 8 and
Table A2).
In order to check whether the analyzed variables are related to each other, we performed a Pearson’s correlation analysis. On one hand it showed a strong negative correlation between pH and Fe and NO
2−, and on the other, a positive correlation with K. Very strong positive correlations were observed between K and Na, PO
43− and TP. We also noticed very strong correlations between Mg and Ca, as well as between Fe and NH
4+ (
Table 9).
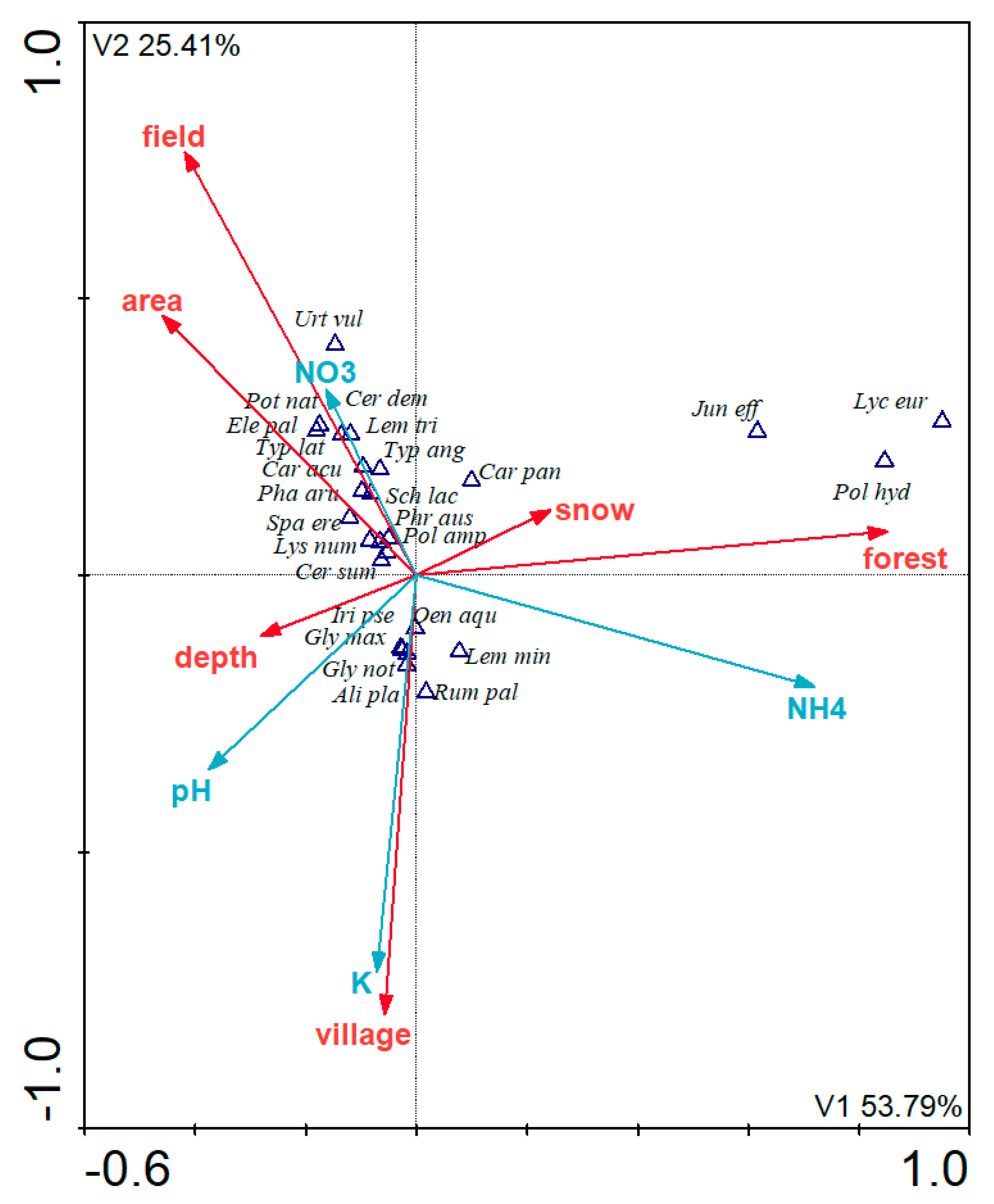
The occurrence of specific macrophyte species were mostly associated with the following variables: the type of pond, its area, and the concentrations of NH
4+, NO
3−, Na, and Fe (
Table 10). Additionally, three plant species were excluded from CCA analysis as outliers. These were:
Carex rostrata Stokes (very strongly correlated with NO
3−), as well as
Rorippa amphibia (L.)
Besser and
Glyceria fluitans (L.)
R. Br. (very strongly correlated with NH
4+ and Fe). Three homogenous groups with similar environmental preferences were identified (
Figure 5).
4. Discussion
So far studies on vegetation in ponds have mostly focused on their floristic composition and the phytocoenosis structure. However, little is known about the relation between these plant communities and their habitats (physical and chemical properties of water and soil). Studies on lakes proved the noticeable dependency between plant communities and specific habitat parameters [
40,
41,
42]. Finding the correlation between the macrophyte diversity and the method of land use in the areas surrounding ponds is necessary not only to understand the mechanisms of maintaining macrophyte diversity but also to provide important information which will help to maintain biodiversity [
25].
In comparison with lakes and other larger freshwater bodies, there was a higher share of reed and pleustophyte communities (Phragmitetea and Lemnetea classes) and a smaller share of communities belonging to the Potametea group among the vegetation in ponds. However, there was a typical feature of aquatic vegetation—one dominant species in phytocoenoses [
43,
44]. Our research findings confirmed this observation.
Existing studies also observed considerable fluctuations in the water level in ponds located in fields and emphasized that this feature distinguished ponds from larger and more stable ecosystems [
27,
41]. Studies conducted all over the world revealed a positive correlation between the small size and low depth of ponds. On one hand, this may result in better light conditions for macrophytes and stimulate their growth [
45]. On the other hand, ponds located in fields are more likely to dry out during the summer [
46,
47]. The variation in water level had a certain influence on the growth of macrophytes [
48,
49]. Those findings were consistent with the results obtained by Wang et al. [
50]. Our research did not confirm the above data for all species we studied. Our research and analysis allowed us to conclude a statistically significant positive correlation between the depth of ponds and the growth of
Phragmites australis (Cav.) Trin. ex Steud and
Typha angustifolia L., and a statistically significant negative correlation between depth and growth of
Glyceria fluitans (L.) R. Br.,
Glyceria maxima (Hartm.) Holmb.
, and
Alisma plantago-aquatica L. Research conducted in the Mediterranean and semi-arid areas proved that droughts may cause lower water levels and even temporal total disappearance of ponds located in fields [
51]. Our research and analyses did not let us conclude that ponds are in danger of shallowing.
Existing studies found that the growth of macrophytes had a significant correlation with air temperature [
52,
53]. For example, the rise in annual average air temperature can promote continuous growth of macrophytes. This finding was consistent with the conclusions obtained by Wang et al. [
50]. Our studies in the ten-year period showed that a statistically significant positive correlation between air temperature and macrophyte growth concerns only a few species:
Juncus effusus L.,
Lycopus europaeus L.,
Polygonum hydropiper L.,
Glyceria fluitans (L.) R. Br.,
and Rumex palustris Sm. We have also found a statistically significant negative correlation between air temperature and macrophyte growth in the case of
Lemna minor L. and
Typha latifolia L. Our studies also showed a statistically significant positive correlation between water temperature and macrophyte development of
Typha latifolia L., and a statistically significant negative correlation between water temperature and macrophyte development for
Lycopus europaeus L.,
Polygonum hydropiper L.,
Glyceria fluitans (L.) R. Br., and
Rumex palustris Sm.
The physicochemical status of water, which largely controls the biodiversity of macrophytes, is affected by the land use method around a pond [
8,
54,
55,
56]. Research results show that the quality of water in ponds located in the catchment area covered by forests was better than the quality of water in the catchment area where land was used differently [
57,
58,
59]. The analysis of the water samples collected during our ten-year research period showed that the water from the forest ponds had the lowest pH as well as the lowest Ca, Na, and Mg concentrations. On one hand, the samples of water from these ponds contained more Fe and NH
4+ than the water from the other ponds. An increase of NH
4+ may be caused either by greater decomposition or anoxia (which can be very harmful for many species) [
60] or by lack of macrophyte species in ponds [
61]. On the other hand, forests reduce surface runoff of nutrients and increase the filtration and interception of these compounds in catchment areas [
62]. Existing studies showed that the distribution area variation had a significant positive correlation with NH
4+ concentration [
63], which was different from Wang et al.’s [
50] study. The results of our studies indicate that a statistically significant positive correlation between the occurrence of macrophytes and the content of NH
4+ in water occurs only in the case of ponds located in forest areas. In the case of ponds in the fields, we found a positive correlation between the occurrence of macrophytes and the content of NO
3−. In the case of villages, we found a positive correlation between the occurrence of macrophytes and the content of K, Na, and total P. N and P are key elements for the existence and health of macrophyte communities, but an excessive load of nutrients may result in eutrophication [
64]. Research conducted on ponds in Patagonia showed that the concentrations of nutrients were good indicators of the state and quality of water. The content of nutrients in ponds affected by sewage should be higher than in more isolated ponds [
65].
Researchers studied the correlation between the species abundance of flora and fauna and the land use in the areas surrounding ponds [
66,
67]. Our analyses confirmed the correlation between the flora species and land use type.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




